
Amblyomma aureolatum Genetic Diversity and Population Dynamics Are Not Related to Spotted Fever Epidemiological Scenarios in Brazil

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Molecular Identification of Ticks
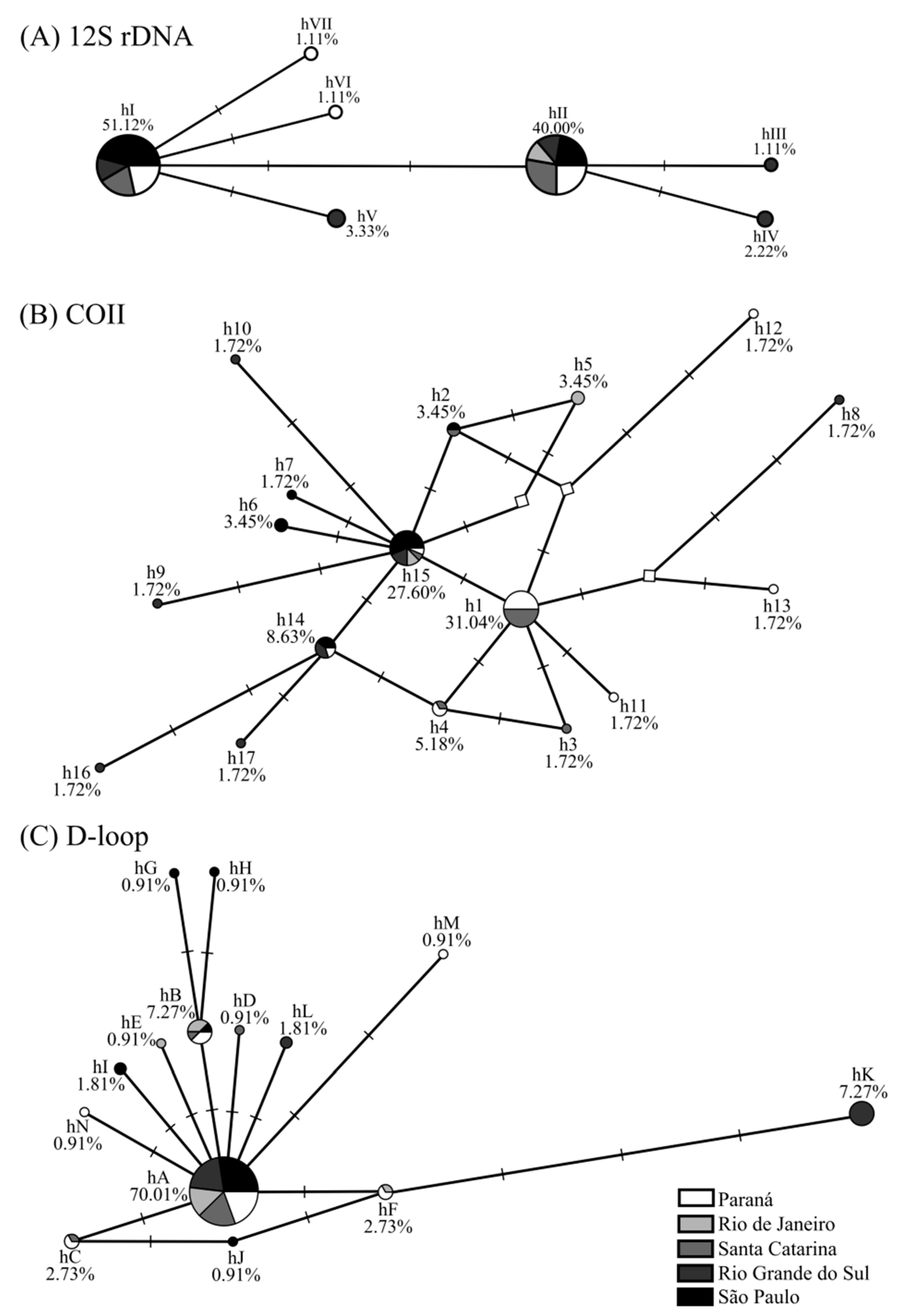
2.2. Intrapopulational Analysis: A. aureolatum Has Unique Haplotypes per Population and Low Nucleotide Diversity
2.3. Population Differentiation, Structuring, and Demographic Dynamics in A. aureolatum from Brazilian Spotted Fever Scenarios
2.4. Identification of Detected Rickettsiae
3. Discussion
4. Materials and Methods
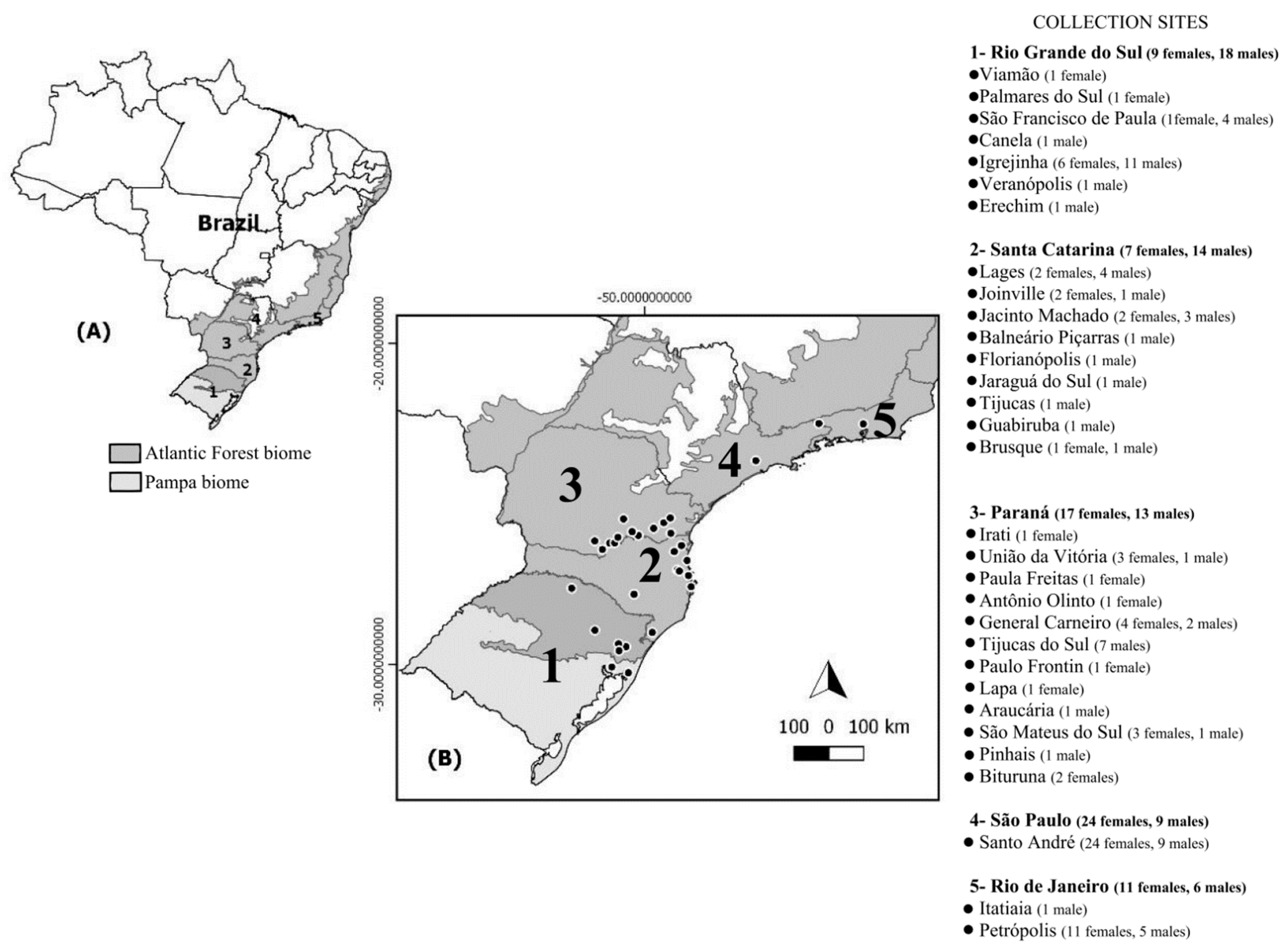
4.1. Sampling and Taxonomy
4.2. DNA Extraction, Amplification, and Sequencing
4.3. Sequence Editing, Identification, and Alignment
4.4. Amblyomma aureolatum Population Genetics Analysis
4.5. Phylogenetic Relationships of Detected Rickettsiae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szabó, M.P.J.; Pinter, A.; Labruna, M.B. Ecology, biology and distribution of spotted fever tick vectors in Brazil. Front. Cell. Infect. Microbiol. 2013, 27, 27. [Google Scholar] [CrossRef] [Green Version]
- Guglielmone, A.A.; Estrada-Peña, A.; Mangold, A.J.; Barros-Battesti, D.M.; Labruna, M.B.; Martins, J.R.; Venzal, J.M.; Arzua, M.; Keirans, J.E. Amblyomma aureolatum (Pallas, 1772) and Amblyomma ovale Kock, 1844: Hosts, distribution and 16S rDNA sequences. Vet. Parasitol. 2003, 113, 273–288. [Google Scholar] [CrossRef]
- Pinter, A.; Dias, R.A.; Gennari, S.M.; Labruna, M.B. Study of the seasonal dynamics, life cycle, and host specificity of Amblyomma aureolatum (Acari: Ixodidae). J. Med. Entomol. 2004, 41, 324–332. [Google Scholar] [CrossRef]
- Dall’Agnol, B.; Souza, U.A.; Weck, B.; Trigo, T.C.; Jardim, M.M.A.; Costa, F.B.; Labruna, M.B.; Peters, F.B.; Favarini, M.O.; Mazim, F.D.; et al. Rickettsia parkeri in free-ranging wild canids from Brazilian Pampa. Transboundary Emerg. Dis. 2018, 65, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Reck, J.; Souza, U.; Souza, G.; Kieling, E.; Dall’Agnol, B.; Webster, A.; Michel, T.; Doyle, R.; Martins, T.F.; Labruna, M.B.; et al. Records of ticks on humans in Rio Grande do Sul state, Brazil. Ticks Tick Borne Dis. 2018, 9, 1296–1301. [Google Scholar] [CrossRef]
- Souza, U.A.; Webster, A.; Dall’Agnol, B.; Peters, F.B.; Favarini, M.O.; Schott, D.; Zitelli, L.C.; Mazim, F.D.; Kasper, C.B.; Ott, R.; et al. Ticks, mites, fleas, and vector-borne pathogens in free-ranging neotropical wild felids from southern Brazil. Ticks Tick Borne Dis. 2021, 12, 101706. [Google Scholar] [CrossRef] [PubMed]
- Ogrzewalska, M.; Saraiva, D.G.; Moraes-Filho, J.; Martins, T.F.; Costa, F.B.; Pinter, A.; Labruna, M.B. Epidemiology of Brazilian spotted fever in the Atlantic Forest, state of São Paulo, Brazil. Parasitology 2012, 139, 1283–1300. [Google Scholar] [CrossRef]
- Scinachi, C.A.; Takeda, G.A.; Mucci, L.F.; Pinter, A. Association of the occurrence of Brazilian spotted fever and Atlantic rain forest fragmentation in the São Paulo metropolitan region, Brazil. Acta Trop. 2017, 166, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Gray, J. Biology of Ixodes species ticks in relation to tick-borne zoonoses. Wien Klin Wochenschr. 2002, 114, 473–478. [Google Scholar] [PubMed]
- Araya-Anchetta, A.; Busch, J.D.; Scoles, G.A.; Wagner, D.M. Thirty years of tick population genetics: A comprehensive review. Infect. Genet. Evol. 2015, 29, 164–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitencourth, K.; Voloch, C.M.; Serra-Freire, N.M.; Machado-Ferreira, E.; Amorim, M.; Gazêta, G.S. Analysis of Amblyomma sculptum haplotypes in an area endemic for Brazilian spotted fever. Med. Vet. Entomol. 2016, 30, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Bitencourth, K.; Amorim, M.; Oliveira, S.V.; Caetano, R.L.; Voloch, C.M.; Gazêta, G.S. Amblyomma sculptum: Genetic diversity and rickettsias in the Brazilian Cerrado biome. Med. Vet. Entomol. 2017, 31, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Bitencourth, K.; Amorim, M.; Oliveira, S.V.; Voloch, C.M.; Gazêta, G.S. Genetic diversity, population structure and rickettsias in Amblyomma ovale in areas of epidemiological interest for spotted fever in Brazil. Med. Vet. Entomol. 2019, 33, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Ogrzewalska, M.; Schwarcz, K.; Bajay, M.M.; Bajay, S.K.; Pinheiro, J.B.; Zucchi, M.I.; Pinter, A.; Labruna, M.B. Characterization of genetic variability and population structure of the tick Amblyomma aureolatum (Acari: Ixodidae). J. Med. Entomol. 2016, 53, 843–850. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Saraiva, D.G.; Soares, H.S.; Soares, J.F.; Labruna, M.B. Feeding period required by Amblyomma aureolatum ticks for transmission of Rickettsia rickettsii to vertebrate hosts. Emerg. Infect. Dis. 2014, 20, 1504–1510. [Google Scholar] [CrossRef]
- Labruna, M.B.; Ogrzewalska, M.; Martins, T.F.; Pinter, A.; Horta, M.C. Comparative susceptibility of larval stages of Amblyomma aureolatum, Amblyomma cajennense, and Rhipicephalus sanguineus to infection by Rickettsia rickettsii. J. Med. Entomol. 2008, 45, 1156–1159. [Google Scholar] [CrossRef]
- Pinter, A.; Labruna, M.B. Isolation of Rickettsia rickettsii and Rickettsia bellii in cell culture from the tick Amblyomma aureolatum in Brazil. Ann. N. Y. Acad. Sci. 2006, 1078, 523–529. [Google Scholar] [CrossRef]
- Luz, H.R.; Faccini, J.L.H.; McIntosh, D. Molecular analyses reveal an abundant diversity of ticks and rickettsial agents associated with wild birds in two regions of primary Brazilian Atlantic Rainforest. Ticks Tick Borne Dis. 2017, 8, 657–665. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Hayes, S.F.; Mavros, A.J. Non pathogenic rickettsiae in Dermacentor andersoni: A limiting factor for the distribution of Rickettsia rickettsii. In Rickettsiae and Rickettsial Diseases; Burgdorfer, W., Anacker, R.L., Eds.; Academic Press: New York, NY, USA, 1981; pp. 585–594. [Google Scholar]
- Sakai, R.K.; Costa, F.B.; Ueno, T.E.; Ramirez, D.G.; Soares, J.F.; Fonseca, A.H.; Labruna, M.B.; Barros-Battesti, D.M. Experimental infection with Rickettsia rickettsii in an Amblyomma dubitatum tick colony, naturally infected by Rickettsia bellii. Ticks Tick Borne Dis. 2014, 5, 917–923. [Google Scholar] [CrossRef]
- Raoult, D.; La Scola, B.; Enea, M.; Fournier, P.E.; Roux, V.; Fenollar, F.; Galvão, M.A.; Lamballerie, X. A flea-associated Rickettsia pathogenic for humans. Emerg. Infect. Dis. 2001, 7, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Barros-Battesti, D.M.; Arzua, M.; Bechara, G.H. Carrapatos de Importância Médico- Veterinária da Região Neotropical: Um guia Ilustrado para Identificação de Espécies; Butantan: São Paulo, Brazil, 2006; 223p. [Google Scholar]
- Medeiros, A.P.; Souza, A.P.; Moura, A.B.; Lavina, M.S.; Bellato, V.; Sartor, A.A.; Nieri-Bastos, F.A.; Richtzenhain, L.J.; Labrina, M.B. Spotted fever group Rickettsia infecting ticks (Acari: Ixodidae) in the state of Santa Catarina, Brazil. Mem. Inst. Oswaldo Cruz 2011, 106, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR- based techniques. Nucleic Acids Res. 1997, 15, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Beati, L.; Keirans, J.E. Analysis of the systematic relationships among ticks of the genera Rhipicephalus and Boophilus (Acari: Ixodidae) based on mitochondrial 12S ribossomal DNA gene sequences and morphological characters. J. Parasitol. 2001, 87, 32–48. [Google Scholar] [CrossRef]
- Burkman, E.J. Genetic structure of Amblyomma cajennense (Acari: Ixodidae) populations based on mitochondrial gene sequences. Master’s Thesis, Georgia Southern University, Statesboro, GA, USA, 2009. [Google Scholar]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of São Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic identification of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (OmpB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labruna, M.B.; McBride, J.W.; Bouyer, D.H.; Camargo, L.M.A.; Camargo, E.P.; Walker, D.H. Molecular evidence for a spotted fever group Rickettsia species in the tick Amblyomma longirostre in Brazil. J. Med. Entomol. 2004, 41, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.; Mitchell, C.; Malloy, D.C.; Dasch, G.A.; Azad, A.F. Detection of murine typhus infection in fleas by using the polymerase chain reaction. J. Clin. Microbiol. 1990, 28, 530–534. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Onsins, S.E.; Rozas, J. Statistical properties of new neutrality tests against population growth. Mol. Biol. Evol. 2002, 19, 2092–2100. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-Del Barrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Gene/Region | Epidemiological Scenario | Population | Sample Size | n | s | k ± SD | h ± SD | nd ± SD | Fu’s Fs (p-Value) | Tajima’s D (p-Value) | R2 (p-Value) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 12S rDNA | Group I | Paraná | 21 | 4 | 4 | 1.22 ± 0.81 | 0.614 ± 0.062 | 0.0033 ± 0.0024 | 0.516 (0.62) | 0.275 (0.65) | 0.148 (0.56) |
| Santa Catarina | 19 | 2 | 2 | 1.06 ± 0.73 | 0.526 ± 0.040 | 0.0028 ± 0.0022 | 3.031 (0.96) | 2.006 (0.99) | 0.263 (1.00) | ||
| Rio de Janeiro | 4 | 1 | 0 | 0.00 ± 0.00 | 0.000 ± 0.000 | 0.0000 ± 0.0000 | N.A. | 0.000 (1.00) | N.A. | ||
| Rio Grande do Sul | 17 | 5 | 5 | 1.71 ± 1.05 | 0.787 ± 0.059 | 0.0046 ± 0.0032 | 0.007 (0.53) | 0.495 (0.71) | 0.166 (0.68) | ||
| Group II | São Paulo | 29 | 2 | 2 | 0.83 ± 0.61 | 0.414 ± 0.077 | 0.0022 ± 0.0018 | 2.787 (0.95) | 1.293 (0.90) | 0.207 (0.93) | |
| COII | Group I | Paraná | 16 | 7 | 8 | 1.31 ± 0.86 | 0.718 ± 0.128 | 0.0027 ± 0.0020 | −3.113 (0.00) * | −1.649 (0.04) * | 0.113 (0.06) |
| Santa Catarina | 13 | 5 | 3 | 0.73 ± 0.58 | 0.533 ± 0.142 | 0.0015 ± 0.0013 | −2.582 (0.01) * | −1.471 (0.03) * | 0.147 (0.17) | ||
| Rio de Janeiro | 4 | 2 | 2 | 1.34 ± 1.03 | 0.667 ± 0.204 | 0.0027 ± 0.0025 | 1.530 (0.86) | 1.893 (0.95) | 0.333 (0.44) | ||
| Rio Grande do Sul | 10 | 7 | 10 | 2.35 ± 1.39 | 0.911 ± 0.077 | 0.0048 ± 0.0032 | −2.785 (0.02) | −1.507 (0.09) | 0.098 (0.00) * | ||
| Group II | São Paulo | 15 | 5 | 4 | 0.76 ± 0.59 | 0.638 ± 0.129 | 0.0016 ± 0.0014 | −2.233 (0.07) | −1.220 (0.08) | 0.105 (0.00) * | |
| D-loop | Group I | Paraná | 24 | 6 | 6 | 0.80 ± 0.60 | 0.601 ± 0.108 | 0.0017 ± 0.0015 | −2.678 (0.01) * | −1.524 (0.03) * | 0.086 (0.06) |
| Santa Catarina | 17 | 4 | 3 | 0.35 ± 0.36 | 0.331 ± 0.143 | 0.0007 ± 0.0009 | −2.527 (0.00) * | −1.706 (0.01) * | 0.127 (014) | ||
| Rio de Janeiro | 16 | 4 | 3 | 0.58 ± 0.49 | 0.517 ± 0.132 | 0.0013 ± 0.0012 | −1.478 (0.06) | −1.055 (0.16) | 0.123 (0.11) | ||
| Rio Grande do Sul | 26 | 3 | 5 | 1.93 ± 1.13 | 0.542 ± 0.075 | 0.0042 ± 0.0028 | 3.520 (0.97) | 1.319 (0.87) | 0.192 (0.92) | ||
| Group II | São Paulo | 27 | 6 | 6 | 0.65 ± 0.52 | 0.399 ± 0.117 | 0.0014 ± 0.0013 | −3.205 (0.01) * | −1.718 (0.01) * | 0.080 (0.08) |
| Population | Marker | Paraná | Santa Catarina | Rio de Janeiro | Rio Grande do Sul |
|---|---|---|---|---|---|
| Santa Catarina | 12S rDNA | −0.03027 | |||
| COII | −0.04289 | ||||
| D-loop | −0.01711 | ||||
| Rio de Janeiro | 12S rDNA | 0.32037 | 0.25709 | ||
| COII | 0.43171 * | 0.54683 * | |||
| D-loop | −0.02385 | 0.00731 | |||
| Rio Grande do Sul | 12S rDNA | −0.01408 | −0.01599 | 0.21181 | |
| COII | 0.31469 * | 0.35290 * | 0.12080 | ||
| D-loop | 0.18499 * | 0.20231 * | 0.18353 * | ||
| São Paulo ♦ | 12S rDNA | 0.01039 | 0.08547 | 0.55767 * | 0.05795 |
| COII | 0.41611* | 0.48016 * | 0.28403 * | 0.05003 | |
| D-loop | −0.01457 | −0.01774 | −0.01781 | 0.21033 * |
| Population | Marker | Group II |
|---|---|---|
| Group I | 12S rDNA | 0.06858 * |
| COII | 0.17215 * | |
| D-loop | 0.02085 |
| Analyzed Marker | |||||||
|---|---|---|---|---|---|---|---|
| AMOVA | (Φ Statistic) | 12S rDNA | COII | D-Loop | |||
| Among groups | (ΦCT) | 3.80% | (0.038) | −1.37% | (−0.014) | −9.14% | (−0.091) |
| Among populations within groups | (ΦSC) | 4.88% | (0.051) | 31.36% | (0.309 *) | 18.30% | (0.168 *) |
| Within populations | (ΦST) | 91.32% | (0.087*) | 70.01% | (0.300 *) | 90.84% | (0.092 *) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitencourth, K.; Amorim, M.; Oliveira, S.V.d.; Gazêta, G.S. Amblyomma aureolatum Genetic Diversity and Population Dynamics Are Not Related to Spotted Fever Epidemiological Scenarios in Brazil. Pathogens 2021, 10, 1146. https://doi.org/10.3390/pathogens10091146
Bitencourth K, Amorim M, Oliveira SVd, Gazêta GS. Amblyomma aureolatum Genetic Diversity and Population Dynamics Are Not Related to Spotted Fever Epidemiological Scenarios in Brazil. Pathogens. 2021; 10(9):1146. https://doi.org/10.3390/pathogens10091146
Chicago/Turabian StyleBitencourth, Karla, Marinete Amorim, Stefan Vilges de Oliveira, and Gilberto Salles Gazêta. 2021. "Amblyomma aureolatum Genetic Diversity and Population Dynamics Are Not Related to Spotted Fever Epidemiological Scenarios in Brazil" Pathogens 10, no. 9: 1146. https://doi.org/10.3390/pathogens10091146
APA StyleBitencourth, K., Amorim, M., Oliveira, S. V. d., & Gazêta, G. S. (2021). Amblyomma aureolatum Genetic Diversity and Population Dynamics Are Not Related to Spotted Fever Epidemiological Scenarios in Brazil. Pathogens, 10(9), 1146. https://doi.org/10.3390/pathogens10091146