
Streptococcus suis Isolates—Serotypes and Susceptibility to Antimicrobials in Terms of Their Use on Selected Repopulated Czech Pig Farms

 ,
,
Abstract
:1. Introduction
2. Results
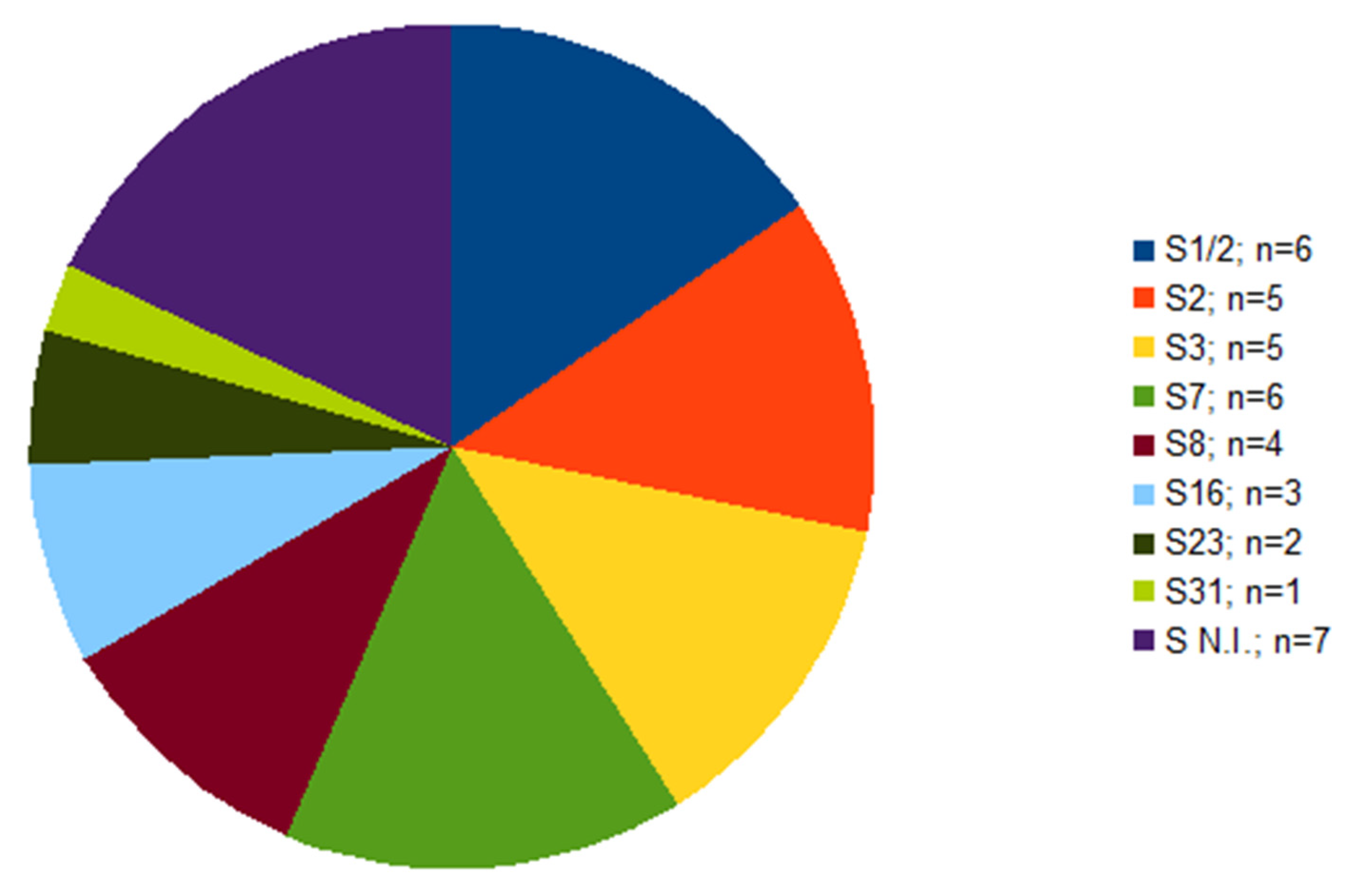
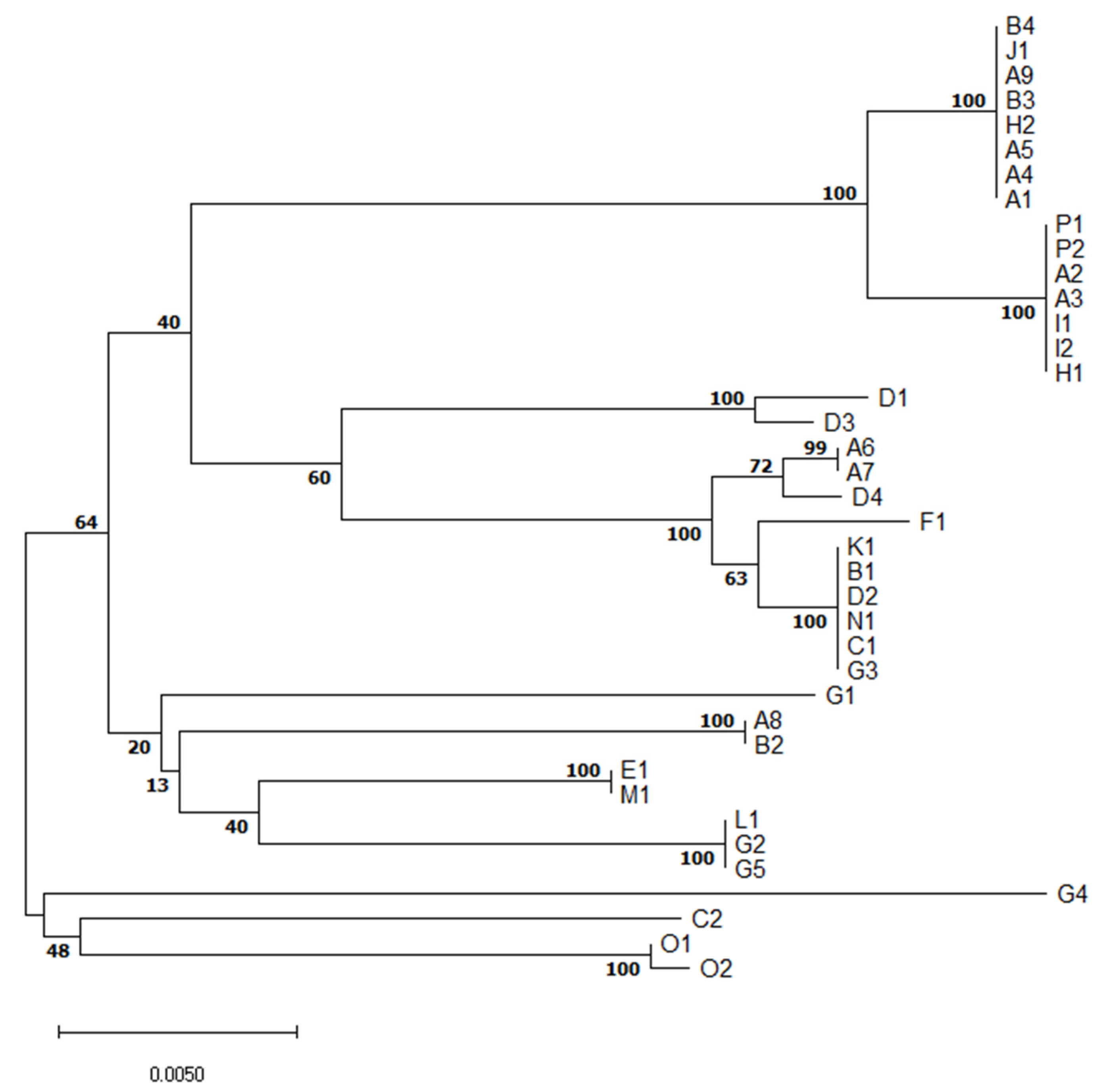
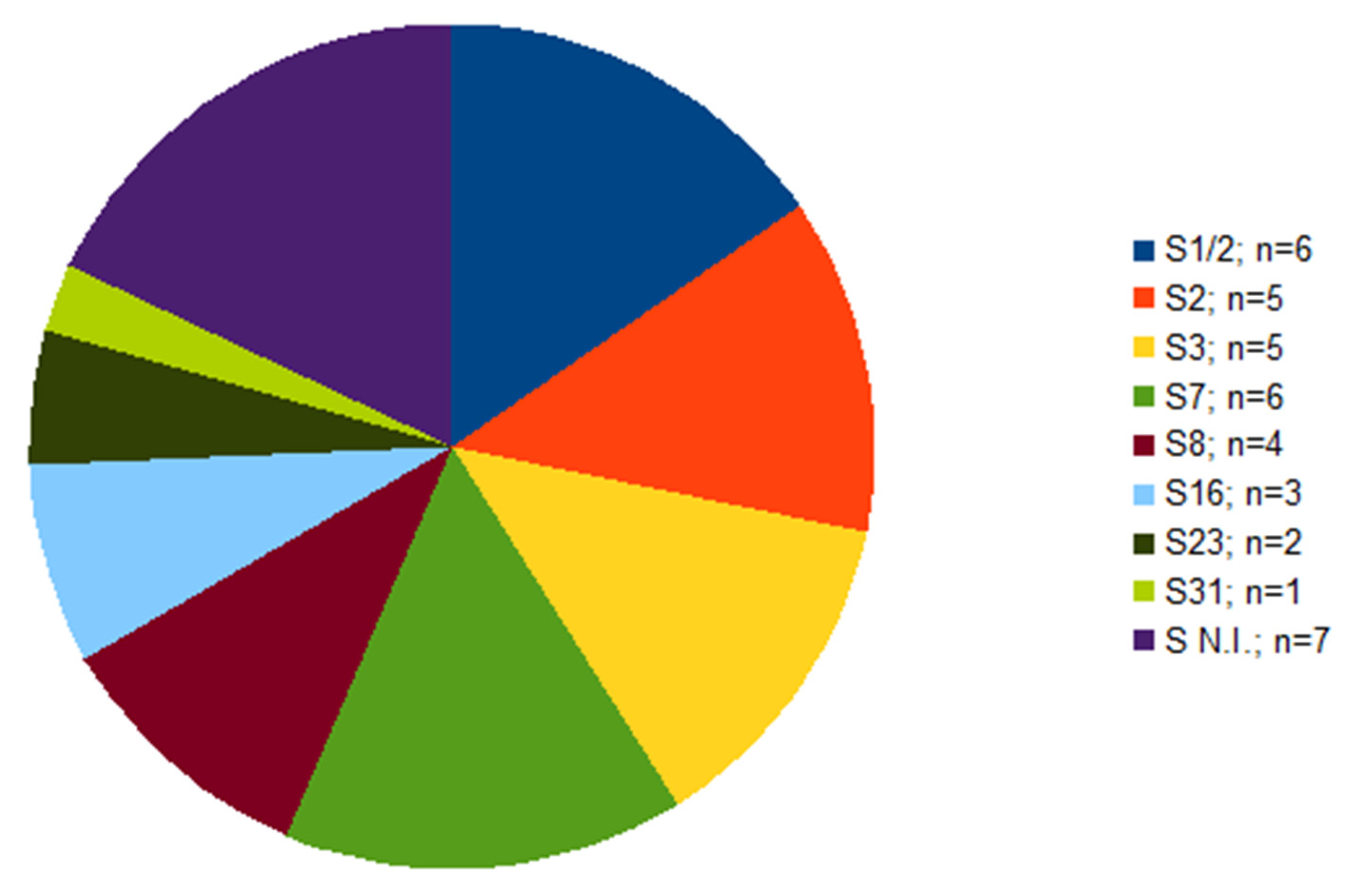
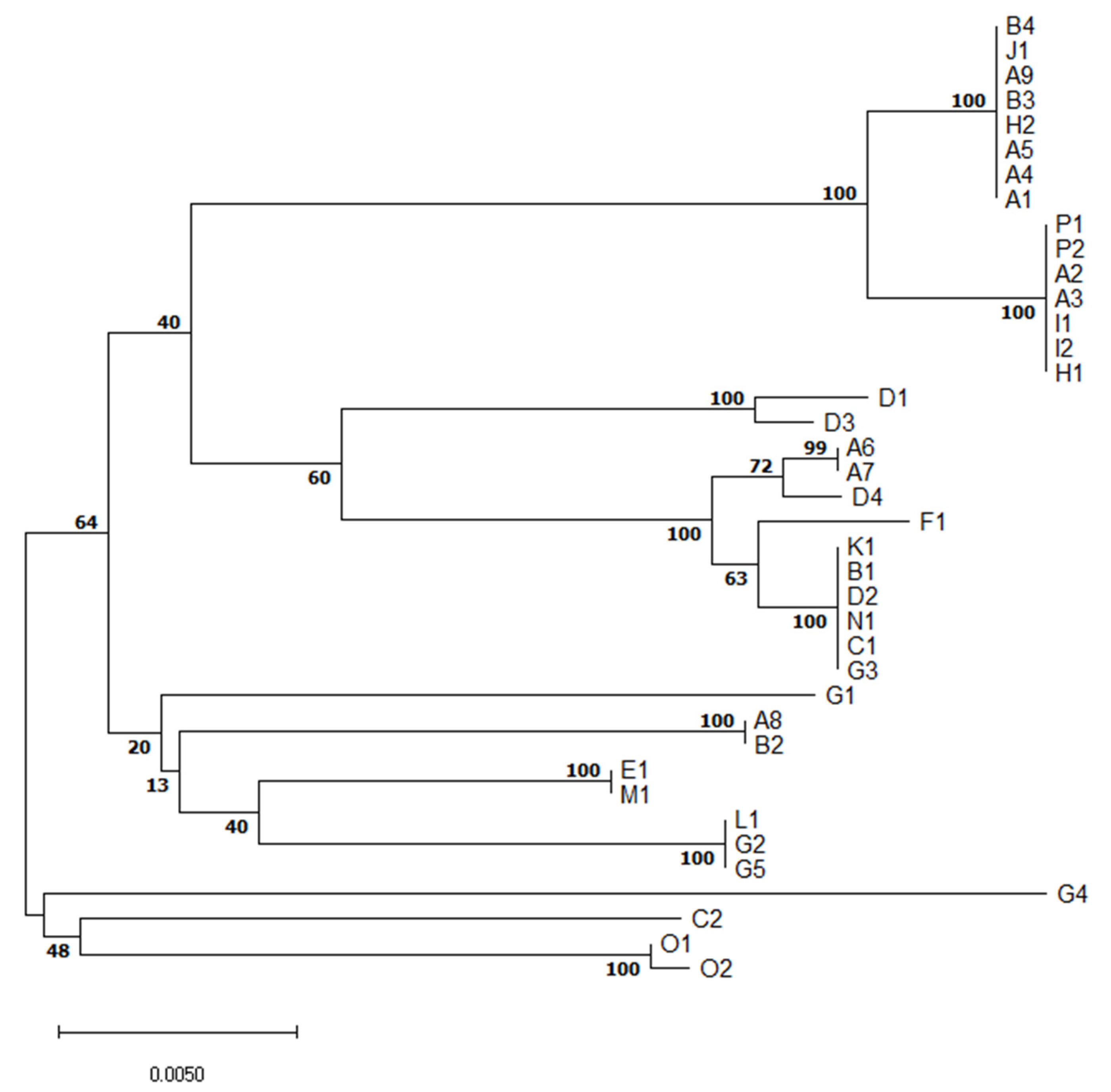
2.1. Serotyping and Multilocus Sequence Typing (MLST)
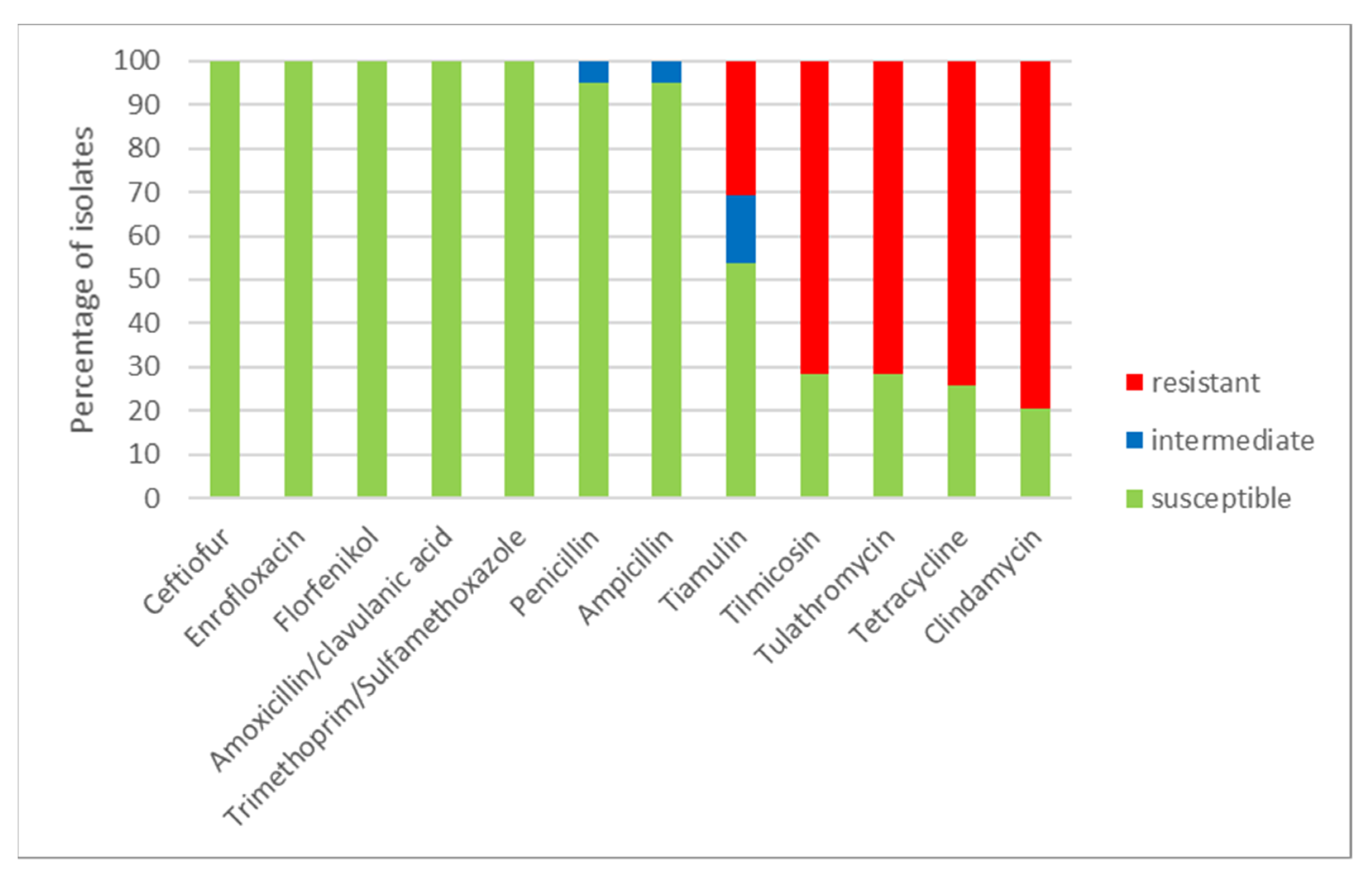
2.2. Antimicrobial Susceptibility Testing
2.3. Routine Antimicrobial Control Treatments and Resistance on Farms
3. Discussion
4. Materials and Methods
4.1. Farms
4.2. Routine Antimicrobial Control Treatments
4.3. Bacterial Sampling
4.4. Bacterial Isolation and Identification
4.5. PCR Serotyping, Species Verification and Multi Locus Sequence Typing
4.6. Evolutionary Relationships of Isolates
4.7. Antimicrobial Susceptibility Testing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Power, S.B. Streptococcus suis type 2 infection in pigs. Vet. Res. 1978, 102, 215–216. [Google Scholar] [CrossRef]
- Gottschalk, M. Streptococcus suis infection. Merck manuals 2014. Available online: www.merckmanuals.com (accessed on 21 August 2021).
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent—an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microb. Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- de Briyne, N.; Iatridou, D. Antimicrobial use in food-producing animals, FVE input to RONAFA report. EFSA J. 2016, 15. [Google Scholar] [CrossRef]
- Perch, B.; Pedersen, K.B.; Heinrichsen, J. Serology of capsulated streptococci pathogenic for pigs: Six new serotypes of Streptococcus suis. J. Clin. Microbiol. 1983, 17, 993–996. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Mittal, K.R.; Henrichsen, J. Description of 14 new capsular types of Streptococcus suis. J. Clin. Microbiol. 1989, 27, 2633–2635. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Beaudoin, M.; Henrichsen, J. Characterization of six new capsular types (23 through 28) of Streptococcus suis. J. Clin. Microbiol. 1991, 29, 2590–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, R.; Gottschalk, M.; Boudreau, M.; Lebrun, A.; Henrichsen, J. Description of six new capsular types (29-34) of Streptococcus suis. J. Vet. Diagn. Invest. 1995, 7, 405–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Tien, H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA-DNA homology and sodA and recN phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef]
- Okura, M.; Lachance, C.; Osaki, M.; Sekizaki, T.; Maruyama, F.; Nozawa, T.; Nakagawa, I.; Hamada, S.; Rossignol, C.; Gottschalk, M.; et al. Development of a two-step multiplex PCR assay for typing of capsular polysaccharide synthesis gene clusters of Streptococcus suis. J. Clin. Microbiol. 2014, 52, 1714–1719. [Google Scholar] [CrossRef] [Green Version]
- Werinder, A.; Aspan, A.; Backhans, A.; Sjolund, M.; Guss, B.; Jacobson, M. Streptococcus suis in Swedish grower pigs: Occurence, serotypes, and antimicrobial susceptibility. A. Vet. Scand. 2020, 62, 36. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, H.; Gottschalk, M.; Bai, X.; Lan, R.; Ji, S.; Liu, H.; Xu, J. Development of multiplex PCR assays for the identification of the 33 serotypes of Streptococcus suis. PLoS ONE 2013, 8, e72070. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Higgins, R.; Boudreau, M. Use of polyvalent coagglutination reagents for serotyping of Streptococcus suis. J. Clin. Microbiol. 1993, 31, 2192–2194. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Segura, M.; Xu, J. Streptococcus suis infections in humans: The Chinese experience and the situation in North America. Anim. Health Res. Rev. 2007, 8, 29–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athey, T.B.; Teatero, S.; Lacouture, S.; Takamatsu, D.; Gottschalk, M.; Fittipaldi, N. Determining Streptococcus suis serotype from short-read whole-genome sequencing data. BMC Microbiol. 2016, 16, 162. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; Athey, T.B.T.; Auger, J.P.; Goyette-Desjardins, G.; Van Calsteren, M.R.; Takamatsu, D.; Okura, M.; Teatero, S.; Alcorlo, M.; Hermoso, J.A.; et al. A single amino acid polymorphism in the glycosyltransferase CpsK defines four Streptococcus suis serotypes. Sci. Rep. 2017, 7, 4066. [Google Scholar] [CrossRef]
- Scherrer, S.; Rademacher, F.; Spoerry Serrano, N.; Schrenzel, J.; Gottschalk, M.; Stephan, R.; Landolt, P. Rapid high resolution melting assay to differentiate Streptococcus suis serotypes 2, 1/2, 1, and 14. Microbiol. open 2020, 9, e995. [Google Scholar] [CrossRef] [PubMed]
- Matiasovic, J.; Zouharova, M.; Nedbalcova, K.; Kralova, N.; Matiaskova, K.; Simek, B.; Kucharovicova, I.; Gottschalk, M. Resolution of Streptococcus suis serotypes 1/2 versus 2 and 1 versus 14 by PCR-Restriction Fragment Lenght Polymorphism Method. J. Clin. Microbiol. 2020, 58, e00480-20. [Google Scholar] [CrossRef]
- Lopez, A.; Martinson, S. Streptococcal pneumonia. In Pathologic Basis of Veterinary Disease, 6th ed.; Zachary, J.F., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. v–vi. [Google Scholar] [CrossRef]
- Segura, M.; Aragon, V.; Brockmeier, S.L.; Gebhart, C.; de Greeff, A.; Kerdsin, A.; O’Dea, M.A.; Okura, M.; Saléry, M.; Schultsz, C.; et al. Update on Streptococcus suis Research and Prevention in the Era of Antimicrobial Restriction: 4th International Workshop on S. suis. Pathogens 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed]
- Varela, N.P.; Gadbois, P.; Tibault, C.; Gottschalk, M.; Dick, P.; Wilson, J. Antimicrobial resistance and prudent drug use for Streptococcus suis. Anim. Health Res. Rev. 2013, 14, 68–77. [Google Scholar] [CrossRef]
- SVS ČR (State Veterinary Administration CZ), SVÚ (State Veterinary Institute) Jihlava (2019). National Programme of Monitoring Antimicrobial Resistance of Pathogens with Veterinary Importance for 2017-Part I (Národní Program Sledování Rezistencí k Antimikrobikům u Veterinárně Významných patogenů za rok 2017 -část I) in Czech. 24 p. Available online: https://www.svscr.cz/wp-content/files/dokumenty-a-publikace/Narodni-program-sledovani-rezistenci-k-antimikrobikum-u-veterinarne-vyznamnych-patogenu-2017-cast-I.pdf (accessed on 1 October 2021).
- SVS ČR (State Veterinary Administration CZ), SVÚ (State Veterinary Institute) Jihlava (2019). National Programme of Monitoring Antimicrobial Resistance of Pathogens with Veterinary Importance for 2018 (Národní Program Sledování Rezistencí k Antimikrobikům u Veterinárně Významných Patogenů za rok 2018) in Czech. 36 p. Available online: https://www.svscr.cz/wp-content/files/dokumenty-a-publikace/Zprava-RL-antibioticke-centrum-pro-veterinarni-klinickou-praxi-za-rok-2018.pdf (accessed on 21 August 2021).
- SVS ČR (State Veterinary Administration CZ), SVÚ (State Veterinary Institute) Jihlava (2020). National Programme of Monitoring Antimicrobial Resistance of Pathogens with Veterinary Importance for 2019 (Národní Program Sledování Rezistencí k Antimikrobikům u Veterinárně Významných Patogenů za rok 2019) in Czech. 36 p. Available online: https://www.svscr.cz/wp-content/files/dokumenty-a-publikace/Zprava-RL-2019.pdf (accessed on 21 August 2021).
- SVÚ (State Veterinary Institute) Jihlava, SVS ČR (State Veterinary Administration CZ) (2021). National Programme of Monitoring Antimicrobial Resistance of Pathogens with Veterinary Importance for 2020 (Národní Program Sledování Rezistencí k Antimikrobikům u Veterinárně Významných Patogenů za rok 2020) in Czech. 36 p. Available online: https://www.svscr.cz/wp-content/files/dokumenty-a-publikace/Narodni-program-sledovani-rezistenci-k-antimikrobikum-u-veterinarne-vyznamnych-patogenu-2020.pdf (accessed on 21 August 2021).
- Helke, K.L.; Ezell, P.C.; Duran-Struuck, R.; Swindle, M.M. Chapter 16—Biology and Diseases of Swine. In Laboratory Animal Medicine, 3rd ed.; Fox, J.G., Anderson, L.C., Otto, G.M., Pritchett-Corning, K.R., Whary, M.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 695–769. [Google Scholar]
- Okura, M.; Osaki, M.; Nomoto, R.; Arai, S.; Osawa, R.; Sekizaki, T.; Takamatsu, D. Current taxonomical situation of Streptococcus suis. Pathogens 2016, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Mai, N.T.H.; Hoa, N.T.; Nga, T.V.T.; Linh, L.D.; Chau, T.T.H.; Sinh, D.X.; Phu, N.H.; Chuong, L.V.; Diep, T.S.; Campbell, J.; et al. Streptococcus suis meningitis in adults in Vietnam. Clin. Infect. Dis. 2008, 46, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Schultsz, C.; Jansen, E.; Keijzers, W.; Rothkamp, A.; Duim, B.; Wagenaar, J.A.; van der Ende, A. Differences in the population structure of invasive Streptococcus suis strains isolated from pigs and from humans in The Netherlands. PLoS ONE 2012, 7, e33854. [Google Scholar] [CrossRef] [Green Version]
- Fittipaldi, N.; Xu, J.; Lacouture, S.; Tharavichitkul, P.; Osaki, M.; Sekizaki, T.; Takamatsu, D.; Gottschalk, M. Lineage and virulence of Streptococcus suis serotype 2 isolates from North America. Emerg. Infect. Dis. 2011, 17, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Vötsch, D.; Willenborg, M.; Weldearegay, Y.B.; Valentin-Weigand, P. Streptococcus suis—The “Two Faces” of a Pathobiont in the Porcine Respiratory Tract. Front. Microbiol. 2018, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Dong, W.; Ma, J.; Zhang, Y.; Zhong, X.; Pan, Z.; Liu, G.; Wu, Z.; Yao, H. Comparative genetic analyses provide clues about capsule switching in Streptococcus suis 2 strains with different virulence levels and genetic backgrounds. Microbiol. Res. 2021, 250, 126814. [Google Scholar] [CrossRef] [PubMed]
- ÚSKVBL (Institute for State Control of Veterinary Biologicals and Medicines). 2020 Consumption of Antimicrobials in Veterinary Medicine in CZ: Comparison of Trends in Consumption of Antimicrobials 2010-2018 Commented in Details (Spotřeba Antimikrobik ve Veterinární Medicíně v ČR: Detailní Komentované Srovnání Trendů ve Spotřebách Antimikrobik 2010-2018) in Czech. 48 p. Available online: http://www.uskvbl.cz/cs/informace/tiskove-centrum/tiskprohl (accessed on 31 August 2021).
- Hernandez-Garcia, J.; Wang, J.; Restif, O.; Holmes, M.A.; Mather, A.E.; Weinert, L.A.; Wileman, T.M.; Thomson, J.R.; Langford, P.R.; Wren, B.W.; et al. Patterns of antimicrobial resistance in Streptococcus suis isolates from pigs with or without streptococcal disease in England between 2009 and 2014. Vet. Microbiol. 2017, 207, 117–124. [Google Scholar] [CrossRef]
- Vela, A.I.; Moreno, M.A.; Cebolla, J.A.; González, S.; Latre, M.V.; Domínguez, L.; Fernández Garayzábala, J.F. Antimicrobial susceptibility of clinical strains of Streptococcus suis isolated from pigs in Spain. Vet. Microbiol. 2004, 105, 143–147. [Google Scholar] [CrossRef]
- Soares, T.C.; Paes, A.C.; Megid, J.; Ribolla, P.E.; Paduan, K.; Gottschalk, M. Antimicrobial susceptibility of Streptococcus suis isolated from clinically healthy swine in Brazil. Canadian, J. Vet. Res. 2014, 78, 145–149. [Google Scholar]
- Yongkiettrakul, S.; Maneerat, K.; Arechanajan, B.; Malila, Y.; Srimanote, P.; Gottschalk, M.; Visessanguan, W. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Vet. Res. 2019, 15, 5. [Google Scholar] [CrossRef] [Green Version]
- EMA 2019: Categorisation of Antibiotics for Use in Animals for Prudent and Responsible Use. Available online: https://www.ema.europa.eu/en/documents/report/infographic-categorisation-antibiotics-use-animals-prudent-responsible-use_en.pdf.
- Kerdsin, A.; Akeda, Y.; Hatrongjit, R.; Detchawna, U.; Sekizaki, T.; Hamada, S.; Gottschalk, M.; Oishi, K. Streptococcus suis serotyping by a new multiplex PCR. J. Med. Microbiol. 2014, 63, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Van Hout, J.; Heuvelink, A.; Gonggrijp, M. Monitoring of antimicrobial susceptibility of Streptococcus suis in the Netherlands, 2013–2015. Vet. Microbiol. 2016, 194, 5–10. [Google Scholar] [CrossRef]
- Ishida, S.; Tien le, H.T.; Osawa, R.; Tohya, M.; Nomoto, R.; Kawamura, Y.; Takahashi, T.; Kikuchi, N.; Kikuchi, K.; Sekizaki, T. Development of an appropriate PCR system for the reclassification of Streptococcus suis. J. Microbiol. Methods 2014, 107, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Mittal, K.R.; Higgins, R.; Larivière, S. Identification and serotyping of Haemophilus pleuropneumoniae by coagglutination test. J. Clin. Microbiol 1983, 18, 1351–1354. [Google Scholar] [CrossRef] [Green Version]
- King, S.J.; Leigh, J.A.; Heath, P.J.; Luque, I.; Tarradas, C.; Dowson, C.G.; Whatmore, A.M. Development of a multilocus sequence typing scheme for the pig pathogen Streptococcus suis: Identification of virulent clones and potential capsular serotype exchange. J. Clin. Microbiol. 2002, 40, 3671–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- CLSI (Clinical and Laboratory Standards Institute) (2018). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI supplement VET08; Clinical and Laboratory Standards Institute: Wayne, PA, USA; p. 170.


{kind=link}
{kind=link}
{kind=link}
| Farm1 | Date of Sampling | Animal Category | Organ of Isolation | Serotype | MLST | MIC (mg/L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PNC | AMP | AMC * | EFT | ENR | FFC | CLI | TIA | TIL | TUL | TET | SXT ** | ||||||
| A1 | 30.01.2020 | weaner | lung | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 2 | ˃16 | 32 | ˃128 | ˃128 | 2 | ≤0.06 |
| A2 | 15.04.2020 | weaner | brain | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 0.5 | ˃128 | ˃128 | ˃32 | ≤0.06 |
| A3 | 15.04.2020 | weaner | pleura | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | 0.5 | ˃128 | ˃128 | ˃32 | ≤0.06 |
| A4 | 05.06.2020 | weaner | lung | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 2 | ˃16 | 32 | ˃128 | ˃128 | 2 | ≤0.06 |
| A5 | 05.06.2020 | weaner | lung | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 2 | ˃16 | 1 | ˃128 | ˃128 | 2 | ≤0.06 |
| A6 | 13.10.2020 | weaner | brain | 2 | 54-like, aroA 1 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | ≤0.25 | ˃128 | ˃128 | 0.5 | ≤0.06 |
| A7 | 13.10.2020 | weaner | brain | 2 | 54-like, aroA 1 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | ≤0.25 | ˃128 | ˃128 | 8 | ≤0.06 |
| A8 | 13.10.2020 | weaner | brain | N.D. | N.D. | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 16 | ˃128 | ˃128 | 32 | ≤0.06 |
| A9 | 13.11.2020 | weaner | brain | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 2 | ˃16 | 32 | ˃128 | ˃128 | 2 | ≤0.06 |
| B1 | 19.06.2020 | weaner | heart | 3 | 54 | 0.06 | 0.06 | ≤0.25 | ≤0.125 | 0.25 | 2 | ˃16 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| B2 | 26.10.2020 | weaner | lung | 23 | N.D. | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 16 | ˃128 | ˃128 | 32 | ≤0.06 |
| B3 | 26.10.2020 | weaner | lung | N.D. | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ≤0.125 | 1 | 4 | ≤1 | 2 | ≤0.06 |
| B4 | 21.01.2021 | piglet | joint | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ≤0.125 | 1 | 4 | ≤1 | 2 | ≤0.06 |
| C1 | 16.07.2020 | weaner | lung | N.D. | 54 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| C2 | 26.01.2021 | fattener | brain | 31 | N.D. | 0.06 | 0.06 | ≤0.25 | ≤0.125 | 0.25 | 2 | ˃16 | ˃32 | ˃128 | ˃128 | 32 | ≤0.06 |
| D1 | 17.01.2020 | weaner | brain | 8 | 1546 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 4 | ˃128 | 128 | 32 | ≤0.06 |
| D2 | 26.06.2020 | weaner | heart | 3 | 54 | ≤0.03 | 0.06 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| D3 | 21.08.2020 | weaner | lung | 3 | 1521-like, aroA 8 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 2 | ˃128 | 64 | 16 | ≤0.06 |
| D4 | 27.10.2020 | weaner | brain | 23 | 54-like (dpr 223 2 diff. ) | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 16 | ˃128 | ˃128 | 32 | ≤0.06 |
| E1 | 27.01.2020 | weaner | lung | 2 | 1 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | 4 | ˃32 | 4 | ≤1 | 32 | ≤0.06 |
| F1 | 28.01.2020 | weaner | lung | N.D. | N.D. | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | ≤0.25 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| G1 | 22.01.2020 | weaner | eye | 16 | 1222-like (gki 294 1 diff.) | 0.25 | 0.06 | ≤0.25 | 0.5 | 0.25 | 1 | 8 | 16 | 2 | ≤1 | 32 | ≤0.06 |
| G2 | 20.08.2020 | weaner | lung | 8 | 87 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ≤0.125 | 0.5 | 2 | ≤1 | 0.5 | 1 |
| G3 | 01.09.2020 | weaner | lung | 3 | 54 | ≤0.03 | 0.06 | ≤0.25 | ≤0.125 | 0.5 | 1 | 8 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| G4 | 01.09.2020 | weaner | lung | N.D. | 912 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | 1 | 2 | 2 | ≤1 | 32 | ≤0.06 |
| G5 | 01.09.2020 | weaner | lung | 8 | 87 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ≤0.125 | 0.5 | 2 | ≤1 | 0.5 | 1 |
| H1 | 07.10.2020 | weaner | brain | 2 | 28 | ≤0.03 | 0.06 | ≤0.25 | ≤0.125 | 0.125 | 1 | ≤0.125 | 1 | 4 | ≤1 | 32 | ≤0.06 |
| H2 | 07.10.2020 | weaner | lung | N.D. | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ≤0.125 | 1 | 4 | ≤1 | 32 | ≤0.06 |
| I1 | 18.08.2020 | weaner | lung | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 1 | ˃128 | ˃128 | 32 | ≤0.06 |
| I2 | 18.08.2020 | weaner | heart | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ˃16 | 0.5 | ˃128 | ˃128 | 32 | ≤0.06 |
| J1 | 09.02.2021 | piglet | brain | 7 | 29 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 2 | ˃16 | 32 | ˃128 | ˃128 | 2 | ≤0.06 |
| K1 | 17.02.2020 | piglet | lung | 3 | 54 | ≤0.03 | 0.06 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| L1 | 10.02.2020 | fattener | lung | 8 | 87 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | 0.5 | ˃128 | ˃128 | ˃32 | ≤0.06 |
| M1 | 10.11.2020 | fattener | pericardium | 2 | 1 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 2 | ˃16 | 0.5 | ˃128 | ˃128 | 32 | ≤0.06 |
| N1 | 26.06.2020 | fattener | lung | N.D. | 54 | ≤0.03 | 0.06 | ≤0.25 | ≤0.125 | 0.25 | 1 | ˃16 | ˃32 | ˃128 | ˃128 | ≤0.25 | ≤0.06 |
| O1 | 12.11.2020 | piglet | lung | 16 | 1280-like (mutS 139 1 diff.) | 0.5 | 1 | ≤0.25 | 1 | 0.25 | 1 | ˃16 | 16 | ˃128 | ˃128 | ˃32 | 0.5 |
| O2 | 12.11.2020 | piglet | lung | 16 | 1280-like (mutS 139 1 diff.) | 0.5 | 1 | ≤0.25 | 1 | 0.25 | 1 | ˃16 | 16 | ˃128 | ˃128 | ˃32 | 0.5 |
| P1 | 23.01.2020 | piglet | joint | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.125 | 1 | ≤0.125 | 1 | 4 | 2 | ˃32 | ≤0.06 |
| P2 | 23.01.2020 | piglet | joint | 1/2 | 28 | ≤0.03 | ≤0.03 | ≤0.25 | ≤0.125 | 0.25 | 1 | ≤0.125 | 1 | 4 | 2 | ˃32 | ≤0.06 |
| Frequency of Resistance by | Phenotype of Resistance | Number of Isolates | Number of Multidrug-Resistant Isolates | |
|---|---|---|---|---|
| Active Substance | Antimicrobial Group | |||
| 0 | 0 | 2 | ||
| 1 | 1 | TET | 6 | |
| 2 | 2 | CLI, TET | 2 | |
| 3 | 2 | CLI, TIL, TUL | 2 | 0 |
| 3 | 3 | CLI, TIA, TET | 1 | |
| 4 | 3 | CLI, TIL, TUL, TET | 15 | |
| 4 | 3 | CLI, TIA, TIL, TUL | 6 | |
| 5 | 4 | CLI, TIA, TIL, TUL, TET | 5 | 27 |
| Farm | Sows (Framework Number) | Antimicrobials Used in | S. suis Isolates (n) | Isolates Resistant to TUL (n) | Multidrug Resistant Isolates (n) | |
|---|---|---|---|---|---|---|
| Piglets | Weaners | |||||
| A | 2000 | TUL | AMO | 9 | 9 | 8 |
| B | 2000 | TUL | - | 4 | 2 | 2 |
| C | 2000 | TUL | - | 2 | 2 | 2 |
| D | 2000 | TUL | AMO | 4 | 4 | 4 |
| E | 700 | TUL | AMO | 1 | 0 | 1 |
| F | 600 | TUL | AMO | 1 | 1 | 0 |
| G | 1000 | TUL | AMO | 5 | 1 | 1 |
| H | 1500 | - | AMO | 2 | 0 | 0 |
| I | 1100 | TUL | AMO | 2 | 2 | 2 |
| J | 1700 | - | AMO | 1 | 1 | 1 |
| K | 600 | TUL | AMO | 1 | 1 | 1 |
| L | 1500 | - | AMO | 1 | 1 | 1 |
| M | 500 | - | AMO | 1 | 1 | 1 |
| N | - | TUL | AMO | 1 | 1 | 1 |
| O | 500 | - | AMO | 2 | 2 | 2 |
| P | 2000 | - | AMO | 2 | 0 | 0 |
| Total | 19,700 | 10/16 | 14/16 | 39 (30 1; 9 2) | 28 (23 1; 5 2) | 27 (22 1; 5 2) |
| PEN | AMP | AMC * | EFT | ENR | FFC | CLI | TIA | TIL | TUL | TET | SXT ** |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 4 | 32 | 16 | 8 | 64 | 16 | 32 | 128 | 128 | 32 | PC |
| 2 | 2 | 16 | 8 | 4 | 32 | 8 | 16 | 64 | 64 | 16 | 4 |
| 1 | 1 | 8 | 4 | 2 | 16 | 4 | 8 | 32 | 32 | 8 | 2 |
| 0.5 | 0.5 | 4 | 2 | 1 | 8 | 2 | 4 | 16 | 16 | 4 | 1 |
| 0.25 | 0.25 | 2 | 1 | 0.5 | 4 | 1 | 2 | 8 | 8 | 2 | 0.5 |
| 0.125 | 0.125 | 1 | 0.5 | 0.25 | 2 | 0.5 | 1 | 4 | 4 | 1 | 0.25 |
| 0.06 | 0.06 | 0.5 | 0.25 | 0.125 | 1 | 0.25 | 0.5 | 2 | 2 | 0.5 | 0.125 |
| 0.03 | 0.03 | 0.25 | 0.125 | 0.06 | 0.5 | 0.125 | 0.25 | 1 | 1 | 0.25 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matiašovic, J.; Nedbalcová, K.; Žižlavský, M.; Fleischer, P.; Pokludová, L.; Kellnerová, D.; Nechvátalová, K.; Šimek, B.; Czanderlová, L.; Zouharová, M.; et al. Streptococcus suis Isolates—Serotypes and Susceptibility to Antimicrobials in Terms of Their Use on Selected Repopulated Czech Pig Farms. Pathogens 2021, 10, 1314. https://doi.org/10.3390/pathogens10101314
Matiašovic J, Nedbalcová K, Žižlavský M, Fleischer P, Pokludová L, Kellnerová D, Nechvátalová K, Šimek B, Czanderlová L, Zouharová M, et al. Streptococcus suis Isolates—Serotypes and Susceptibility to Antimicrobials in Terms of Their Use on Selected Repopulated Czech Pig Farms. Pathogens. 2021; 10(10):1314. https://doi.org/10.3390/pathogens10101314
Chicago/Turabian StyleMatiašovic, Ján, Kateřina Nedbalcová, Marek Žižlavský, Petr Fleischer, Lucie Pokludová, Dita Kellnerová, Kateřina Nechvátalová, Bronislav Šimek, Linda Czanderlová, Monika Zouharová, and et al. 2021. "Streptococcus suis Isolates—Serotypes and Susceptibility to Antimicrobials in Terms of Their Use on Selected Repopulated Czech Pig Farms" Pathogens 10, no. 10: 1314. https://doi.org/10.3390/pathogens10101314
APA StyleMatiašovic, J., Nedbalcová, K., Žižlavský, M., Fleischer, P., Pokludová, L., Kellnerová, D., Nechvátalová, K., Šimek, B., Czanderlová, L., Zouharová, M., Bernardy, J., Králová, N., & Šlosárková, S. (2021). Streptococcus suis Isolates—Serotypes and Susceptibility to Antimicrobials in Terms of Their Use on Selected Repopulated Czech Pig Farms. Pathogens, 10(10), 1314. https://doi.org/10.3390/pathogens10101314