
Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Field Symptoms
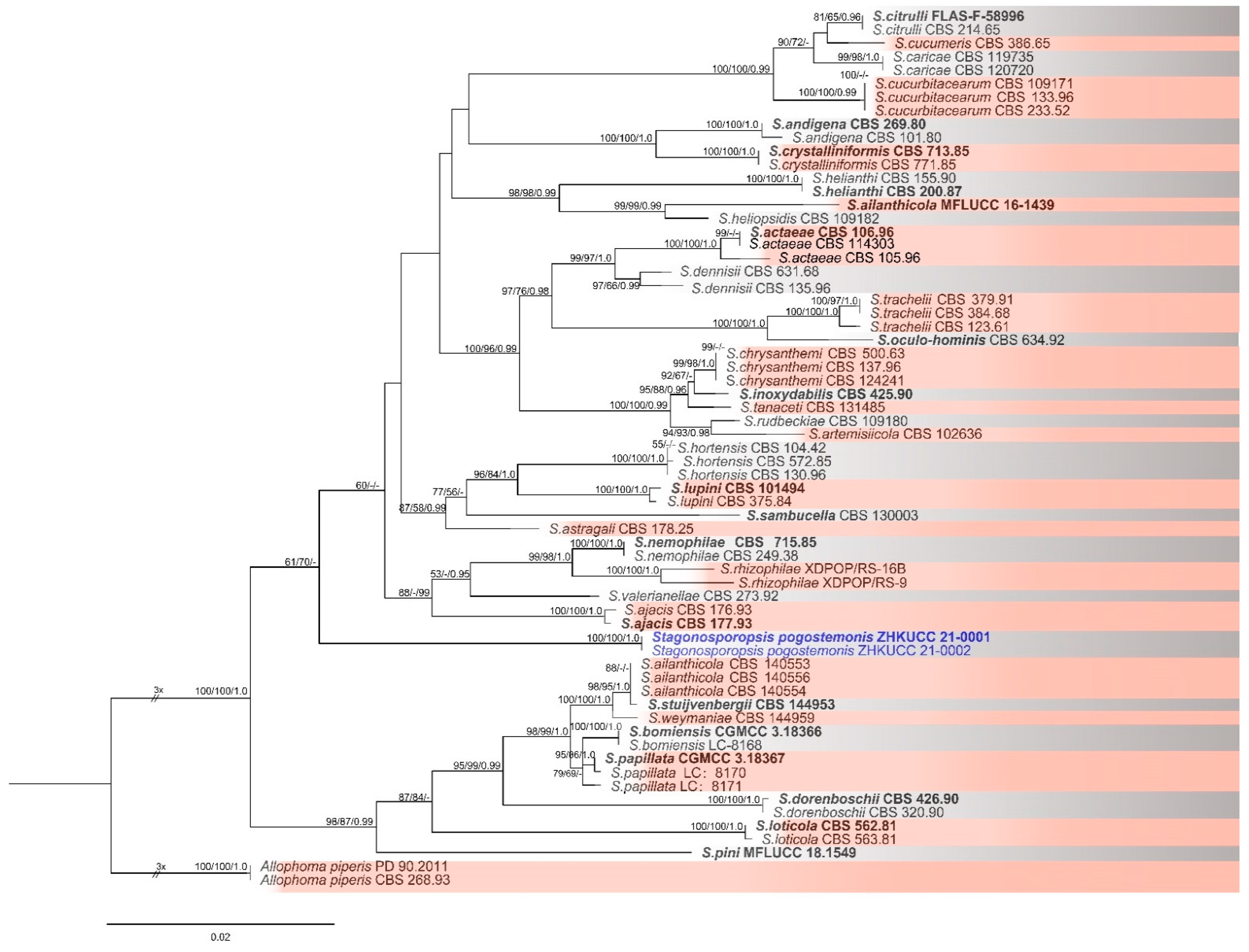
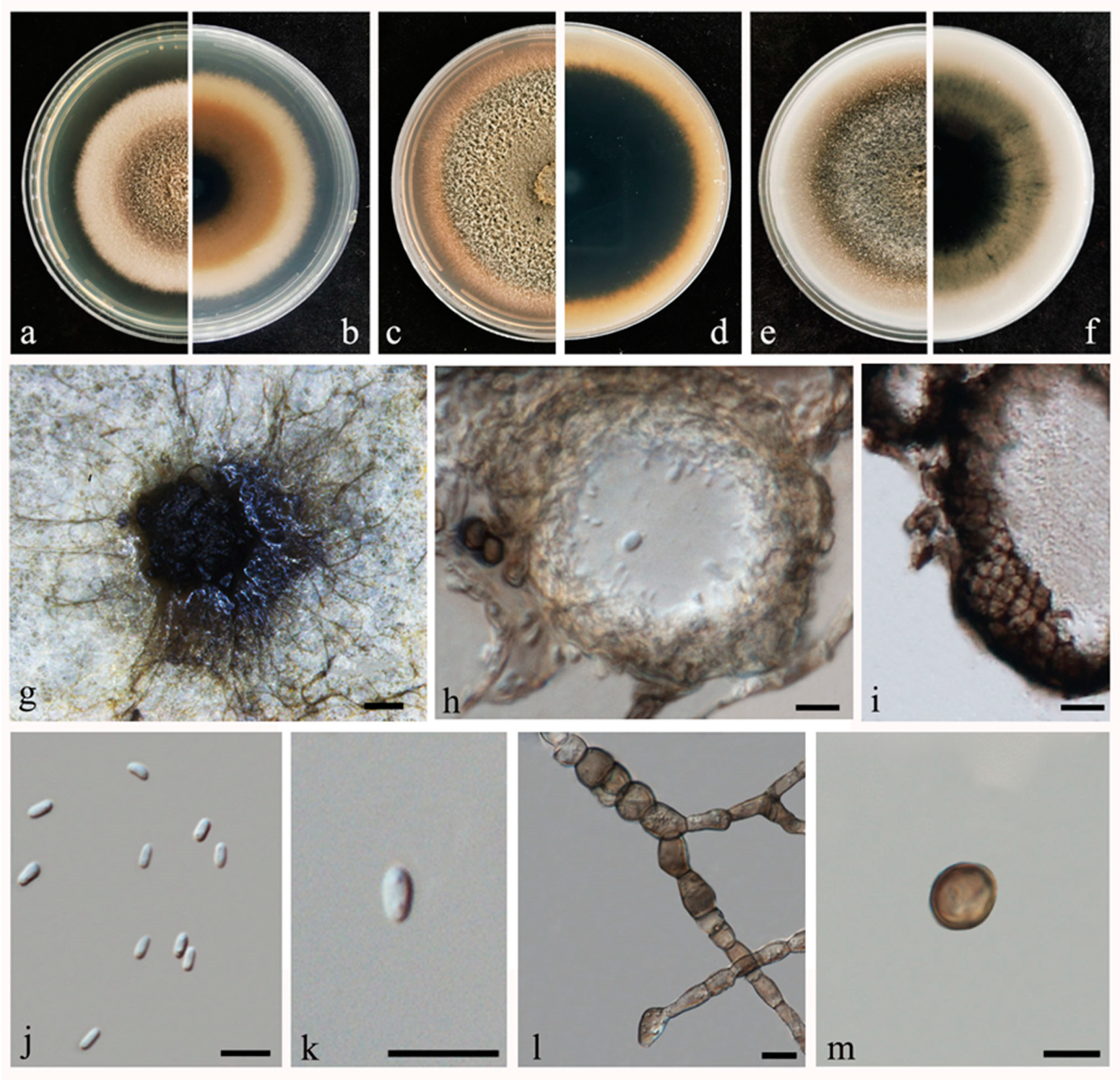
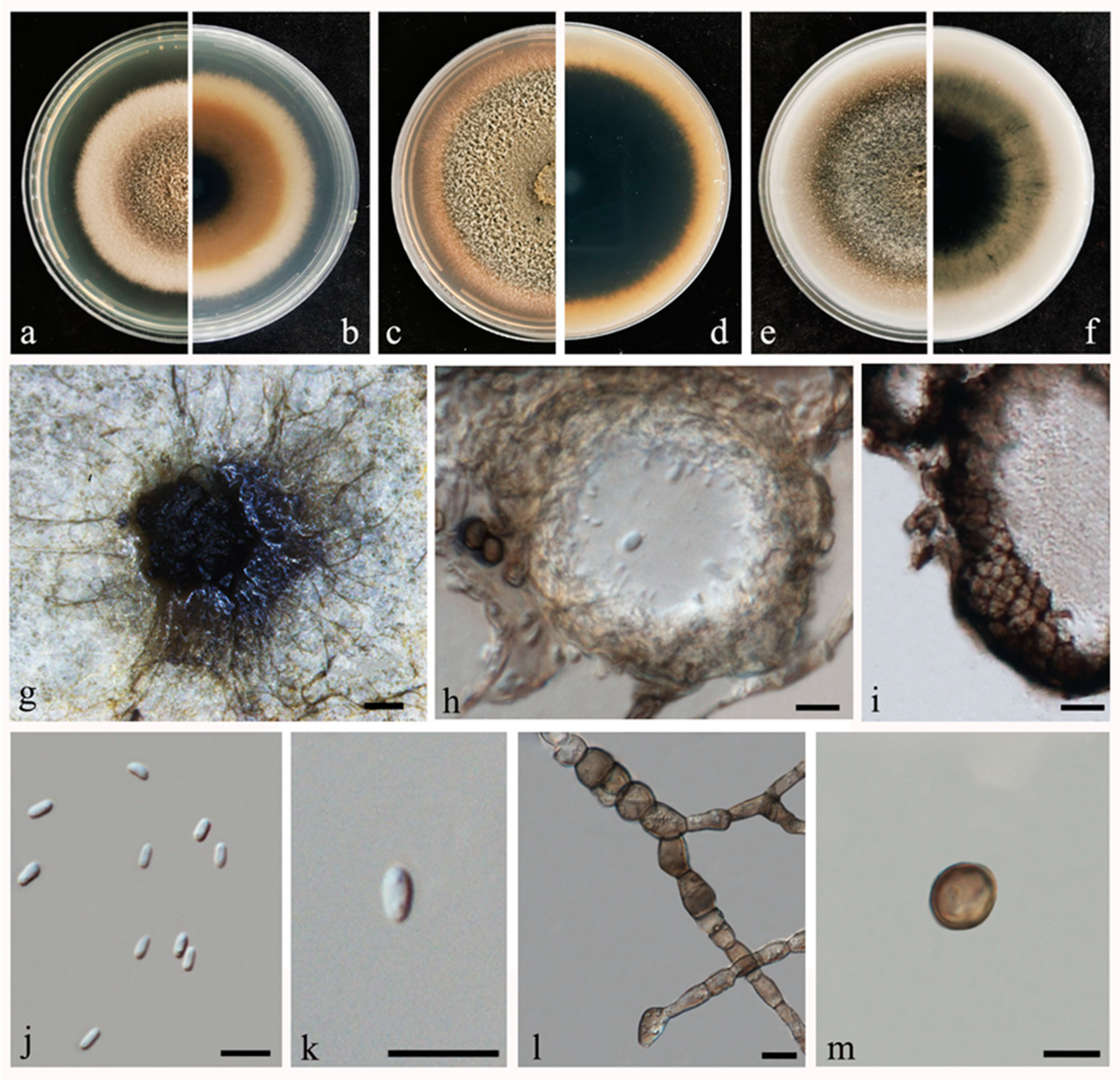
2.2. Morphological and Molecular Characterization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Strain Number 1 | Substrate (Including Host) | GenBank Accession Numbers 2 | |||
|---|---|---|---|---|---|---|
| rpb2 | tub2 | LSU | ITS | |||
| Stagonosporopsis Actaeae * | CBS 106.96; PD 94/1318 | Actaea spicata | KT389672 | GU237671 | GU238166 | GU237734 |
| S. actaeae | CBS 114303; UPSC 2962 | Actaea spicata | — | KT389847 | KT389760 | KT389544 |
| S. actaeae | CBS 105.96; PD 74/230 | Cimicifuga simplex | MT018018 | GU237670 | GU238165 | GU237733 |
| S. ailanthicola * | MFLUCC 16-1439 | Ailanthus altissima | KY100876 | KY100878 | KY100874 | KY100872 |
| CBS 140554 | House dust | MT018036 | MT005561 | MN943664 | MN973462 | |
| CBS 140553 | House dust | MT018037 | MT005562 | MN943665 | MN973463 | |
| CBS 140556 | House dust | MT018038 | MT005563 | MN943666 | MN973464 | |
| S. ajacis * | CBS 177.93; PD 90/115 | Delphinium sp. | KT389673 | GU237673 | GU238168 | GU237791 |
| S. ajacis | CBS 176.93; PD 86/547 | Delphinium sp. | MT018035 | GU237672 | GU238167 | GU237790 |
| S. andigena * | CBS 269.80; PD 75/914 | Solanum sp. | MT018026 | GU237675 | GU238170 | GU237817 |
| S. andigena | CBS 101.80; IMI 386090; PD 75/909 | Solanum sp. | — | GU237674 | GU238169 | GU237714 |
| S. artemisiicola | CBS 102636; PD 73/1409 | Artemisia dracunculus | KT389674 | GU237676 | GU238171 | GU237728 |
| S. astragali | CBS 178.25; MUCL 9915 | Astragalus sp. | MT018030 | GU237677 | GU238172 | GU237792 |
| S. bomiensis * | CGMCC 3.18366; LC 8167 | Boraginaceae | KY742189 | KY742365 | KY742277 | KY742123 |
| S. bomiensis | LC 8168 | Boraginaceae | KY742190 | KY742366 | KY742278 | KY742124 |
| S. caricae | CBS 119735 | Caricae papaya | MN983680 | MN984054 | MN973431 | MN973042 |
| S. caricae | CBS 120720 | Sechium edule | MN983681 | MN984055 | MN973432 | MN973043 |
| S. chrysanthemi | CBS 137.96; PD 84/75 | Chrysanthemum indicum | MT018011 | GU237696 | GU238191 | GU237783 |
| S. chrysanthemi | CBS 124241; PD 89/1016-4 | Chrysanthemum sinense | MT018010 | MT005550 | MN943653 | MN973451 |
| S. chrysanthemi | CBS 500.63; MUCL 8090 | Chrysanthemum indicum | MT018012 | GU237695 | GU238190 | GU237871 |
| S. citrulli * | FLAS-F-58996; C5-5 | Citrullus lanatus | — | KJ855602 | — | KJ855546 |
| S. cucurbitacearum | CBS 214.65; BBA 9963 | Cucumis sativus | MT018020 | MT005553 | MN943656 | MN973454 |
| S. crystalliniformis | CBS 771.85; IMI 386091; PD 85/772 | Solanum tuberosum | — | GU237684 | GU238179 | GU237906 |
| S. crystalliniformis * | CBS 713.85; ATCC 76027; PD 83/826 | Lycopersicon esculentum | KT389675 | GU237683 | GU238178 | GU237903 |
| S. cucumeris | CBS 386.65 | Cucumis sativus | MT018021 | MT005554 | MN943657 | MN973455 |
| S. cucurbitacearum | CBS 233.52 | — | MT018024 | MT005555 | MN943658 | MN973456 |
| S. cucurbitacearum | CBS 133.96; PD 79/127 | Cucurbita sp. | KT389676 | GU237686 | GU238181 | GU237780 |
| S. cucurbitacearum | CBS 109171; PD 91/310 | Cucurbita sp. | MN983682 | GU237685 | GU238180 | GU237922 |
| S. dennisii * | CBS 631.68; PD 68/147 | Solidago floribunda | KT389677 | GU237687 | GU238182 | GU237899 |
| S. dennisii | CBS 135.96; PD 95/4756 | Solidago canadensis | MT018019 | GU237688 | GU238183 | GU237782 |
| S. dorenboschii * | CBS 426.90; IMI 386093; PD 86/551 | Physostegia virginiana | KT389678 | GU237690 | GU238185 | GU237862 |
| S. dorenboschii | CBS 320.90; PD 86/932 | Physostegia virginiana | MT018039 | GU237689 | GU238184 | GU237830 |
| S. helianthi | CBS 155.90 | Helianthus annuus | MT018025 | MT005556 | MN943659 | MN973457 |
| S. helianthi * | CBS 200.87 | Helianthus annuus | KT389683 | KT389848 | KT389761 | KT389545 |
| S. heliopsidis | CBS 109182; PD 74/231 | Heliopsis patula | KT389679 | GU237691 | GU238186 | GU237747 |
| S. hortensis | CBS 572.85; PD 79/269 | Phaseolus vulgaris | KT389681 | GU237704 | GU238199 | GU237893 |
| S. hortensis | CBS 104.42 | — | KT389680 | GU237703 | GU238198 | GU237730 |
| S. hortensis | CBS 130.96 | Phaseolus vulgaris | MT018027 | MT005557 | MN943660 | MN973458 |
| S. inoxydabilis * | CBS 425.90; PD 81/520 | Chrysanthemum parthenii | KT389682 | GU237693 | GU238188 | GU237861 |
| S. loticola * | CBS 562.81; PDDCC 6884 | Lotus pedunculatus | KT389684 | GU237697 | GU238192 | GU237890 |
| S. loticola * | CBS 563.81; PDDCC 6799 | Lotus pedunculatus | MT018040 | MT005564 | MN943667 | MN973465 |
| S. lupini | CBS 375.84; PD 80/1250 | Lupinus mutabilis | MT018028 | GU237700 | GU238195 | GU237844 |
| S. lupini * | CBS 101494; PD 98/5247 | Lupinus albus | KT389685 | GU237699 | GU238194 | GU237724 |
| S. nemophilae | CBS 249.38 | Nemophila insignis | MT018032 | MT005560 | MN943663 | MN973461 |
| S. nemophilae * | CBS 715.85; PD 74/364 | Nemophila insignis | MT018031 | MT005559 | MN943662 | MN973460 |
| S. oculo-hominis * | CBS 634.92; IMI 193307 | Corneal ulcer | KT389686 | GU237701 | GU238196 | GU237901 |
| S. papillata | LC 8170 | Rumex nepalensis | KY742192 | KY742368 | KY742280 | KY742126 |
| S. papillata * | CGMCC 3.18367; LC 8169 | Rumex nepalensis | KY742191 | KY742367 | KY742279 | KY742125 |
| S. papillata | LC 8171 | Boraginaceae | KY742193 | KY742369 | KY742281 | KY742127 |
| S. pini * | MFLUCC 18-1549 | Pinus sp. | MK434860 | MK412886 | MK348019 | MK347800 |
| S. pogostemonis * | ZHKUCC 21-0001 | Pogostemon cablin | MZ203135 | MZ203132 | MZ191532 | MZ156571 |
| ZHKUCC 21-0002 | Pogostemon cablin | MZ203136 | MZ203133 | MZ191533 | MZ156572 | |
| S. rhizophilae | XDPOP-RS-9 | Populus deltoides | MN422105 | MN422099 | MN422103 | MN422101 |
| S. rhizophilae | XDPOP-RS-16B | Populus deltoides | MN422104 | MN422098 | MN422102 | MN422100 |
| S. rudbeckiae | CBS 109180; PD 79/175 | Rudbeckia bicolor | MT018015 | GU237702 | GU238197 | GU237745 |
| S. sambucella * | CBS 130003 | Sambucus nigra | MT018029 | MT005558 | MN943661 | MN973459 |
| S. stuijvenbergii * | CBS 144953; JW 132011 | Garden soil | MN824475 | MN824623 | MN823300 | MN823449 |
| S. tanaceti | CBS 131485 | Tanacetum cinerariifolium | MT018014 | MT005551 | MN943654 | MN973452 |
| S. trachelii | CBS 379.91; PD 77/675 | Campanula isophylla | KT389687 | GU237678 | GU238173 | GU237850 |
| S. trachelii | CBS 384.68 | Campanula isophylla | MT018016 | GU237679 | GU238174 | GU237856 |
| S. trachelii | CBS 123.61 | Campanula isophylla | MT018017 | MT005552 | MN943655 | MN973453 |
| S. valerianellae | CBS 273.92; PD 82/43 | Valerianella locusta | MT018033 | GU237705 | GU238200 | GU237819 |
| S. weymaniae | CBS 144959; JW 201003 | Garden soil | MN824479 | MN824627 | MN823304 | MN823453 |
| Allophoma piperis | CBS 268.93; PD 88/720 | Peperomia pereskifolia | KT389554 | GU237644 | GU238129 | GU237816 |
| Al. piperis | PD 90/2011 | Peperomia sp. | MT018045 | GU237645 | GU238130 | GU237921 |
2.3. Disease Symptoms and Pathogenicity Tests
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Fungal Isolation and Purification
4.3. DNA Extraction, PCR Amplification and Sequencings
4.4. Phylogenetic Analysis
4.5. Morphological Identification
4.6. Pathogenicity Test
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miyazawa, M.; Okuno, Y.; Nakamura, S.; Kosaka, H. Antimutagenic activity of flavonoids from Pogostemon cablin. J. Agric. Food Chem. 2000, 48, 642–647. [Google Scholar] [CrossRef]
- Bizzo, H.R.; Hovell, A.; Rezende, C.M. Leos essenciais no Brasil: Aspectos gerais, desenvolvimento e perspectivas. Química Nova 2009, 32, 588–594. [Google Scholar] [CrossRef]
- Wu, Y.G.; Guo, Q.S.; Zheng, H.Q. Textual research on history of introduction and herbal medicine of Pogostemon cablin. Chin. J. Chin. Mater. Med. 2007, 32, 2114–2117. [Google Scholar]
- Xu, S.J.; Wang, X.F.; Xu, X.H.; Xu, H.H.; Li, W.; Xu, L.; Deng, F.T.; Zhao, T. The classification of cultivars of Pogostemon cablin cultivated in Guangdong province of China. J. South Chin. North Univ. Nat. Sci. 2003, 1, 82–86. [Google Scholar]
- Mallappa, K.S.; Uma, R.S. Patchouli (Pogostemon cablin Benth): Botany, agrotechnology and biotechnological aspects. Ind. Crops Prod. 2016, 87, 161–176. [Google Scholar]
- Wan, F.; Fu, P.; Liang, X.; Chen, J.P.; Cheng, P.; Min, D. In vitro and In vivo antibacterial activity of Patchouli alcohol from Pogostemon cablin. Chin. J. Integr. Med. 2021, 27, 125–130. [Google Scholar] [CrossRef]
- Wu, X.; Yuan, X.L.; Zhai, F.F.; Xi, P.G.; Jiang, Z.D. First report of root rot of Pogostemon cablin caused by Phytophthora palmivora in China. Plant Dis. 2016, 100, 1249. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, G.; Li, Q.; He, L.; Zhang, Y.; Wang, Y.; He, H. Identification and characterization of virulence-attenuated mutants in Ralstonia solanacearum as potential biocontrol agents against bacterial wilt of Pogostemon cablin. Microb. Pathog. 2020, 147, 104418. [Google Scholar] [CrossRef]
- Thuy, T.T.T.; Yen, N.T.; Tuyet, N.T.A.; Te, L.L.; Waele, D.D. Population dynamics of Meloidogyne incognita on black pepper plants in two agro-ecological regions in Vietnam. Arch. Phytopathol. Plant Prot. 2012, 45, 1527–1537. [Google Scholar] [CrossRef]
- Borah, B.; Ahmed, R.; Hussain, M.; Phukon, P.; Wann, S.B.; Sarmah, D.K.; Singh, B.B. Suppression of root-knot disease in Pogostemon cablin caused by Meloidogyne incognita in a rhizobacteria mediated activation of phenylpropanoid pathway. Biol. Control 2018, 119, 43–50. [Google Scholar] [CrossRef]
- Singh, M.K.; Chandel, V.; Hallan, V.; Ram, R.; Zaidi, A.A. Occurrence of Peanut stripe virus on patchouli and raising of virus-free patchouli plants by meristem tip culture. J. Plant Dis. Prot. 2009, 116, 2–6. [Google Scholar] [CrossRef]
- Chen, X.Y.; Sui, C.; Gan, B.C.; Wei, J.H.; Zhou, Y.K. First report of Corynespora leaf spot on patchouli caused by Corynespora cassiicola in China. Plant Dis. 2010, 94, 1508. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al–Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Aveskamp, M.M.; Gruyter, J.D.; Woudenberg, J.H.C.; Verkley, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef]
- Vaghefi, N.; Hay, F.S.; Ades, P.K.; Pethybridge, S.J.; Ford, R.; Taylor, P.W.J. Rapid changes in the genetic composition of Stagonosporopsis tanaceti population in Australian Pyrethrum fields. Phytopathology 2015, 105, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qian, Y.; Zheng, X.; Zhou, Y.; Xiong, Q. Stagonosporopsis trachelii causes leaf spot on Ningpo Figwort (Scrophularia ningpoensis) in China. Australas. Plant Dis. Notes 2019, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Chester, F.D. Notes on three new or noteworthy diseases of plants. Bull. Torrey Bot. Club 1891, 18, 371–374. [Google Scholar] [CrossRef]
- Mahapatra, S.; Rao, E.S.; Sandeepkumar, G.M.; Sriram, S. Stagonosporopsis cucurbitacearum the causal agent of gummy stem blight of watermelon in India. Australas. Plant Dis. Notes 2020, 15, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Chiu, W.F.; Walker, J.C. Physiology and pathogenicity of the cucurbit black-rot fungus. J. Agric. Res. 1949, 78, 589–615. [Google Scholar]
- Basım, E.; Basım, H.; Abdulai, M.; Baki, D.; Nurhan, Z. Identification and characterization of Didymella bryoniae causing gummy stem blight disease of watermelon (Citrullus lanatus) in Turkey. Crop Prot. 2016, 90, 150–156. [Google Scholar] [CrossRef]
- Power, H.J. South Carolina vegetable statistics: 1991. South Carol. Agric. Exp. Stn. Bull. 1992, 472, 14. [Google Scholar]
- Vaghefi, N.; Pethybridge, S.J.; Ford, R.; Nicolas, M.E.; Crous, P.W.; Taylor, P.W.J. Stagonosporopsis spp. associated with ray blight disease of Asteraceae. Australas. Plant Pathol. 2012, 41, 675–686. [Google Scholar] [CrossRef]
- Walftor, D.; Park, M.J.; Park, J.H.; Yang, C.Y.; Back, C.G. First report of bacterial shot-hole disease caused by Xanthomonas arboricola pv. pruni on plumcot in South Korea. Plant Dis. 2020, 105, 697. [Google Scholar]
- Sun, L.F.; Zhang, Y.H.; Pei, K.Q. A rapid extraction of genomic DNA from fungi. Mycosystem 2009, 28, 299–302. [Google Scholar]
- Kim, W.; Chen, W. Phytotoxic, Metabolites produced by legume-associated Ascochyta and its related genera in the Dothideomycetes. Toxins 2019, 11, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohm, R.A.; Feau, N.; Henrissat, B.; Schoch, C.L.; Horwitz, B.A.; Barry, K.W.; Condon, B.J.; Copeland, A.C.; Dhillon, B.; Glaser, F.; et al. Diverse lifestyles and strategies of plant pathogenesis encoded in the genomes of eighteen Dothideomycetes fungi. PLoS Pathog. 2012, 8, e1003037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.W.; Groenewald, J.Z.; Pfenning, L.H.; Yarden, O.; Crous, P.W.; Cai, L. The Phoma-like dilemma. Stud. Mycol. 2020, 96, 309–396. [Google Scholar] [CrossRef]
- Bracale, M.F.; Nobrega, T.F.; Barreto, R.W. Fungal diseases of non-conventional food plants: First report of Stagonosporopsis caricae causing leaf spots on Vasconcellea monoica. Australas. Plant. Dis. Notes 2020, 15, 20. [Google Scholar] [CrossRef]
- Vaghefi, N.; Pethybridge, S.J.; Hay, F.S.; Ford, R.; Nicolas, M.E.; Taylor, P.W.J. Revisiting Stagonosporopsis species associated with Chrysanthemum and Pyrethrum ray blight. Australas. Plant Pathol. 2016, 45, 561–570. [Google Scholar] [CrossRef]
- Bhuiyan, M.A.H.B.; Vaghefi, N.; Taylor, P.W.J. Ray blight of Pyrethrum in Australia: A review of the current status and future opportunities. Plant Pathol. 2019, 68, 620–627. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsa-ard, J.; Thangavel, R.; et al. Fungal plant description sheets: 951–1041. Persoonia Mol. Phylogeny Evol. Fungi 2019, 43, 223–425. [Google Scholar] [CrossRef]
- He, Y.L.; He, G.; Li, Q.Q.; Lin, W.; Yuan, G.Q. First report of Stagonosporopsis vannaccii causing leaf spot on Crassocephalum crepidioides in China. Plant Dis. 2020, 105, 499. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, J.; Zhang, L.; Xu, L.; Yan, C.; Gong, Z. Identification and characteristics of Stagonosporopsis cucurbitacearum pathogenic factors influencing pumpkin seeding survival in northeast China. J. Phytopathol. 2019, 167, 104418. [Google Scholar] [CrossRef] [Green Version]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Rennberger, G.; Keinath, A.P. Susceptibility of fourteen new cucurbit species to gummy stem blight caused by Stagonosporopsis citrulli under field conditions. Plant Dis. 2018, 102, 1365–1375. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, M.A.H.B.; Groom, T.; Nicolas, M.E.; Taylor, P.W.J. TaqMan PCR assay for detection and quantification of Stagonosporopsis tanaceti in Pyrethrum seed and seedlings. Eur. J. Plant Pathol. 2018, 150, 1041–1048. [Google Scholar] [CrossRef]
- Van Laethem, S.; Frans, M.; Aerts, R.; Ceusters, J. pH modulation of the environment by Stagonosporopsis cucurbitacearum, an important pathogen causing fruit rot in Cucurbitaceae. Eur. J. Plant Pathol. 2021, 159, 235–245. [Google Scholar] [CrossRef]
- Liu, P.Q.; Wei, M.Y.; Zhu, L.; Li, B.J.; Weng, Q.Y.; Chen, Q.H. First report of spot blight on water spinach (Ipomoea aquatica) caused by Stagonosporopsis cucurbitacearum in China. Plant Dis. 2017, 101, 838–839. [Google Scholar] [CrossRef]
- Liang, Y.M.; Yang, Y.U.; Sun, Y.P.; Liu, J.S.; Zong, H.M.; Liu, H.T.; Wang, G.; Pharmacy, S. Secondary metabolites of endophytic fungus Stagonosporopsis oculihominis from Dendrobium huoshanense. Nat. Prod. Res. Dev. 2018, 30, 783–788. [Google Scholar]
- Manawasinghe, I.S.; Phillips, A.; Hyde, K.D.; Chethana, K.W.T.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X. Mycosphere Essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Dissanayake, A.J.; Li, X.H.; Liu, M.; Wanasinghe, D.N.; Xu, J.P.; Zhao, W.S.; Zhang, W.; Zhou, Y.; Hyde, K.D.; et al. High genetic diversity and species complexity of Diaporthe associated with grapevine dieback in China. Front. Microbiol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S.; et al. What is a species in fungal plant pathogens? Fungal Divers. 2021, 1–28. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.L.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Tępień, L.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in plant pathology: Beyond the species level. Fungal Divers. 2021, 1–16. [Google Scholar] [CrossRef]
- Stephen, A.R.; Gary, J.S. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; White, T.; Lee, S.H.; Taylor, L.; Shawetaylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols: A guide to methods and applications. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799. [Google Scholar] [CrossRef]
- Wei, H.; He, X.H.; Riccardo, B.; Yang, Y.; Yuan, Z. Stagonosporopsis rhizophilae sp. nov. (Didymellaceae, Pleosporales), a new rhizospheric soil fungus associated with Populus deltoides Marsh. Phytotaxa 2021, 491, 23–34. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Diver. Evolut. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4.0b10. Mac. Vers. 2002. [Google Scholar] [CrossRef]
- Ronquist, F.R.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinf. Oxf. 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in Hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest Version 2; Evolutionary Biology Centre Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 309, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Garampalli, R.H.; Gapalkrishna, M.K.; Li, H.X.; Brewer, M.T. Two Stagonosporopsis species identified as causal agents of gummy stem blight epidemics of gherkin cucumber (Cucumis sativus) in Karnataka, India. Eur. J. Plant Pathol. 2016, 145, 507–512. [Google Scholar] [CrossRef]



| Amplified Gene. | Primer Pairs | Optimized PCR Protocols | References |
|---|---|---|---|
| rpb2 | fRPB2-5F | 95 °C: 5 min, (94 °C: 30 s, 54 °C: 30 s, 72 °C: 1 min) × 32 cycles, 72 °C: 10 min | [49] |
| fRPB2-7cR | |||
| β-tubulin | Bt2a | 95 °C: 5 min, (94 °C: 30 s, 58 °C: 30 s, 72 °C: 1 min) × 32 cycles, 72 °C: 10 min | [48] |
| Bt2b | |||
| LSU | LROR | 95 °C: 5 min, (94 °C: 30 s, 49 °C: 30 s, 72 °C: 1 min) × 32 cycles, 72 °C: 10 min | [45,46] |
| LR5 | |||
| ITS | ITS1 | 95 °C: 5 min, (94 °C: 30 s, 53 °C: 30 s, 72 °C: 1 min) × 32 cycles, 72 °C: 10 min | [47] |
| ITS4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.-Y.; Huang, Y.-H.; Manawasinghe, I.S.; Wanasinghe, D.N.; Liu, J.-W.; Shu, Y.-X.; Zhao, M.-P.; Xiang, M.-M.; Luo, M. Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens 2021, 10, 1093. https://doi.org/10.3390/pathogens10091093
Dong Z-Y, Huang Y-H, Manawasinghe IS, Wanasinghe DN, Liu J-W, Shu Y-X, Zhao M-P, Xiang M-M, Luo M. Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens. 2021; 10(9):1093. https://doi.org/10.3390/pathogens10091093
Chicago/Turabian StyleDong, Zhang-Yong, Ying-Hua Huang, Ishara S. Manawasinghe, Dhanushka N. Wanasinghe, Jia-Wei Liu, Yong-Xin Shu, Min-Ping Zhao, Mei-Mei Xiang, and Mei Luo. 2021. "Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China" Pathogens 10, no. 9: 1093. https://doi.org/10.3390/pathogens10091093
APA StyleDong, Z.-Y., Huang, Y.-H., Manawasinghe, I. S., Wanasinghe, D. N., Liu, J.-W., Shu, Y.-X., Zhao, M.-P., Xiang, M.-M., & Luo, M. (2021). Stagonosporopsis pogostemonis: A Novel Ascomycete Fungus Causing Leaf Spot and Stem Blight on Pogostemon cablin (Lamiaceae) in South China. Pathogens, 10(9), 1093. https://doi.org/10.3390/pathogens10091093