
What Babesia microti Is Now

Cummings School of Veterinary Medicine, Tufts University, Grafton, MA 01536, USA
Pathogens 2021, 10(9), 1168; https://doi.org/10.3390/pathogens10091168
Submission received: 11 August 2021
/
Revised: 1 September 2021
/
Accepted: 2 September 2021
/
Published: 10 September 2021
(This article belongs to the Special Issue Babesia and Human Babesiosis)
{kind=link}
Abstract
:Parasites from diverse hosts morphologically identified as Babesia microti have previously been shown to belong to a paraphyletic species complex. With a growing number of reports of B. microti-like parasites from across the world, this paper seeks to report on the current knowledge of the diversity of this species complex. Phylogenetic analysis of 18S rDNA sequences obtained from GenBank shows that the diversity of the B. microti species complex has markedly increased and now encompasses at least five distinct clades. This cryptic diversity calls into question much of our current knowledge of the life cycle of these parasites, as many biological studies were conducted before DNA sequencing technology was available. In many cases, it is uncertain which B. microti-like parasite was studied because parasites from different clades may occur sympatrically and even share the same host. Progress can only be made if future studies are conducted with careful attention to parasite identification and PCR primer specificity.
Babesia microti has historically been identified by the intra-erythrocytic morphology of the parasites, typically appearing in infected cells as small (1–2.0 µm in diameter) basket-shaped rings with extended chromatin. Relying solely on this morphology for identification, parasites from diverse hosts, such as shrews, mice, rats, raccoons, and dogs, are all referred to as B. microti or B. microti-like [1,2,3,4]. In 2003, a phylogenetic study was published demonstrating that such parasites are not a single organism. Sequences from both the 18S ribosomal and the beta-tubulin genes were paraphyletic, demonstrating that B. microti is, in fact, a species complex comprised of three genetically distinct clades [5]. Furthermore, parasites from only one of the clades are responsible for most human babesiosis cases. Indeed, cryptic diversity is common among parasites with few morphological traits that can be used for differentiation [6]. In the intervening 20 years, there has been a growing number of reports of B. microti-like parasites from diverse hosts worldwide. It is now clear that this species complex is even more diverse than originally described. This paper seeks to report the current knowledge of the species diversity and clarify, once again, what Babesia microti is.
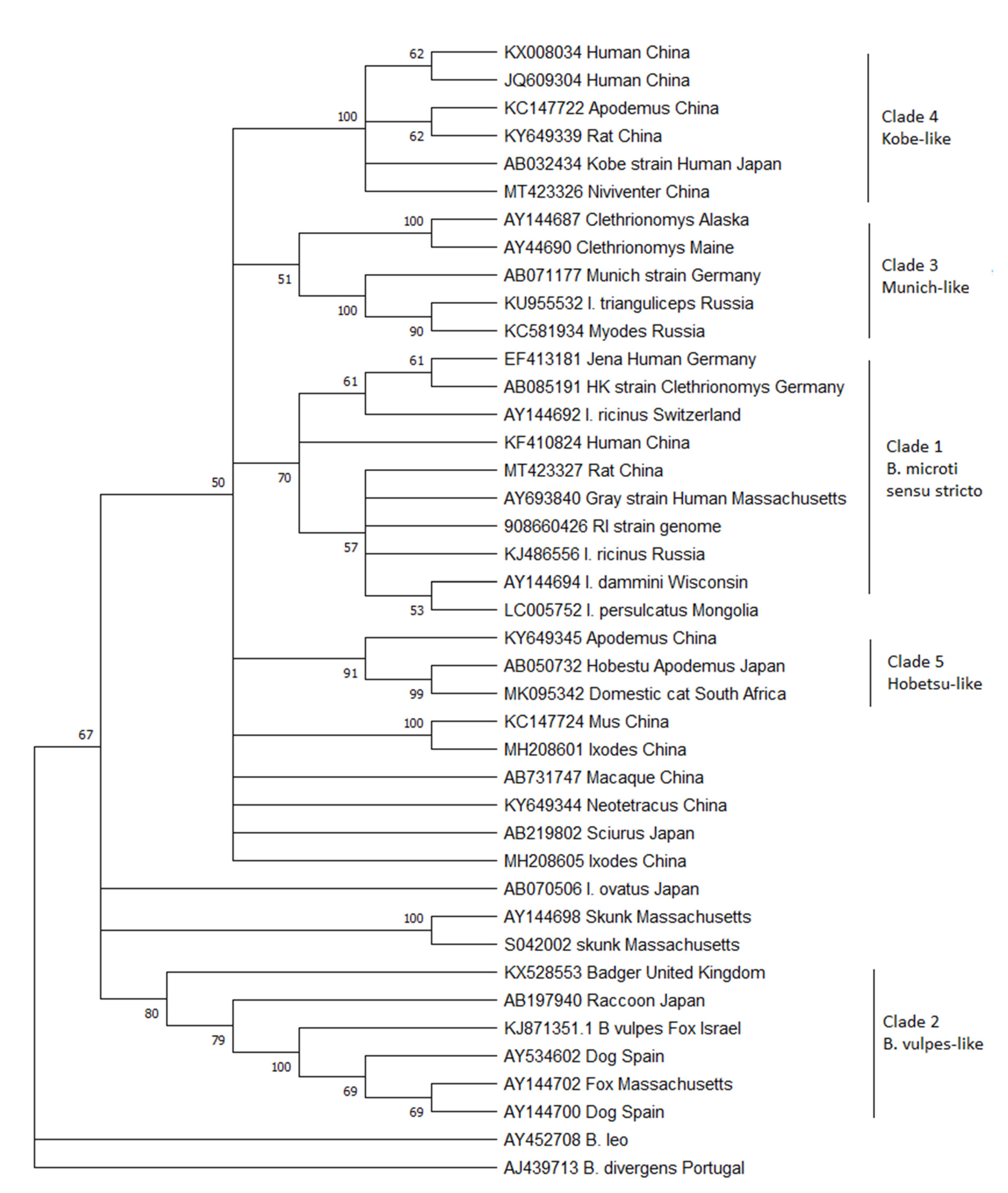
To this aim, all the 18S ribosomal DNA sequences >800 bp currently in GenBank that are either named Babesia microti in the database or appear on a Blast search with a sequence similarity of >95% to the B. microti human strains from the United States were downloaded. Although the 18S gene may not be the best target for describing diversity because of its highly conserved nature, it is the only gene that is reliably sequenced from the majority of studies. Limiting this analysis to large pieces of the gene maximizes the amount of diversity obtained from this conserved gene. From the large list of sequences available, a sample was chosen that attempted to encompass the genetic diversity of the entire database while removing large numbers of highly similar sequences that make the trees difficult to interpret. Thirty-nine sequences were aligned using Geneious (GenBank numbers are in the figure) and then trimmed so that they were all the same length, corresponding to bases 478–1350 from the Gray strain (GenBank #AY693840). A neighbor-joining tree was constructed with MEGA X [7] using B. divergens and B. leo as outgroups (Figure 1). This new analysis reveals that the three originally described clades remain, but they are joined by at least 2 additional clades.
Clade 1: Clade 1 has also been referred to as B. microti sensu stricto (or the US-type), as the parasites from this clade are arguably the most important because of their public health impact as the major cause of human babesiosis worldwide. This parasite is also the most studied. Clade 1 parasites are remarkably conserved, with virtually identical 18S rDNA sequences described across the globe: North America, Europe, and Asia. Despite this, human cases are only common in the United States. There, people are readily exposed because of the highly anthropophilic tick Ixodes dammini (the northern clade of Ixodes scapularis) that serves as its main vector [8,9]. In Europe, B. microti ss is thought to be maintained by I. trianguliceps, a host-specific tick that does not attack people, which would explain the lack of human disease. However, much of the work with I. trianculiceps was conducted before molecular methods were available, and it is uncertain whether the parasite under study belonged to this clade or clade 3 (For example see [3,10,11]). To date, no definitive molecular sequencing of field-derived I. trianguliceps has shown B. microti ss in these ticks. I. ricinus, an anthropophilic tick that serves as the vector for Lyme disease, is often sympatric, feeds on similar rodent reservoir hosts, and has been shown to be capable of transmitting piroplasms in the laboratory [12]. Indeed, B. microti ss is regularly detected in this vector, as well as I. persulcatus in Eurasia [13,14,15]. Therefore, the zoonotic potential should exist throughout the range of these ticks in Europe and Asia. Serosurveys show that tick-exposed people are indeed exposed to the parasite, but few cases of illness have been detected [16,17]. Whether the parasites are less virulent in the rest of the world compared to those in North America or whether physicians fail to diagnosis this disease because of lack of physician awareness and diagnostic capabilities has not been determined [18].
Clade 2: Clade 2 includes Babesia spp. that are known to infect carnivores, including raccoons, foxes, and badgers across the world. Also included in this clade is the parasite originally described from sick domestic dogs from Spain, which has been called by many names: B. microti-like, Babesia c.f. microti, Theileria annae, and Babesia annae [2,19]. Recently, it has been proposed that this parasite be designated a new species called Babesia vulpes [20,21]. The phylogenetic trees published by Baneth et al. in 2015 suggest that they propose the name B. vulpes only be applied to the Babesia in wild foxes, which also causes disease in domestic dogs, but not to the closely related Babesia found in other carnivores. The current phylogenetic analysis clearly shows that other sequences from raccoons and badgers group with B. vulpes in a strongly supported clade to the exclusion of the other B. microti-like parasites. If the name B. vulpes is indeed adopted, it seems unduly confusing to continue to refer to the other carnivore Babesia as B. microti-like. The vectors for the parasites in this clade have not been definitively established but are likely to be Ixodid ticks that feed primarily on carnivores, such as I. texanus in North America. In Europe, I. hexagonus and Dermacentor reticulatus have both been suggested as possible vectors [22,23]. It may also be transmitted by other routes that do not involve a vector, such as direct transmission through bites [24].
Not included within Clade 2 are the sequences from skunks originally described from Massachusetts [6]. The phylogenetic position of these skunk sequences is unstable, as they either cluster with the B. vulpes group, with the B. microti ss group, or, as is the case in this analysis, separated from both, depending on the type of algorithm used (see [6]). To date, no other similar sequences have been described, despite recent work in the U.S. characterizing small Babesia in medium-sized mammals [25,26,27], leaving our knowledge of this parasite limited and their placement among the B. microti clade uncertain. It is clear, however, that these piroplasms are distinct from the previously described Babesia from skunks, B. mephitis [28].
Clade 3: Clade 3 includes B. microti similar to the Munich strain that have been primarily detected from voles. These piroplasms occur in Europe and North America but have not been found in Asia. There is distinct separation in this clade between the sequences originating from the two continents. The Babesia from this clade are not known to cause human infection. In fact, they have been detected from areas of the US where human babesiosis has not been described and the anthropophilic vector, I. dammini, is not present [29,30]. Instead, I. angustus is known to occur in these areas, suggesting that this host-specific tick that rarely bites humans is the major vector in North America and the reason that this piroplasm is not known to infect humans. In Europe, the Munich type appears to be present only in areas where I. trianguliceps occurs, and it has been suggested that this tick is the primary vector [31]. However, throughout much of its range, either I. ricinus or I. persulcatus also occur, and as mentioned above, many ecological studies could not discern between the Munich-type and US-type parasites. Rar et al. [32] showed that in an area with sympatric I. trianguliceps and I. persulcatus, Munich-type B. microti was only detected in I. trianguliceps [32] and concluded that I. persulcatus was not the vector. However, others have detected sequences consistent with the Munich-type in I. ricinus [33,34,35], thus leaving the zoonotic potential for this piroplasm and the enzoonotic cycle in nature uncertain.
Clades 4 and 5: Clades 4 and 5 comprise Babesia that have only been detected in Asia. Clade 4 includes the Kobe strain from Japan together with sequences derived from China. Clade 5 comprises the Hobetsu and Otsu types, along with other sequences from voles from China. Although often referred to as if they are different parasites, these are actually the same and have 100% sequence similarity in the 18S rDNA gene (only Hobetsu is shown on the tree in Figure 1). None of the parasites from either clade have been detected in Europe or North America, though the US-type occurs sympatrically with both these parasites in Asia. Hobetsu parasites have been found primarily in Japan, with one report in rodents from mainland China [36,37], but Kobe appears to be more widespread in Japan, mainland China, and other parts of southeastern Asia and has been detected in more diverse rodent hosts [38,39,40]. In Japan, parasites from these two clades are often sympatric, but only the Kobe clade has been shown to infect humans. The Hobetsu strain has been detected in I. ovatus, and laboratory studies have confirmed the competence of this vector [37]. However, the vector for the Kobe strain remains undescribed; to date, it has never been detected in field-collected ticks. There is an odd report from a sick domestic cat in South Africa, which appeared to be coinfected with B. felis and B. microti-like parasites with 100% similarity to the Hobetsu strain [41]. This lone report remains an anomaly, as cats are not otherwise known to become infected with B. microti-like parasites, though they do harbor other small Babesia that are more closely related to B. rodhaini, B. leo, and B. felis [42].
Finally, there are a number of sequences from GenBank which fall within the B. microti species complex but do not group with other previously described parasites to create well-defined clades. The vast majority of these new sequences originate from rodents or ticks collected primarily in China but also Japan. Most remain as single reports or unpublished sequences deposited in GenBank, so little is known about their life cycles or their zoonotic potential. Interestingly, similar sequences were detected in squirrels collected in Japan and macaques from China [43,44]. Further investigations are necessary to characterize these parasites as well as create isolates. In the future, there will likely be 4 additional clades added to the B. microti species complex.
As this analysis shows, the parasites that are part of the B. microti species complex are a diverse group with unique life cycles. However, the understanding of the ecology of these parasites has been muddled because of the lack of precision in many studies and the confusion between similar parasites. As pointed out above, much of the basic biology of B. microti, both in the US and Europe, was conducted before molecular methods were available to distinguish between parasites. It is virtually impossible to know for certain which parasite, Clade 1 B. microti ss or Clade 3 Munich-type, was being studied in the older literature (for example, [45,46]), but also in more recent work (for example, [11,47,48]). In the United States in particular, researchers have focused their efforts on B. microti ss because of its public health importance there. Indeed, most studies are conducted in areas of the country where human cases are detected and presume the presence of that single parasite. This is likely to be an accurate assumption when surveying ticks. To date, B. microti ss is the only B. microti-like parasite found in the zoonotic vectors I. dammini and I. scapularis. Other Ixodes ticks that are more host-specific, such as I. cookei and I. angustus, are rarely studied. Surprisingly few studies in the U.S. have actually sequenced PCR amplicons obtained from wildlife sources to confirm the identity of B. microti unless the host is from an area where human babesiosis has not been detected. Unexpected results can arise when performing due diligence to confirm the identity of parasites (see [29,30] and the descriptions of B. conradae [49] and B. duncani [50]). Therefore, it is imperative at the start of any new study to confirm the identity of parasites by sequencing using a sufficiently informative gene segment (such as [30,51]). Once the specific clade of B. microti has been confirmed, it is not necessary to sequence every positive PCR given that the primers used are specific enough to amplify only the intended target. The use of non-specific PCR primers that are capable of amplifying other B. microti-like parasites can call into question the conclusions of a paper. Many different primer sets have been used in the literature, and a quick search using PrimerBlast from NCBI is useful to give a reader an estimate of their specificity. This issue becomes even more crucial with the use of real-time PCR, which usually amplifies small pieces of DNA that cannot be confirmed subsequently by sequencing.
It is clear from this analysis that there is much still unknown about the basic biology of the many parasites that make up the B. microti species complex. However, progress will only be obtained with well-designed studies that are careful to identify which B. microti-like parasite is being studied.
Funding
The author is supported by National Institutes of Health grants R01 AI 130105 and R01 AI 137424.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in the study is publicly available from GenBank. Accession numbers are included in the figure.
Conflicts of Interest
The author declares no conflict of interest.
References
- Spielman, A.; Etkind, P.; Piesman, J.; Ruebush, T.K.; Juranek, D.D.; Jacobs, M.S. Reservoir Hosts of Human Babesiosis on Nantucket Island. Am. J. Trop. Med. Hyg. 1981, 30, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.T.; Guitián, F.J.; Pallas, E.; Gestal, J.J.; Olmeda, A.S.; Goethert, H.K.; Telford, S.R. Infection of Dogs in North-West Spain with a Babesia Microti-like Agent. Vet. Rec. 2001, 149, 552–555. [Google Scholar] [CrossRef]
- Young, A.S. Investigations on the Epidemiology of Blood Parasites of Small Mammals with Special Reference to Piroplasms. Ph.D. Thesis, King’s College, London, UK, 1970. [Google Scholar]
- Healing, T.D. Infections with Blood Parasites in the Small British Rodents Apodemus Sylvaticus, Clethrionomys Glareolus and Microtus Agrestis. Parasitology 1981, 83, 179–189. [Google Scholar] [CrossRef]
- Goethert, H.K.; Telford III, S. What Is Babesia Microti? Parasitology 2003, 127, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, S.L.; Martinsen, E.S.; Falk, B.G. Do Molecules Matter More than Morphology? Promises and Pitfalls in Parasites. Parasitology 2011, 138, 1664–1674. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Spielman, A. Human Babesiosis on Nantucket Island: Transmission by Nymphal Ixodes Ticks. Am. J. Trop. Med. Hyg. 1976, 25, 784–787. [Google Scholar] [CrossRef]
- Spielman, A.; Wilson, M.L.; Levine, J.F.; Piesman, J. Ecology of Ixodes Dammini-Borne Human Babesiosis and Lyme Disease. Ann. Rev. Ent. 1985, 30, 439–460. [Google Scholar] [CrossRef]
- Krampitz, H.E. Babesia Microti: Morphology, Distribution and Host Relationship in Germany. Zentralbl. Bakteriol. Orig. A 1979, 244, 411–415. [Google Scholar]
- Randolph, S.E. The Effect of Babesia Microti on Feeding and Survival in Its Tick Vector, Ixodes Trianguliceps. Parasitology 1991, 102, 9–16. [Google Scholar] [CrossRef]
- Gray, J.; von Stedingk, L.V.; Gürtelschmid, M.; Granström, M. Transmission Studies of Babesia Microti in Ixodes Ricinus Ticks and Gerbils. J. Clin. Microbiol. 2002, 40, 1259–1263. [Google Scholar] [CrossRef] [Green Version]
- Duh, D.; Petrovec, M.; Avsic-Zupanc, T. Diversity of Babesia Infecting European Sheep Ticks (Ixodes Ricinus). J. Clin. Microbiol. 2001, 39, 3395–3397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foppa, I.M.; Krause, P.J.; Spielman, A.; Goethert, H.; Gern, L.; Brand, B.; Telford, S.R. Entomologic and Serologic Evidence of Zoonotic Transmission of Babesia Microti, Eastern Switzerland. Emerg. Infect. Dis. 2002, 8, 722–726. [Google Scholar] [CrossRef]
- Rar, V.A.; Epikhina, T.I.; Livanova, N.N.; Panov, V.V. Genetic Diversity of Babesia in Ixodes Persulcatus and Small Mammals from North Ural and West Siberia, Russia. Parasitology 2011, 138, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.-P.; Lambert, A.; Kampen, H.; Albert, S.; Epe, C.; Brade, V.; Tenter, A.M. Seroprevalence of Babesia Infections in Humans Exposed to Ticks in Midwestern Germany. J. Clin. Microbiol. 2002, 40, 2431–2436. [Google Scholar] [CrossRef] [Green Version]
- Wilhelmsson, P.; Lovmar, M.; Krogfelt, K.A.; Nielsen, H.V.; Forsberg, P.; Lindgren, P.E. Clinical/Serological Outcome in Humans Bitten by Babesia Species Positive Ixodes Ricinus Ticks in Sweden and on the Aland Islands. Ticks Tick Borne Dis. 2020, 11, 101455. [Google Scholar] [CrossRef]
- Hunfeld, K.-P.; Brade, V. Zoonotic Babesia: Possibly Emerging Pathogens to Be Considered for Tick-Infested Humans in Central Europe. Int. J. Med. Microbiol. Suppl. 2004, 293, 93–103. [Google Scholar] [CrossRef]
- Zahler, M.; Rinder, H.; Schein, E.; Gothe, R. Detection of a New Pathogenic Babesia Microti-like Species in Dogs. Vet. Parasitol. 2000, 89, 241–248. [Google Scholar] [CrossRef]
- Baneth, G.; Florin-Christensen, M.; Cardoso, L.; Schnittger, L. Reclassification of Theileria Annae as Babesia Vulpes Sp. Nov. Parasites Vectors 2015, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Baneth, G.; Cardoso, L.; Brilhante-Simões, P.; Schnittger, L. Establishment of Babesia Vulpes n. Sp. (Apicomplexa: Babesiidae), a Piroplasmid Species Pathogenic for Domestic Dogs. Parasites Vectors 2019, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, A.T.; Pallas, E.; Gestal, J.J.; Guitián, F.J.; Olmeda, A.S.; Telford, S.R.; Spielman, A. Ixodes Hexagonus Is the Main Candidate as Vector of Theileria Annae in Northwest Spain. Vet. Parasitol. 2003, 112, 157–163. [Google Scholar] [CrossRef]
- Hodžić, A.; Zörer, J.; Duscher, G.G. Dermacentor Reticulatus, a Putative Vector of Babesia Cf. Microti (Syn. Theileria Annae) Piroplasm. Parasitol. Res. 2017, 116, 1075–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeagley, T.J.; Reichard, M.V.; Hempstead, J.E.; Allen, K.E.; Parsons, L.M.; White, M.A.; Little, S.E.; Meinkoth, J.H. Detection of Babesia Gibsoni and the Canine Small Babesia ‘Spanish Isolate’ in Blood Samples Obtained from Dogs Confiscated from Dog Fighting Operations. J. Am. Vet. Med. Assoc. 2009, 235, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Savick, K.; Butler, J. Babesia Microti in Rodents and Raccoons from Northeast Florida. J. Parasitol. 2012, 98, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Garrett, K.B.; Hernandez, S.M.; Balsamo, G.; Barron, H.; Beasley, J.C.; Brown, J.D.; Cloherty, E.; Farid, H.; Gabriel, M.; Groves, B.; et al. Prevalence, Distribution, and Diversity of Cryptic Piroplasm Infections in Raccoons from Selected Areas of the United States and Canada. Int. J. Parasitol. Parasites Wildl. 2019, 9, 224–233. [Google Scholar] [CrossRef]
- Modarelli, J.J.; Westrich, B.J.; Milholland, M.; Tietjen, M.; Castro-Arellano, I.; Medina, R.F.; Esteve-Gasent, M.D. Prevalence of Protozoan Parasites in Small and Medium Mammals in Texas, USA. Int. J. Parasitol. Parasites Wildl. 2020, 11, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, A.A.; Frerichs, W.M. Babesia Mephitis Sp. n. (Protozoa: Piroplasmida), a Hematozoan Parasite of the Striped Skunk, Mephitis Mephitis. J. Parasitol. 1970, 56, 930–931. [Google Scholar] [CrossRef]
- Goethert, H.K.; Cook, J.A.; Lance, E.W.; Telford, S.R. Fay and Rausch 1969 Revisited: Babesia Microti in Alaskan Small Mammals. J. Parasitol. 2006, 92, 826–831. [Google Scholar] [CrossRef]
- Goethert, H.K.; Lubelcyzk, C.; LaCombe, E.; Holman, M.; Rand, P.; Smith, R.P., Jr.; Telford, S.R., III. Enzootic Babesia Microti in Maine. J. Parasitol. 2003, 89, 1069–1071. [Google Scholar] [CrossRef]
- Bown, K.J.; Lambin, X.; Telford, G.R.; Ogden, N.H.; Telfer, S.; Woldehiwet, Z.; Birtles, R.J. Relative Importance of Ixodes Ricinus and Ixodes Trianguliceps as Vectors for Anaplasma Phagocytophilum and Babesia Microti in Field Vole (Microtus Agrestis) Populations. Appl. Environ. Microbiol. 2008, 74, 7118–7125. [Google Scholar] [CrossRef] [Green Version]
- Rar, V.; Yakimenko, V.; Makenov, M.; Tikunov, A.; Epikhina, T.; Tancev, A.; Bobrova, O.; Tikunova, N. High Prevalence of Babesia Microti “Munich” Type in Small Mammals from an Ixodes Persulcatus/Ixodes Trianguliceps Sympatric Area in the Omsk Region, Russia. Parasitol. Res. 2016, 115, 3619–3629. [Google Scholar] [CrossRef]
- Pieniążek, N.; Sawczuk, M.; Skotarczak, B. Molecular Identification of Babesia Parasities Isolated from Ixodes Ricinus Ticks Collected in Northwestern Poland. Parasitology 2006, 92, 32–35. [Google Scholar] [CrossRef]
- Siński, E.; Bajer, A.; Welc, R.; Pawełczyk, A.; Ogrzewalska, M.; Behnke, J.M. Babesia Microti: Prevalence in Wild Rodents and Ixodes Ricinus Ticks from the Mazury Lakes District of North-Eastern Poland. Int. J. Med. Microbiol. 2006, 296, 137–143. [Google Scholar] [CrossRef]
- Welc-Falęciak, R.; Bajer, A.; Paziewska-Harris, A.; Baumann-Popczyk, A.; Siński, E. Diversity of Babesia in Ixodes Ricinus Ticks in Poland. Adv. Med. Sci. 2012, 57, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-R.; Ye, L.I.; Fan, J.-W.; Li, C.; Tang, F.; Liu, W.; Ren, L.-Z.; Bai, J.-Y. Detection of Kobe-Type and Otsu-Type Babesia Microti in Wild Rodents in China’s Yunnan Province. Epidemiol. Infect. 2017, 145, 2704–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamoto-Niikura, A.; Tsuji, M.; Qiang, W.; Nakao, M.; Hirata, H.; Ishihara, C. Detection of Two Zoonotic Babesia Microti Lineages, the Hobetsu and U.S. Lineages, in Two Sympatric Tick Species, Ixodes Ovatus and Ixodes Persulcatus, Respectively, in Japan. Appl. Environ. Microbiol. 2012, 78, 3424–3430. [Google Scholar] [CrossRef]
- Saito-Ito, A.; Takada, N.; Ishiguro, F.; Fujita, H.; Yano, Y.; Ma, X.-H.; Chen, E.-R. Detection of Kobe-Type Babesia Microti Associated with Japanese Human Babesiosis in Field Rodents in Central Taiwan and Southeastern Mainland China. Parasitology 2008, 135, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Saito-Ito, A.; Kasahara, M.; Kasai, M.; Dantrakool, A.; Kawai, A.; Fujita, H.; Yano, Y.; Kawabata, H.; Takada, N. Survey of Babesia Microti Infection in Field Rodents in Japan: Records of the Kobe-Type in New Foci and Findings of a New Type Related to the Otsu-Type. Microbiol. Immunol. 2007, 51, 15–24. [Google Scholar] [CrossRef]
- Tsuji, M.; Wei, Q.; Zamoto, A.; Morita, C.; Arai, S.; Shiota, T.; Fujimagari, M.; Itagaki, A.; Fujita, H.; Ishihara, C. Human Babesiosis in Japan: Epizootiologic Survey of Rodent Reservoir and Isolation of New Type of Babesia Microti-Like Parasite. J. Clin. Microbiol. 2001, 39, 4316–4322. [Google Scholar] [CrossRef] [Green Version]
- Bosman, A.-M.; Penzhorn, B.L.; Brayton, K.A.; Schoeman, T.; Oosthuizen, M.C. A Novel Babesia Sp. Associated with Clinical Signs of Babesiosis in Domestic Cats in South Africa. Parasites Vectors 2019, 12, 138. [Google Scholar] [CrossRef]
- Penzhorn, B.L.; Oosthuizen, M.C. Babesia Species of Domestic Cats: Molecular Characterization Has Opened Pandora’s Box. Front. Vet. Sci. 2020, 7, 134. [Google Scholar] [CrossRef]
- Tsuji, M.; Zamoto, A.; Kawabuchi, T.; Kataoka, T.; Nakajima, R.; Asakawa, M.; Ishihara, C. Babesia Microti-Like Parasites Detected in Eurasian Red Squirrels (Sciurus Vulgaris OriEnt.is) in Hokkaido, Japan. J. Vet. Med. Sci. 2006, 68, 643–646. [Google Scholar] [CrossRef] [Green Version]
- Voorberg-vd Wel, A.; Kocken, C.H.M.; Zeeman, A.-M.; Thomas, A.W. Detection of New Babesia Microti-like Parasites in a Rhesus Monkey (Macaca Mulatta) with a Suppressed Plasmodium Cynomolgi Infection. Am. J. Trop. Med. Hyg. 2008, 78, 643–645. [Google Scholar]
- Coles, A. Blood Parasites Found in Mammals, Birds and Fishes in England. Parasitology 1914, 7, 17–61. [Google Scholar] [CrossRef]
- Franca, C. Sur Une Piroplasme Nouvelle Chez Une Mangouste. Bull. Soc. Pathol. Exot. 1908, 1, 410. [Google Scholar]
- Anderson, J.F.; Johnson, R.C.; Magnarelli, L.A.; Hyde, F.W.; Myers, J.E. Peromyscus Leucopus and Microtus Pennsylvanicus Simultaneously Infected with Borrelia Burgdorferi and Babesia Microti. J. Clin. Microbiol. 1986, 23, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.F.; Magnarelli, L.A.; Kurz, J. Intraerythrocytic Parasites in Rodent Populations of Connecticut: Babesia and Grahamella Species. J. Parasitol. 1979, 65, 599–604. [Google Scholar] [CrossRef]
- Kjemtrup, A.M.; Wainwright, K.; Miller, M.; Penzhorn, B.L.; Carreno, R.A. Babesia Conradae, Sp. Nov., a Small Canine Babesia Identified in California. Vet. Parasitol. 2006, 138, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Conrad, P.A.; Kjemtrup, A.M.; Carreno, R.A.; Thomford, J.; Wainwright, K.; Eberhard, M.; Quick, R.; Telford III, S.R.; Herwaldt, B.L. Description of Babesia Duncani n.Sp. (Apicomplexa: Babesiidae) from Humans and Its Differentiation from Other Piroplasms. Int. J. Parasitol. 2006, 36, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Katavolos, P.; Caporale, D.A.; Smith, R.P.; Spielman, A.; Telford, S.R. Diversity of Babesia Infecting Deer Ticks (Ixodes Dammini). Am. J. Trop. Med. Hyg. 1998, 58, 739–742. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of the 18S rDNA gene of B. microti-like piroplasms. A neighbor-joining tree was constructed using MEGA X with 500 bootstrap replicates. Evolutionary distances were calculated using the Kimera-2 parameter method with B. divergens and B. leo as outgroups. Branches with less than 50% bootstrap support were consolidated. GenBank accession numbers are listed on the tree.
Figure 1.
Phylogenetic analysis of the 18S rDNA gene of B. microti-like piroplasms. A neighbor-joining tree was constructed using MEGA X with 500 bootstrap replicates. Evolutionary distances were calculated using the Kimera-2 parameter method with B. divergens and B. leo as outgroups. Branches with less than 50% bootstrap support were consolidated. GenBank accession numbers are listed on the tree.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Goethert, H.K. What Babesia microti Is Now. Pathogens 2021, 10, 1168. https://doi.org/10.3390/pathogens10091168
AMA Style
Goethert HK. What Babesia microti Is Now. Pathogens. 2021; 10(9):1168. https://doi.org/10.3390/pathogens10091168
Chicago/Turabian StyleGoethert, Heidi K. 2021. "What Babesia microti Is Now" Pathogens 10, no. 9: 1168. https://doi.org/10.3390/pathogens10091168
APA StyleGoethert, H. K. (2021). What Babesia microti Is Now. Pathogens, 10(9), 1168. https://doi.org/10.3390/pathogens10091168
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.