
Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana



Abstract
1. Introduction
2. Results
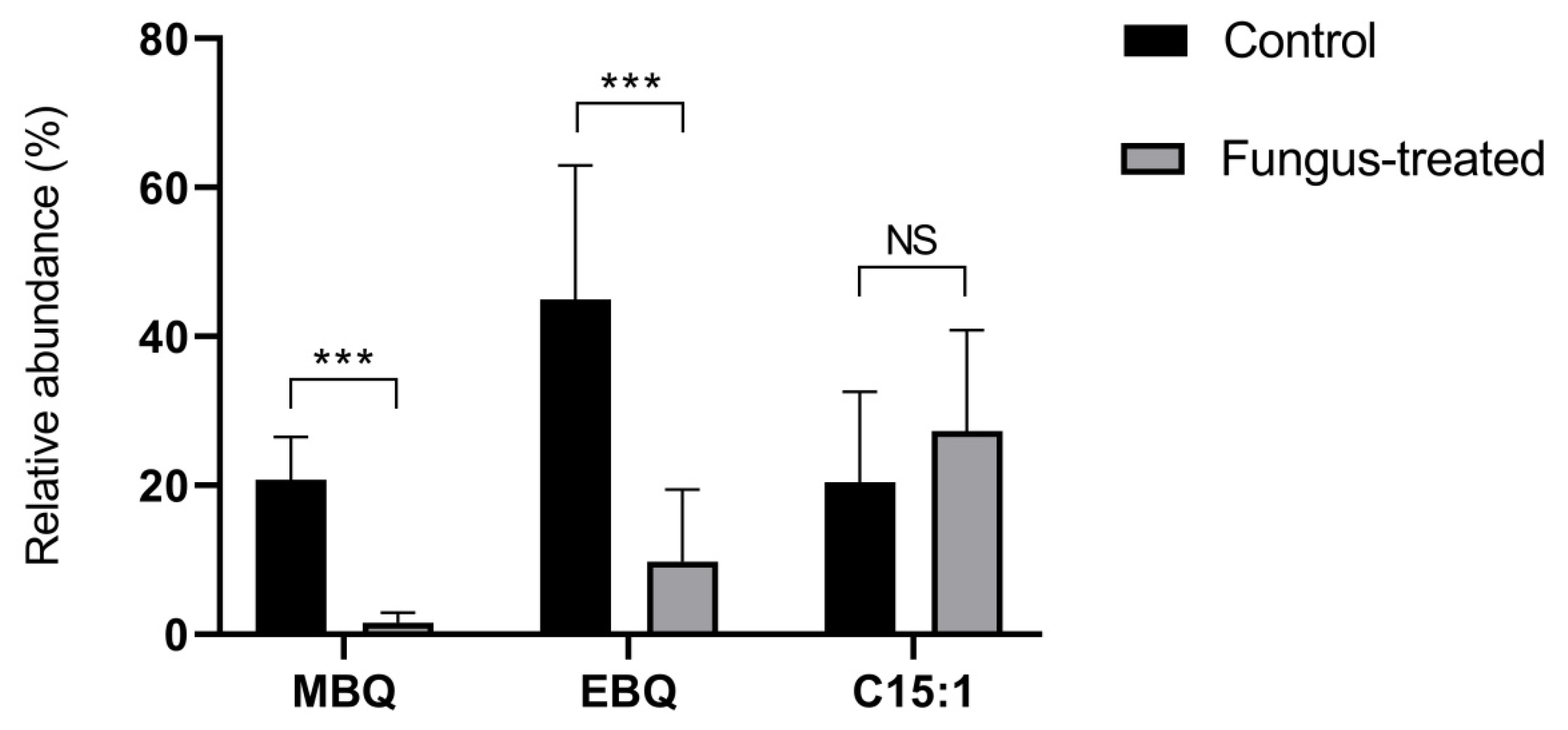
2.1. Volatile Organic Compounds (VOCs) Analysis by Solid Phase Micro Extraction (SPME) Coupled to Gas Chromatography–Flame Ionization Detection (GC-FID)
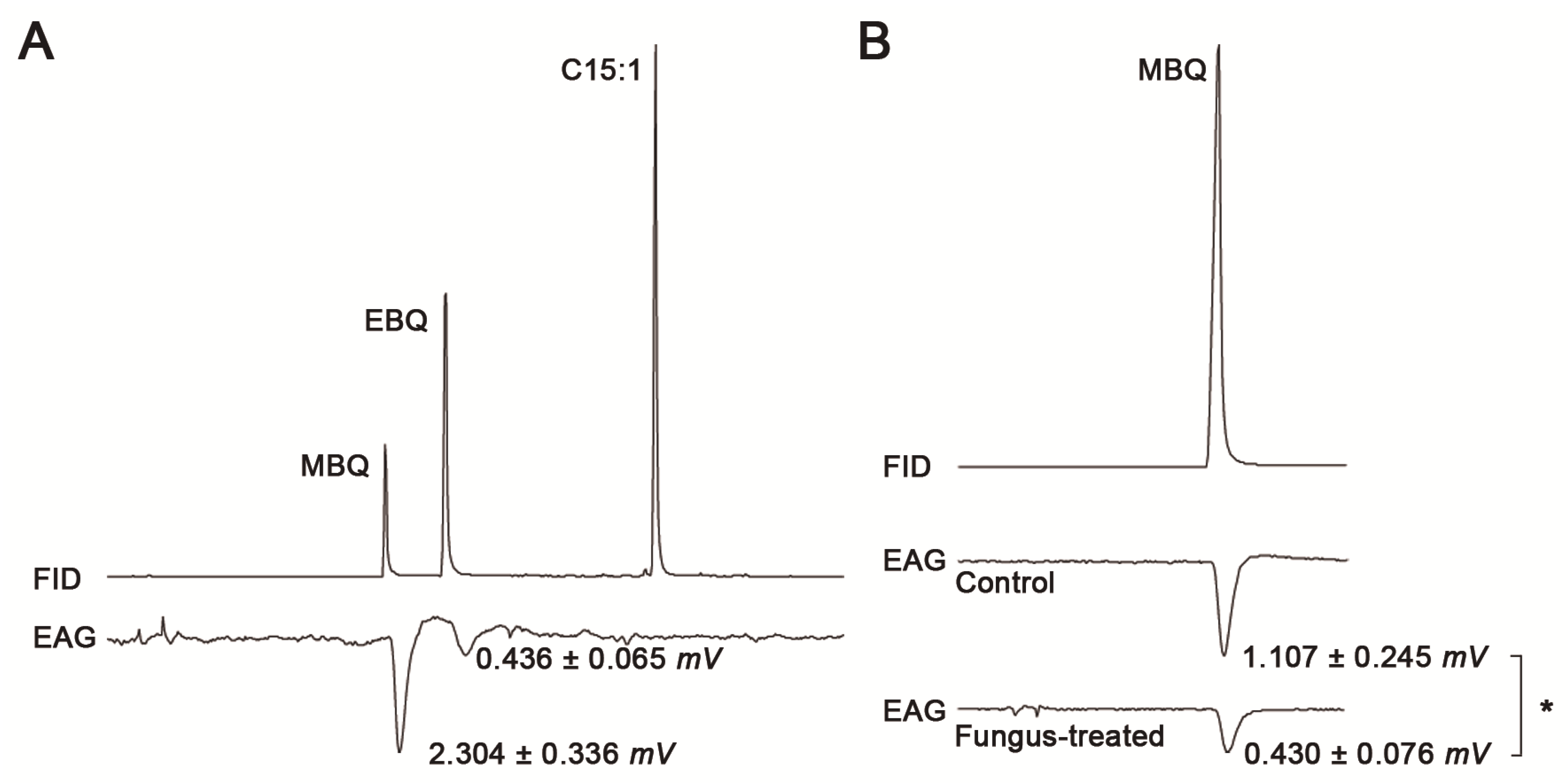
2.2. VOC Perception by Gas Chromatography–Electroantennographic Detection (GC-EAD)
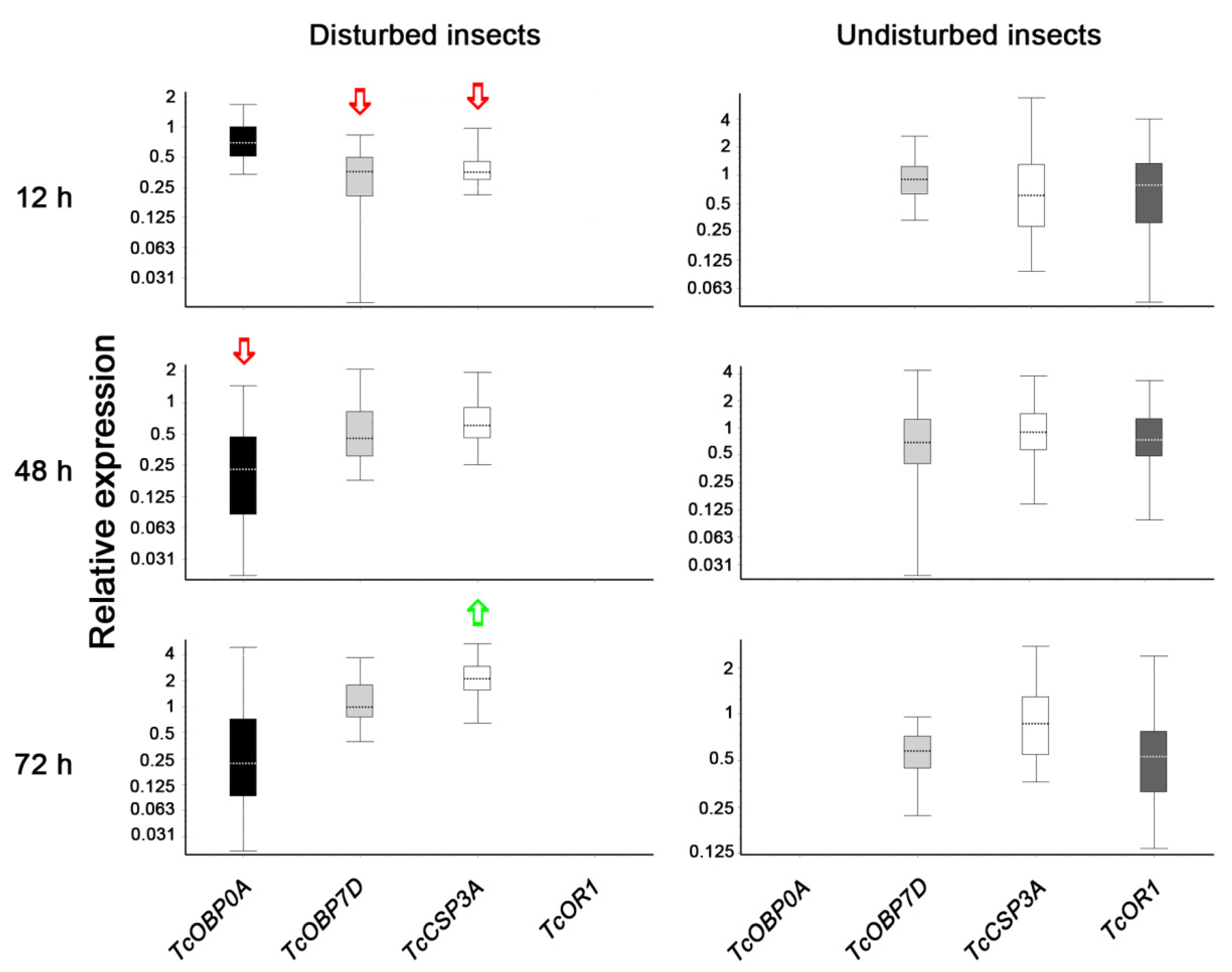
2.3. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. Fungi
4.3. Tribolium Castaneum Treatment with Beauveria bassiana Conidia
4.4. Volatile Organic Compound (VOC) Analysis by Solid Phase Micro Extraction (SPME) Coupled to Gas Chromatography–Flame Ionization Detection (GC-FID)
4.5. Volatile Organic Compound (VOC) Perception by Gas Chromatography–Electroantennographic Detection (GC-EAD)
4.6. Identification and Expression Analysis of Potential Perception Genes by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conchou, L.; Lucas, P.; Meslin, C.; Proffit, M.; Staudt, M.; Renou, M. Insect odorscapes: From plant volatiles to natural olfactory scenes. Front. Physiol. 2019, 10, 972. [Google Scholar] [CrossRef]
- Brown, W.L., Jr.; Eisner, T.; Whittaker, R.H. Allomones and kairomones: Transspecific chemical messengers. Bioscience 1970, 20, 21. [Google Scholar] [CrossRef]
- Roth, L.M. Studies on the gaseous secretion of Tribolium confusum Duval II. The odoriferous glands of Tribolium confusum. Ann. Entomol. Soc. Am. 1943, 36, 397–424. [Google Scholar] [CrossRef][Green Version]
- Eisner, T. Spray mechanism of the cockroach Diploptera punctata. Science 1958, 128, 148–149. [Google Scholar] [CrossRef]
- Tschinkel, W.R. 1975. A comparative study of the chemical defensive system of tenebrionid beetles: Chemistry of the secretions. J. Insect Physiol. 1975, 21, 753–783. [Google Scholar] [CrossRef]
- Villaverde, M.L.; Juárez, M.P.; Mijailovsky, S. Detection of Tribolium castaneum (Herbst) volatile defensive secretions by solid phase microextraction–capillary gas chromatography (SPME–CGC). J. Stored Prod. Res. 2007, 43, 540–545. [Google Scholar] [CrossRef]
- Duehl, A.J.; Arbogast, R.T.; Teal, P.E. Density-related volatile emissions and responses in the red flour beetle, Tribolium castaneum. J. Chem. Ecol. 2011, 37, 525–532. [Google Scholar] [CrossRef]
- Balakrishnan, K.; Holighaus, G.; Weißbecker, B.; Schütz, S. Electroantennographic responses of red flour beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) to volatile organic compounds. J. Appl. Entomol. 2017, 141, 477–486. [Google Scholar] [CrossRef]
- Faustini, D.L.; Burkholder, W.E. Quinone-aggregation pheromone interaction in the red flour beetle. Anim. Behav. 1987, 35, 601–603. [Google Scholar] [CrossRef]
- Pelosi, P.; Calvello, M.; Ban, L. Diversity of odorant-binding proteins and chemosensory proteins in insects. Chem. Senses 2005, 30, i291–i292. [Google Scholar] [CrossRef]
- Dippel, S.; Oberhofer, G.; Kahnt, J.; Gerischer, L.; Opitz, L.; Schachtner, J.; Angeli, S. Tissue-specific transcriptomics, chromosomal localization, and phylogeny of chemosensory and odorant binding proteins from the red flour beetle Tribolium castaneum reveal subgroup specificities for olfaction or more general functions. BMC Genom. 2014, 15, 1141. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef]
- Araújo, J.P.; Hughes, D.P. Diversity of entomopathogenic fungi: Which groups conquered the insect body? Adv. Genet. 2016, 94, 1–39. [Google Scholar] [CrossRef]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies (Review). Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz Urquiza, A.; Huarte Bonnet, C.; Fan, Y.; Juárez, M.P.; Keyhani, N.O. Tenebrionid secretions and a fungal benzoquinone oxidoreductase form competing components of an arms race between a host and pathogen. Proc. Natl. Acad. Sci. USA 2015, 112, 3651–3660. [Google Scholar] [CrossRef]
- Padín, S.B.; Dal Bello, G.M.; Vasicek, A.L. Pathogenicity of Beauveria bassiana for adults of Tribolium castaneum (Col.: Tenebrionidae) in stored grains. Entomophaga 1997, 42, 569–574. [Google Scholar] [CrossRef]
- Rice, W.C.; Cogburn, R.R. Activity of the entomopathogenic fungus Beauveria bassiana (Deuteromycota: Hyphomycetes) against three coleopteran pests of stored grain. J. Econ. Entomol. 1999, 92, 691–694. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Howard, R.W. Diatomaceous earth increases the efficacy of Beauveria bassiana against Tribolium castaneum larvae and increases conidia attachment. J. Econ. Entomol. 2004, 97, 273–280. [Google Scholar] [CrossRef]
- Lord, J.C. Eficcacy of Beauveria bassiana for control of Tribolium castaneum with reduced oxygen and increased carbon dioxide. J. Appl. Entomol. 2009, 133, 101–107. [Google Scholar] [CrossRef]
- Pedrini, N.; Villaverde, M.L.; Fuse, C.B.; Dal Bello, G.; Juárez, M.P. Beauveria bassiana infection alters colony development and defensive secretions of the beetles Tribolium castaneum and Ulomoides dermestoides (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2010, 103, 1094–1099. [Google Scholar] [CrossRef]
- Roth, L.M.; Eisner, T. Chemical defenses of arthropods. Ann. Rev. Entomol. 1962, 7, 107–136. [Google Scholar] [CrossRef]
- Eisner, T.; Meinwald, J. Defensive secretions of arthropods: Insects, millipedes, and some of their relatives, discharge noxious secretions that repel predators. Science 1966, 153, 1341–1350. [Google Scholar] [CrossRef]
- Dettner, K.; Schwinger, G. Defensive secretions of three Oxytelinae rove beetles (Coleoptera: Staphylinidae). J. Chem. Ecol. 1982, 8, 1411–1420. [Google Scholar] [CrossRef]
- Pointer, M.D.; Gage, M.J.G.; Spurgin, L.G. Tribolium beetles as a model system in evolution and ecology. Heredity 2021, 126, 869–883. [Google Scholar] [CrossRef]
- Yezerski, A.; Gilmor, T.P.; Stevens, L. Genetic analysis of benzoquinone production in Tribolium confusum. J. Chem. Ecol. 2004, 30, 1035–1044. [Google Scholar] [CrossRef]
- Verheggen, F.; Ryne, C.; Olsson, P.O.; Arnaud, L.; Lognay, G.; Högberg, H.E.; Löfstedt, C. Electrophysiological and behavioral activity of secondary metabolites in the confused flour beetle, Tribolium confusum. J. Chem. Ecol. 2007, 33, 525–539. [Google Scholar] [CrossRef]
- Gokhale, C.S.; Traulsen, A.; Joop, G. Social dilemma in the external immune system of the red flour beetle? It is a matter of time. Ecol. Evol. 2017, 7, 6758–6765. [Google Scholar] [CrossRef]
- Rath, A.; Benita, M.; Doron, J.; Scharf, I.; Gottlieb, D. Social communication activates the circadian gene Tctimeless in Tribolium castaneum. Sci. Rep. 2021, 11, 16152. [Google Scholar] [CrossRef]
- Ockenfels-Syntech. Electroantennography a Practical Introduction. 2004; 29p. Available online: http://www.ockenfels-syntech.com/wp-content/uploads/EAGpract.pdf (accessed on 19 April 2022).
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MBQ (ng/µL) | EAD Response (mV) |
|---|---|
| 300 | 0.22 ± 0.11 a |
| 514 | 1.11 ± 0.25 b |
| 1800 | 0.77 ± 0.32 b |
| Gene (Acronym Used) | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| TcOBP7D | TGCTCCTCTTTCTCGCTTTGGC | TTTGGCGTCGTCGGTGAAGTC |
| TcOBP0A | CGTGAAGGCTTCTGCATGCTTG | CGCCGTCTCCCAATTCACTTTC |
| TcCSP3A | CGGGACGTCATTCCAGATGCTC | TGTTGCCAATCGCTGTTGCG |
| TcOR1 | GGCGATCAAATACTGGGTGGAG | AACAGCAAATAGCCCAGAACCG |
| RP49 1 | TGACCGTTATGGCAAACTCA | TAGCATGTGCTTCGTTTTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davyt-Colo, B.; Girotti, J.R.; González, A.; Pedrini, N. Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana. Pathogens 2022, 11, 487. https://doi.org/10.3390/pathogens11050487
Davyt-Colo B, Girotti JR, González A, Pedrini N. Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana. Pathogens. 2022; 11(5):487. https://doi.org/10.3390/pathogens11050487
Chicago/Turabian StyleDavyt-Colo, Belén, Juan R. Girotti, Andrés González, and Nicolás Pedrini. 2022. "Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana" Pathogens 11, no. 5: 487. https://doi.org/10.3390/pathogens11050487
APA StyleDavyt-Colo, B., Girotti, J. R., González, A., & Pedrini, N. (2022). Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana. Pathogens, 11(5), 487. https://doi.org/10.3390/pathogens11050487