
The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors

Abstract
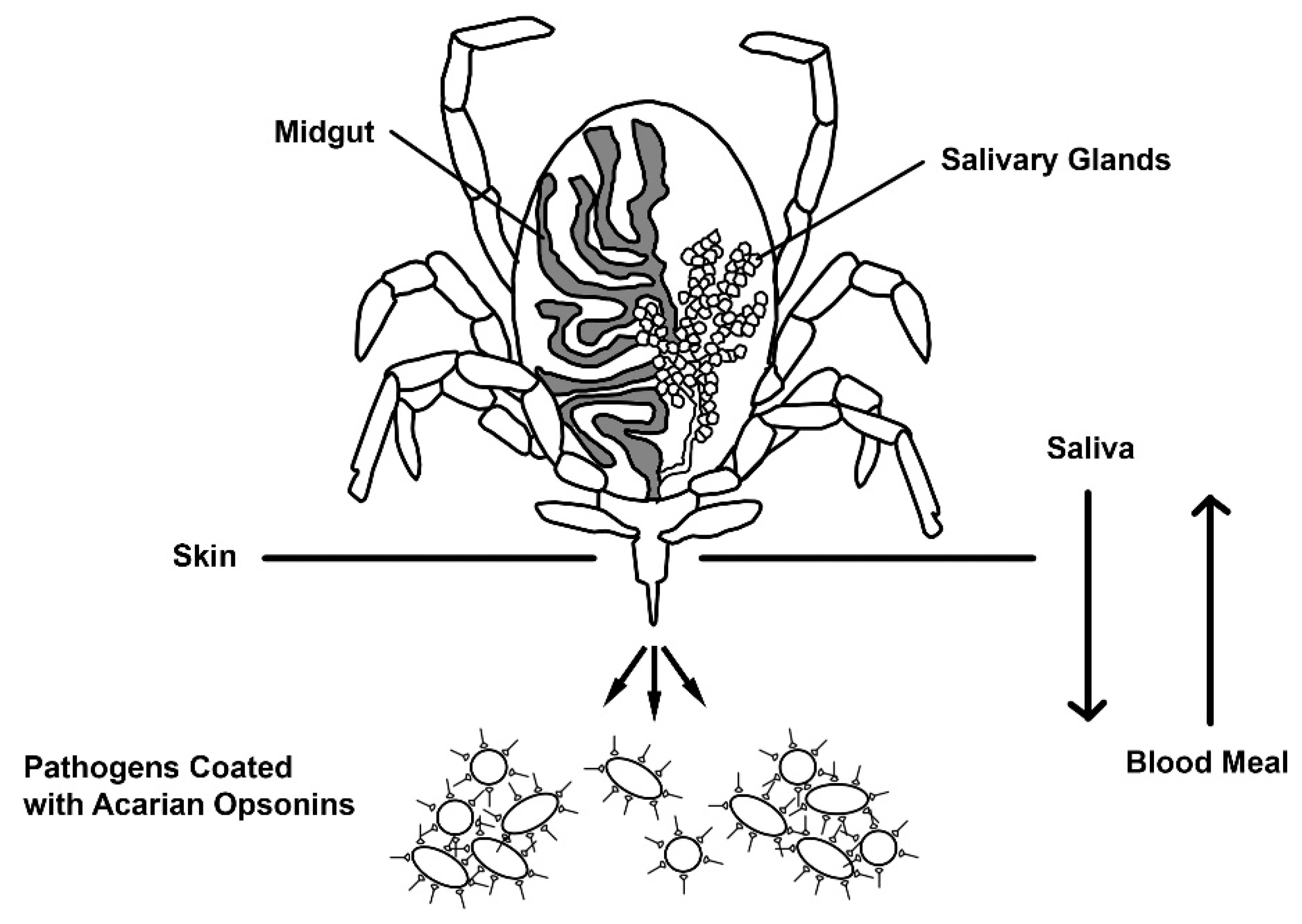
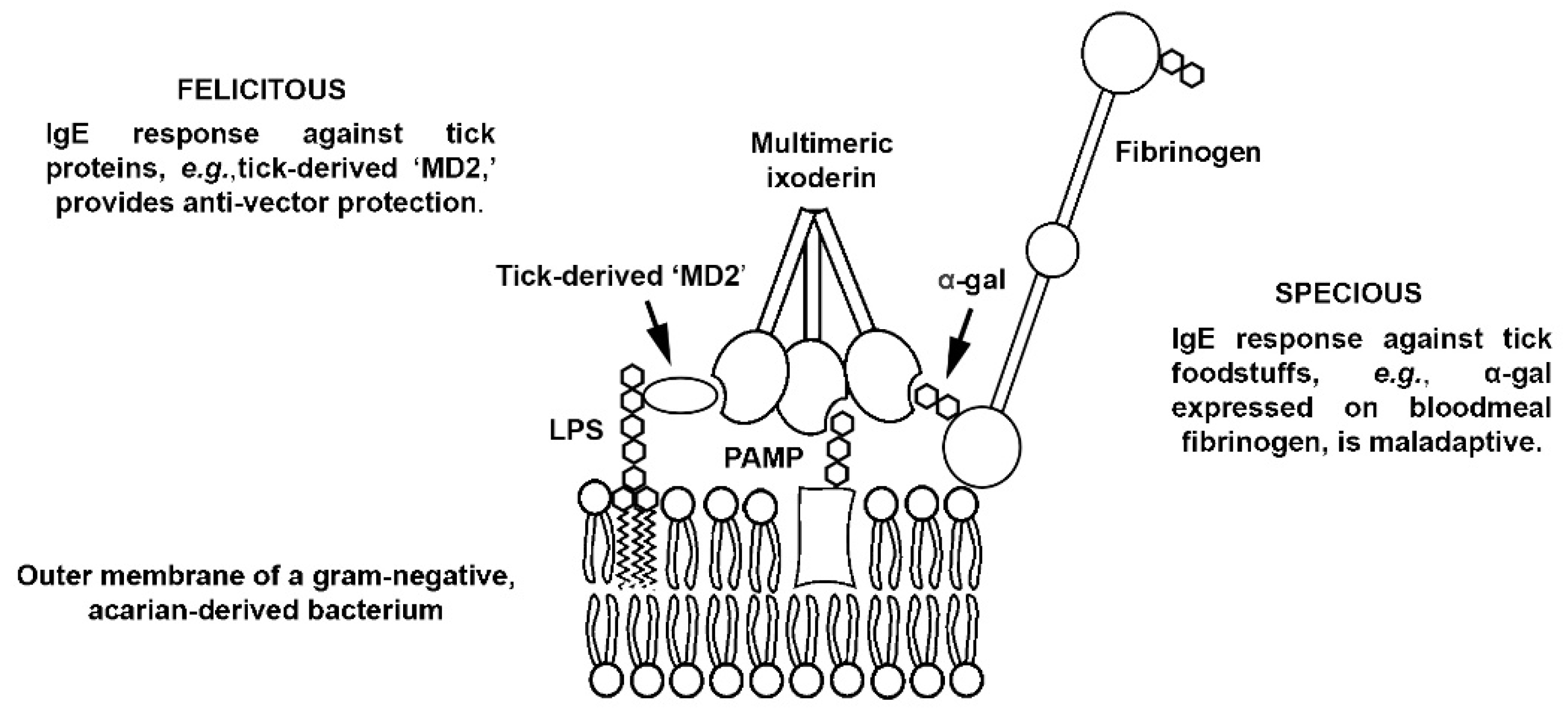
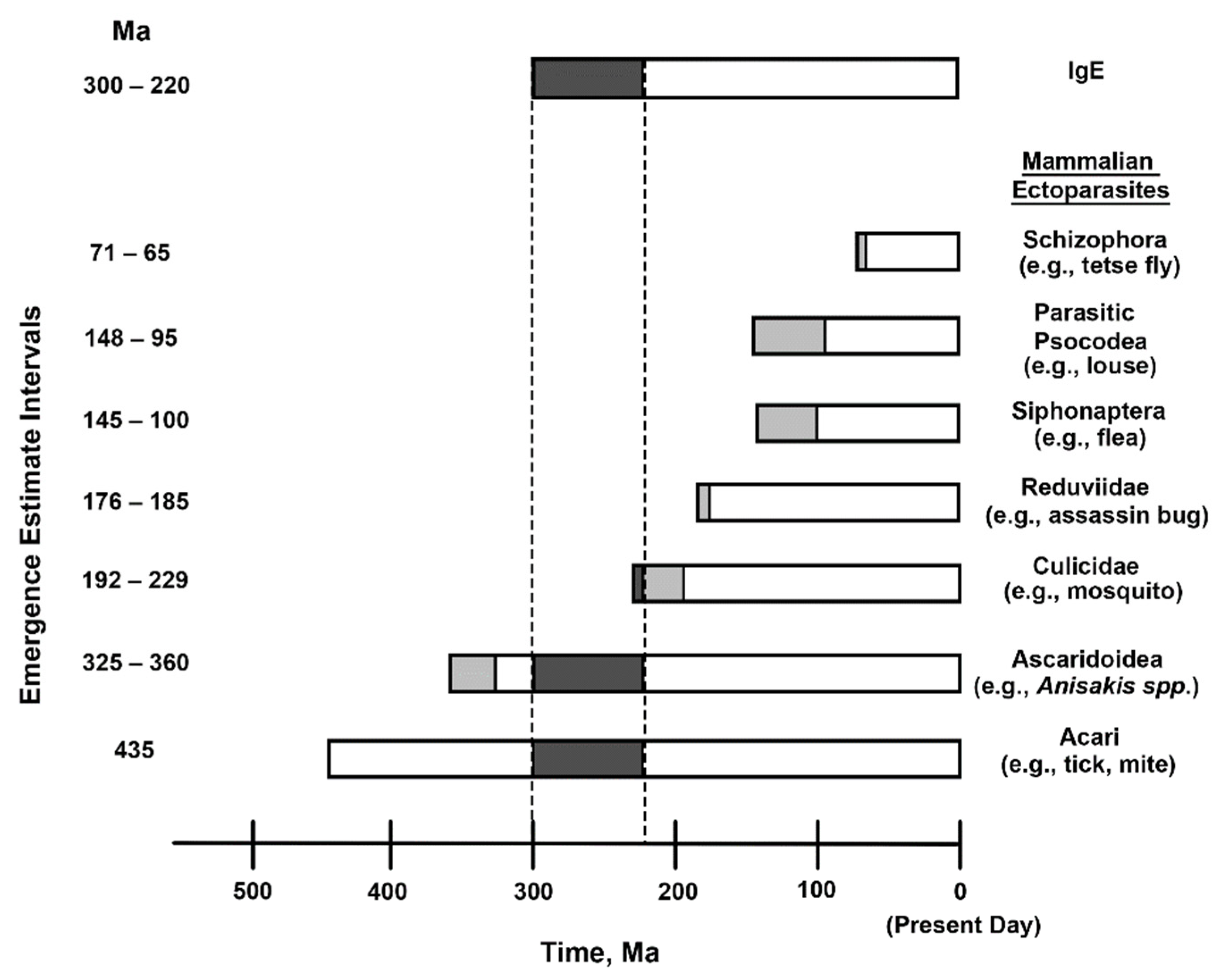
1. Background/Introduction
2. Discussion
3. Closing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Commins, S.P.; Jerath, M.R.; Cox, K.; Erickson, L.D.; Platts-Mills, T. Delayed anaphylaxis to alpha-gal, an oligosaccharide in mammalian meat. Allergol. Int. 2015, 65, 16–20. [Google Scholar] [CrossRef]
- Batard, T.; Baron-Bodo, V.; Martelet, A.; Le Mignon, M.; Lemoine, P.; Jain, K.; Mariano, S.; Horiot, S.; Chabre, H.; Harwanegg, C.; et al. Patterns of IgE sensitization in house dust mite-allergic patients: Implications for allergen immunotherapy. Allergy 2015, 71, 220–229. [Google Scholar] [CrossRef]
- Calderón, M.A.; Linneberg, A.; Kleine-Tebbe, J.; De Blay, F.; de Rojas, D.H.F.; Virchow, J.C.; Demoly, P. Respiratory allergy caused by house dust mites: What do we really know? J. Allergy Clin. Immunol. 2014, 136, 38–48. [Google Scholar] [CrossRef]
- Kim, C.-R.; Jeong, K.Y.; Yi, M.-H.; Kim, H.-P.; Shin, H.-J.; Yong, T.-S. Cross-reactivity between group-5 and -21 mite allergens from Dermatophagoides farinae, Tyrophagus putrescentiae and Blomia tropicalis. Mol. Med. Rep. 2015, 12, 5467–5474. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Nesvorna, M.; Klimov, P.; Dowd, S.E.; Sopko, B.; Erban, T. Differential allergen expression in three Tyrophagus putrescentiae strains inhabited by distinct microbiome. Allergy 2019, 74, 2502–2507. [Google Scholar] [CrossRef] [PubMed]
- Jakubas-Zawalska, J.; Asman, M.; Solarz, K. Sensitization to the storage mites Lepidoglyphus destructor and Tyrophagus pu-trescentiae (Acari, Sarcoptiformes, Astigmatina) in a suburban population in Southern Poland. Ann. Parasitol. 2017, 63, 183–188. [Google Scholar] [CrossRef]
- Matsumoto, T.; Hisano, T.; Hamaguchi, M.; Miike, T. Systemic Anaphylaxis after Eating Storage-Mite-Contaminated Food. Int. Arch. Allergy Immunol. 1996, 109, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Borges, M.; Capriles-Hulett, A.; Fernandez-Caldas, E.; Suarez-Chacon, R.; Caballero, F.; Castillo, S.; Sotillo, E. Mite-contaminated foods as a cause of anaphylaxis1. J. Allergy Clin. Immunol. 1997, 99, 738–743. [Google Scholar] [CrossRef]
- Johansson, E.; Kolmodin-Hedman, B.; Kallstrom, E.; Kaiser, L.; van Hage-Hamsten, M. IgE-mediated sensitization to predatory mites in Swedish greenhouse workers. Allergy 2003, 58, 337–341. [Google Scholar] [CrossRef]
- Spieksma, F.T.M. Domestic mites from an acarologic perspective. Allergy 1997, 52, 360–368. [Google Scholar] [CrossRef]
- Li, C.; Zhan, X.; Sun, E.; Zhao, J.; Wang, H.; He, J.; Wang, J. The density and species of mite breeding in stored products in China. Nutr. Hosp. 2014, 31, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Stejskal, V.; Munzbergová, Z.; Kubátová, A.; Váňová, M.; Žd’Árková, E. Mites and Fungi in Heavily Infested Stores in the Czech Republic. J. Econ. Èntomol. 2004, 97, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.R. Mites and Other Filth in Dried Shrimp Imported into the United States From the Orient. J. Food Prot. 1982, 45, 1204–1207. [Google Scholar] [CrossRef]
- Somerfield, K.G.; Manson, D.C.M.; Dale, P.S. Insects and mites associated with dried milk product storage areas in New Zealand. N. Z. J. Exp. Agric. 1980, 8, 83–85. [Google Scholar] [CrossRef]
- Karagoz, E.U.; Akdemir, C.; Direkel, S.; Guler, N.C. The Investigation of the Presence of Mites in Some Served Dry Foodstuffs. Turk. Parazitol. Derg. 2017, 41, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Vijayambika, V.; John, P.A. Internal morphology of the hypopus of Lardoglyphus konoi, a tyroglyphid pest on dried stored fish. Acarologia 1973, 15, 342–348. [Google Scholar]
- Gupta, A.; Sharma, Y.K. Human demodex mite: The versatile mite of dermatological importance. Indian J. Dermatol. 2014, 59, 302. [Google Scholar] [CrossRef]
- Retzinger, A.C.; Retzinger, G.S. Mites, ticks, anaphylaxis and allergy: The Acari hypothesis. Med. Hypotheses 2020, 144, 110257. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ding, Q.; Zhang, X. Ficolins and infectious diseases. Virol. Sin. 2014, 29, 25–32. [Google Scholar] [CrossRef]
- Sorrell, T.C.; Chen, S.C.A. Fungal-Derived Immune Modulating Molecules. Adv. Exp. Med. Biol. 2009, 666, 108–120. [Google Scholar] [CrossRef]
- Jun, J.-I.; Lau, L.F. CCN1 is an opsonin for bacterial clearance and a direct activator of Toll-like receptor signaling. Nat. Commun. 2020, 11, 1242. [Google Scholar] [CrossRef]
- Hajdušek, O.; Šíma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; De La Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Hanada, K.; Tsuji, N.; Igarashi, I.; Nagasawa, H.; Mikami, T.; Fujisaki, K. Characterization of Phagocytic Hemocytes in Ornithodoros moubata (Acari: Ixodidae). J. Med. Èntomol. 2001, 38, 514–519. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mondeková, H.H.; Sima, R.; Urbanová, V.; Kovar, V.; Rego, R.; Grubhoffer, L.; Kopacek, P.; Hajdusek, O. Characterization of Ixodes ricinus Fibrinogen-Related Proteins (Ixoderins) Discloses Their Function in the Tick Innate Immunity. Front. Cell. Infect. Microbiol. 2017, 7, 509. [Google Scholar] [CrossRef]
- Šimo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The Essential Role of Tick Salivary Glands and Saliva in Tick Feeding and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 281, Published 2017 Jun 22. [Google Scholar] [CrossRef]
- Tjernberg, I.; Hamsten, C.; Apostolovic, D.; Van Hage, M. IgE reactivity to α-Gal in relation to Lyme borreliosis. PLoS ONE 2017, 12, e0185723. [Google Scholar] [CrossRef]
- Rego, R.O.; Hajdusek, O.; Kovář, V.; Kopáček, P.; Grubhoffer, L.; Hypša, V. Molecular cloning and comparative analysis of fibrinogen-related proteins from the soft tick Ornithodoros moubata and the hard tick Ixodes ricinus. Insect Biochem. Mol. Biol. 2005, 35, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.A.; Galletti, M.F.B.D.M.; Ribeiro, J.M.; Fujita, A.; Costa, F.B.; Labruna, M.B.; Daffre, S.; Fogaça, A.C. The Distinct Transcriptional Response of the Midgut of Amblyomma sculptum and Amblyomma aureolatum Ticks to Rickettsia rickettsii Correlates to Their Differences in Susceptibility to Infection. Front. Cell. Infect. Microbiol. 2017, 7, 129. [Google Scholar] [CrossRef]
- Kovář, V.; Kopáček, P.; Grubhoffer, L. Isolation and characterization of Dorin M, a lectin from plasma of the soft tick Ornithodoros moubata. Insect Biochem. Mol. Biol. 2000, 30, 195–205. [Google Scholar] [CrossRef]
- Rego, R.; Kovar, V.; Kopacek, P.; Weise, C.; Man, P.; Sauman, I.; Grubhoffer, L. The tick plasma lectin, Dorin M, is a fibrinogen-related molecule. Insect Biochem. Mol. Biol. 2006, 36, 291–299. [Google Scholar] [CrossRef]
- Doolittle, R.F.; McNamara, K.; Lin, K. Correlating structure and function during the evolution of fibrinogen-related domains. Protein Sci. 2012, 21, 1808–1823. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. Variable immune molecules in invertebrates. J. Exp. Biol. 2013, 216, 4313–4319. [Google Scholar] [CrossRef]
- Hanington, P.C.; Zhang, S.-M. The Primary Role of Fibrinogen-Related Proteins in Invertebrates Is Defense, Not Coagulation. J. Innate Immun. 2011, 3, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Zuliani-Alvarez, L.; Marzeda, A.; Deligne, C.; Schwenzer, A.; McCann, F.E.; Marsden, B.D.; Piccinini, A.M.; Midwood, K.S. Mapping tenascin-C interaction with toll-like receptor 4 reveals a new subset of endogenous inflammatory triggers. Nat. Commun. 2017, 8, 1595. [Google Scholar] [CrossRef]
- Endo, Y.; Matsushita, M.; Fujita, T. New Insights into the Role of Ficolins in the Lectin Pathway of Innate Immunity. Int. Rev. Cell Mol. Biol. 2015, 316, 49–110. [Google Scholar] [CrossRef]
- Mörgelin, M.; Kasetty, G.; Olin, A.I.; Schmidtchen, A.; Herwald, H.; Påhlman, L.I. Antimicrobial activity of fibrinogen and fibrinogen-derived peptides—A novel link between coagulation and innate immunity. Thromb. Haemost. 2013, 109, 930–939. [Google Scholar] [CrossRef]
- Smiley, S.T.; King, J.A.; Hancock, W.W. Fibrinogen Stimulates Macrophage Chemokine Secretion Through Toll-Like Receptor 4. J. Immunol. 2001, 167, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Matsushita, M.; Fujita, T. The role of ficolins in the lectin pathway of innate immunity. Int. J. Biochem. Cell Biol. 2011, 43, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Bidula, S.; Sexton, D.W.; Schelenz, S. Ficolins and the Recognition of Pathogenic Microorganisms: An Overview of the Innate Immune Response and Contribution of Single Nucleotide Polymorphisms. J. Immunol. Res. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Casals, C.; García-Fojeda, B.; Minutti, C.M. Soluble defense collagens: Sweeping up immune threats. Mol. Immunol. 2019, 112, 291–304. [Google Scholar] [CrossRef]
- Grubhoffer, L.; Kovář, V.; Rudenko, N. Tick lectins: Structural and functional properties. Parasitology 2004, 129, S113–S125. [Google Scholar] [CrossRef] [PubMed]
- Md, A.; Maeda, M.; Matsui, T.; Takasato, Y.; Ito, K.; Kimura, Y. Purification and molecular characterization of a truncated-type Ara h 1, a major peanut allergen: Oligomer structure, antigenicity, and glycoform. Glycoconj. J. 2021, 38, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiao, H.; Zhou, P. Glycation by saccharides of different molecular sizes affected the allergenicity of shrimp tropomyosin via epitope loss and the generation of advanced glycation end products. Food Funct. 2019, 10, 7042–7051. [Google Scholar] [CrossRef] [PubMed]
- Altmann, F. The Role of Protein Glycosylation in Allergy. Int. Arch. Allergy Immunol. 2006, 142, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghouleh, A.; Johal, R.; Sharquie, I.K.; Emara, M.; Harrington, H.; Shakib, F.; Ghaemmaghami, A.M. The Glycosylation Pattern of Common Allergens: The Recognition and Uptake of Der p 1 by Epithelial and Dendritic Cells Is Carbohydrate Dependent. PLoS ONE 2012, 7, e33929. [Google Scholar] [CrossRef]
- Halim, A.; Carlsson, M.C.; Madsen, C.B.; Brand, S.; Möller, S.R.; Olsen, C.E.; Vakhrushev, S.; Brimnes, J.; Wurtzen, P.A.; Ipsen, H.; et al. Glycoproteomic Analysis of Seven Major Allergenic Proteins Reveals Novel Post-translational Modifications. Mol. Cell. Proteom. 2015, 14, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Batanero, E.; Villalba, M.; Rodríouez, R. Glycosylation site of the major allergen from olive tree pollen. Allergenic implications of the carbohydrate moiety. Mol. Immunol. 1994, 31, 31–37. [Google Scholar] [CrossRef]
- Trompette, A.; Divanovic, S.; Visintin, A.; Blanchard, C.; Hegde, R.S.; Madan, R.; Thorne, P.; Wills-Karp, M.; Gioannini, T.L.; Weiss, J.P.; et al. Allergenicity resulting from functional mimicry of a Toll-like receptor complex protein. Nature 2008, 457, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, R.; Akashi, S.; Ogata, H.; Nagai, Y.; Fukudome, K.; Miyake, K.; Kimoto, M. MD-2, a Molecule that Confers Lipopolysaccharide Responsiveness on Toll-like Receptor 4. J. Exp. Med. 1999, 189, 1777–1782. [Google Scholar] [CrossRef]
- Jerala, R. Structural biology of the LPS recognition. Int. J. Med. Microbiol. 2007, 297, 353–363. [Google Scholar] [CrossRef]
- Mueller, G.A.; Edwards, L.L.; Aloor, J.J.; Fessler, M.; Glesner, J.; Pomés, A.; Chapman, M.; London, R.E.; Pedersen, L. The structure of the dust mite allergen Der p 7 reveals similarities to innate immune proteins. J. Allergy Clin. Immunol. 2010, 125, 909–917.e4. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lü, L.; Nadler, S.A.; Gibson, D.I.; Zhang, L.-P.; Chen, H.-X.; Zhao, W.-T.; Guo, Y.-N. Molecular Phylogeny and Dating Reveal a Terrestrial Origin in the Early Carboniferous for Ascaridoid Nematodes. Syst. Biol. 2018, 67, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, A.C.M.; Azeredo-Espin, A.M.L.; Paulo, D.; Marinho, M.A.T.; Tomsho, L.P.; Drautz-Moses, D.I.; Purbojati, R.W.; Ratan, A.; Schuster, S.C. Large-scale mitogenomics enables insights into Schizophora (Diptera) radiation and population diversity. Sci. Rep. 2016, 6, 21762. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Hastriter, M.W.; Whiting, M.F.; Dittmar, K. Fleas (Siphonaptera) are Cretaceous, and evolved with Theria. Mol. Phylogenet. Evol. 2015, 90, 129–139. [Google Scholar] [CrossRef] [PubMed]
- de Moya, R.S.; Yoshizawa, K.; Walden, K.K.O.; Sweet, A.D.; Dietrich, C.H.; Johnson, K.P. Phylogenomics of Parasitic and Nonparasitic Lice (Insecta: Psocodea): Combining Sequence Data and Exploring Compositional Bias Solutions in Next Generation Data Sets. Syst. Biol. 2020, 70, 719–738. [Google Scholar] [CrossRef]
- Hwang, W.S.; Weirauch, C. Evolutionary History of Assassin Bugs (Insecta: Hemiptera: Reduviidae): Insights from Divergence Dating and Ancestral State Reconstruction. PLoS ONE 2012, 7, e45523. [Google Scholar] [CrossRef] [PubMed]
- Reidenbach, K.R.; Cook, S.; Bertone, M.A.; Harbach, R.E.; Wiegmann, B.M.; Besansky, N.J. Phylogenetic analysis and temporal diversification of mosquitoes (Diptera: Culicidae) based on nuclear genes and morphology. BMC Evol. Biol. 2009, 9, 298. [Google Scholar] [CrossRef] [PubMed]
- Dabert, M.; Witalinski, W.; Kaźmierski, A.; Olszanowski, Z.; Dabert, J. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. Mol. Phylogenet. Evol. 2010, 56, 222–241. [Google Scholar] [CrossRef] [PubMed]
- Hellman, L.T.; Akula, S.; Thorpe, M.; Fu, Z. Tracing the Origins of IgE, Mast Cells, and Allergies by Studies of Wild Animals. Front. Immunol. 2017, 8, 1749. [Google Scholar] [CrossRef]
- Morishima, R.; Motojima, S.; Tsuneishi, D.; Kimura, T.; Nakashita, T.; Fudouji, J.; Ichikawa, S.; Ito, H.; Nishino, H. Anisakis is a major cause of anaphylaxis in seaside areas: An epidemiological study in Japan. Allergy 2019, 75, 441–444. [Google Scholar] [CrossRef]
- Aibinu, I.; Smooker, P.; Lopata, A.L. Anisakis Nematodes in Fish and Shellfish- from infection to allergies. Int. J. Parasitol. Parasites Wildl. 2019, 9, 384–393. [Google Scholar] [CrossRef]
- Galli, S.J.; Metz, M.; Starkl, P.; Marichal, T.; Tsai, M. Mast cells and IgE in defense against lethality of venoms: Possible “benefit” of allergy. Allergo J. Int. 2020, 29, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Starkl, P.; Gaudenzio, N.; Marichal, T.; Reber, L.L.; Sibilano, R.; Watzenboeck, M.L.; Fontaine, F.; Mueller, A.C.; Tsai, M.; Knapp, S.; et al. IgE antibodies increase honeybee venom responsiveness and detoxification efficiency of mast cells. Allergy 2021. [Google Scholar] [CrossRef] [PubMed]
- Erban, T.; Harant, K.; Hubalek, M.; Vitamvas, P.; Kamler, M.; Poltronieri, P.; Tyl, J.; Markovic, M.; Titera, D. In-depth proteomic analysis of Varroa destructor: Detection of DWV-complex, ABPV, VdMLV and honeybee proteins in the mite. Sci. Rep. 2015, 5, srep13907. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Valdés, J.J. Are ticks venomous animals? Front. Zool. 2014, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Pienaar, R.; Neitz, A.W.H.; Mans, B.J. Tick Paralysis: Solving an Enigma. Vet. Sci. 2018, 5, 53. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein 1 | Sequence Length 2 | Identical Positions | Similar Positions | Global Identity, 2 % | Tissue Expression |
|---|---|---|---|---|---|
| Ficolin 1 (O00602) | 326 | 110 | 80 | 32 | Monocytes, lungs and spleen |
| Ixoderin A (I6LAP5) | 277 | Hemocytes and Malpighian tubule | |||
| Ficolin 2 (Q15485) | 313 | 100 | 83 | 21 | Liver (serum protein) |
| Ixoderin C (GCJO01000224) | 463 | Ubiquitous expression in all tissues; highest in gut and trachea | |||
| Ficolin 3 (O75636) | 299 | 71 | 89 | 21 | Gallbladder and lungs |
| Ixoderin B (Q5IUW6) | 286 | Salivary glands |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Retzinger, A.C.; Retzinger, G.S. The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors. Pathogens 2021, 10, 1220. https://doi.org/10.3390/pathogens10091220
Retzinger AC, Retzinger GS. The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors. Pathogens. 2021; 10(9):1220. https://doi.org/10.3390/pathogens10091220
Chicago/Turabian StyleRetzinger, Andrew C., and Gregory S. Retzinger. 2021. "The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors" Pathogens 10, no. 9: 1220. https://doi.org/10.3390/pathogens10091220
APA StyleRetzinger, A. C., & Retzinger, G. S. (2021). The Acari Hypothesis, II: Interspecies Operability of Pattern Recognition Receptors. Pathogens, 10(9), 1220. https://doi.org/10.3390/pathogens10091220