Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures


Abstract
1. Introduction
2. Materials and Methods
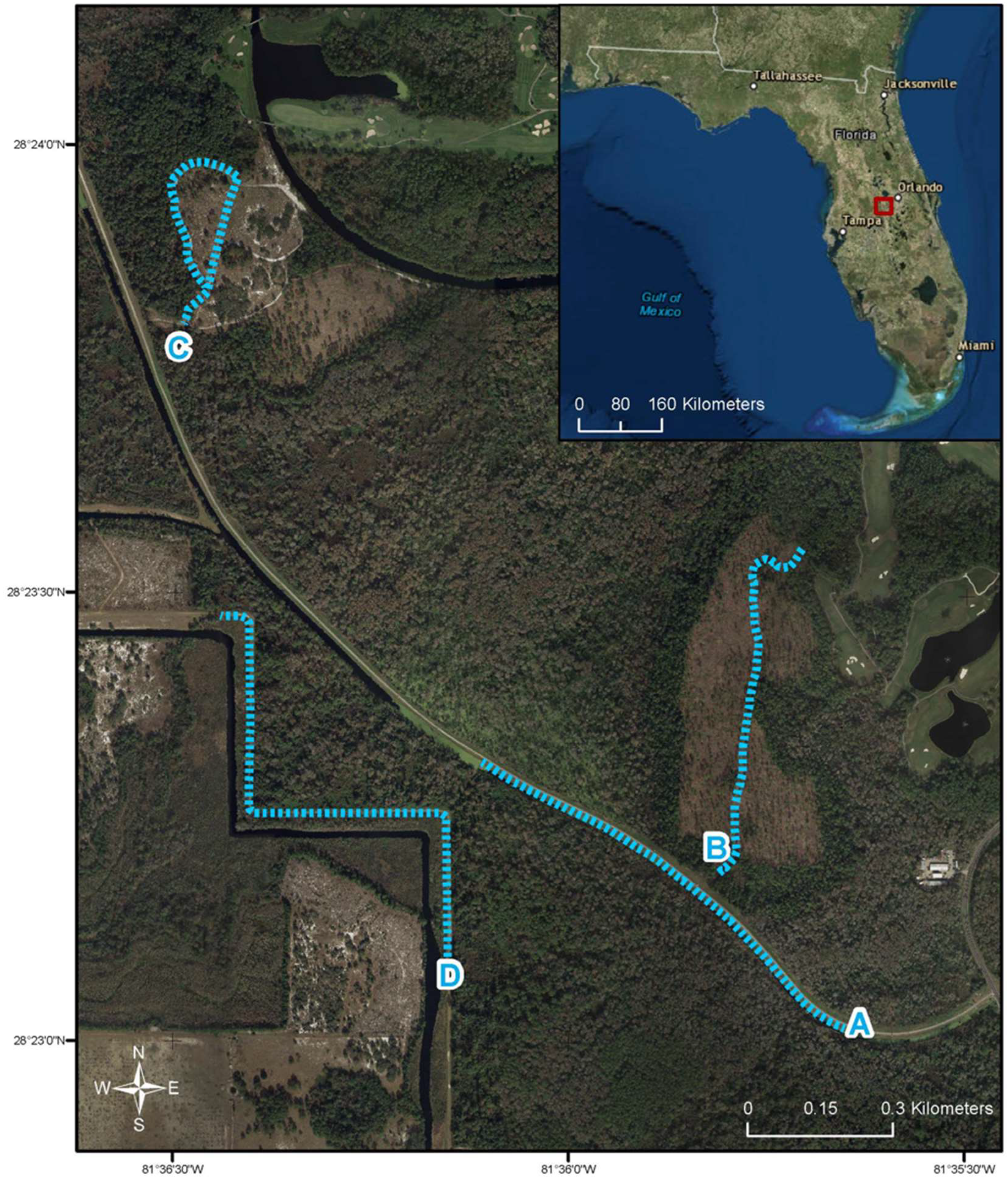
2.1. Study System
2.2. Sampling Design
2.3. Analyses
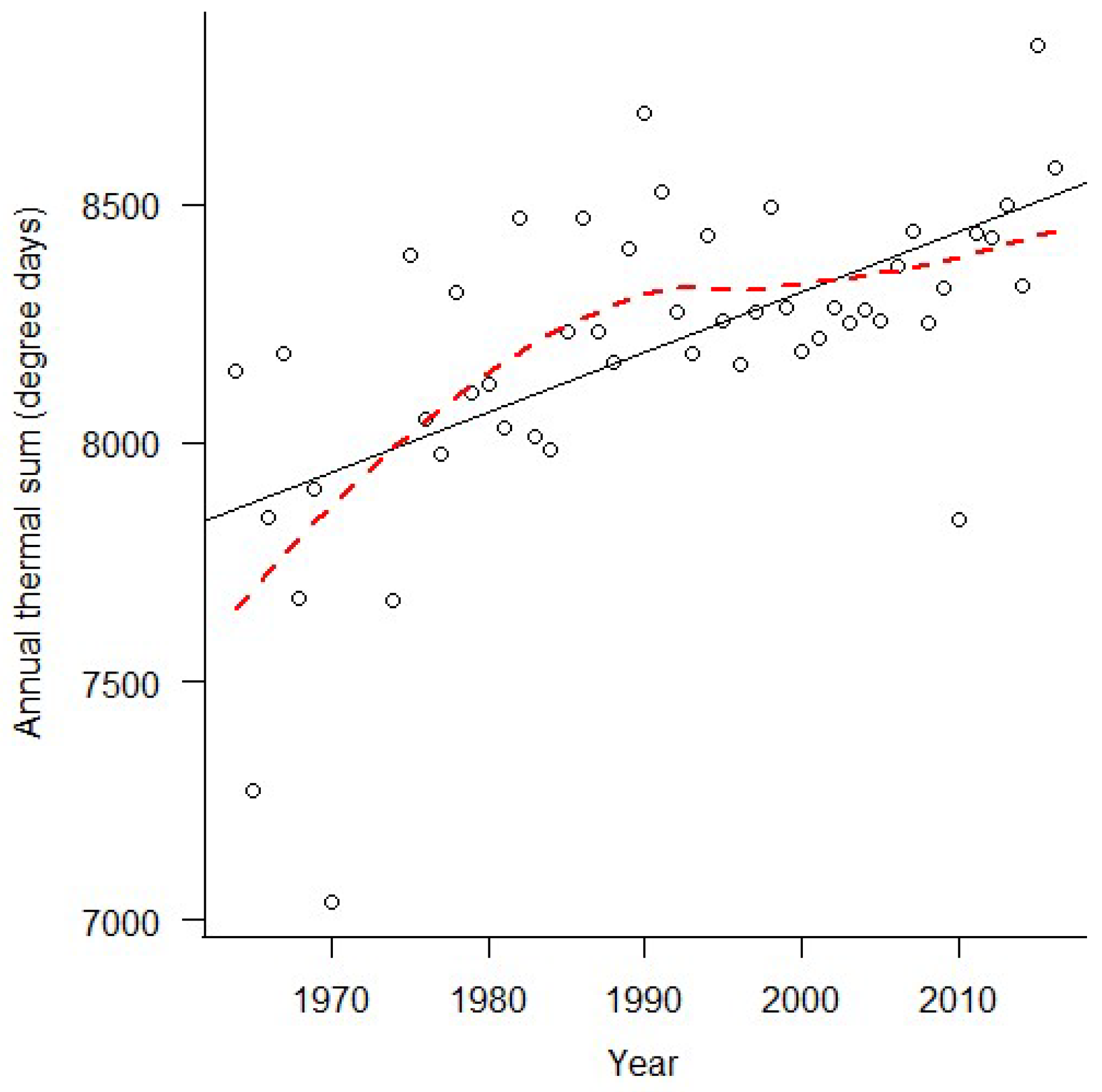
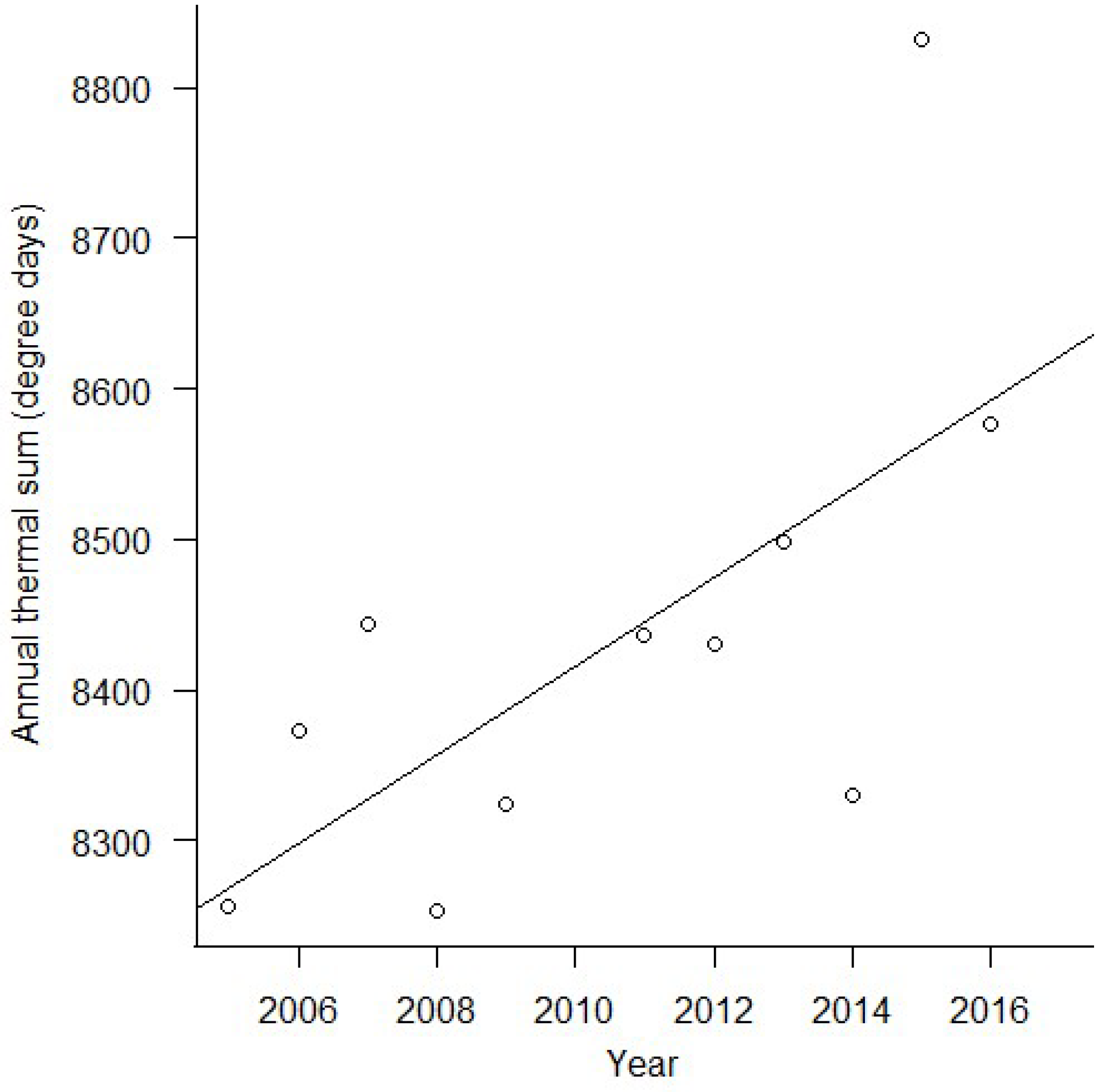
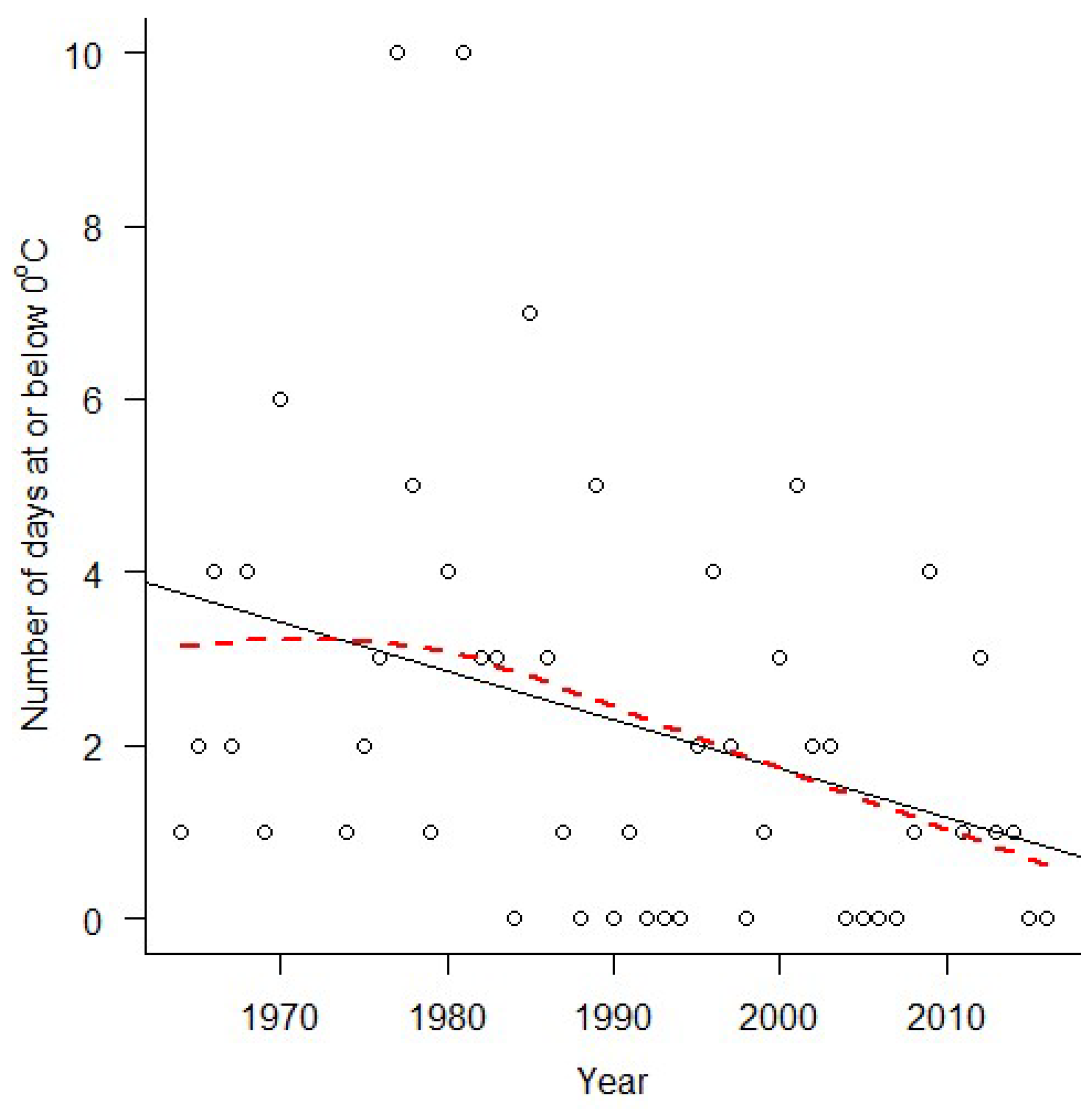
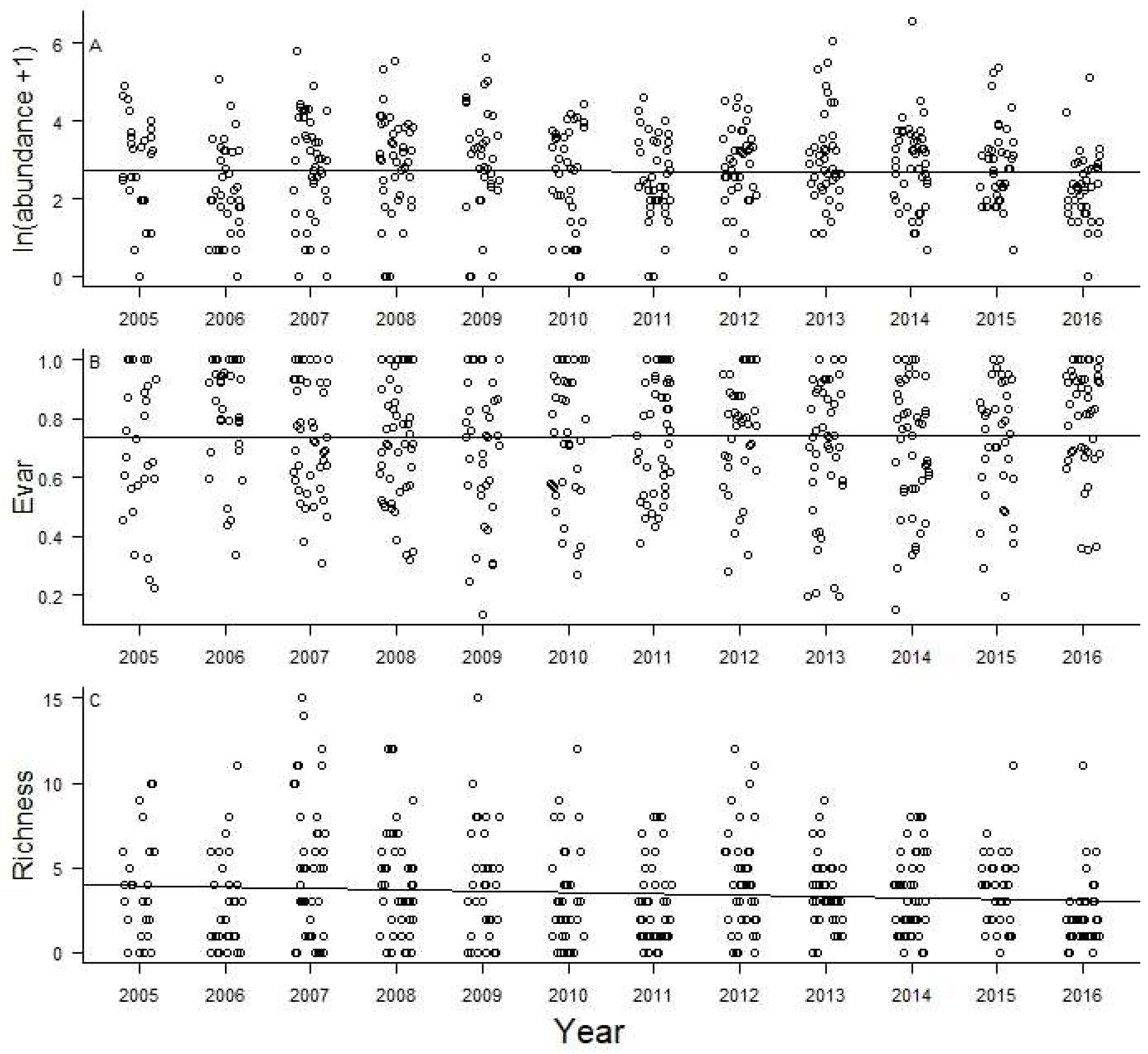
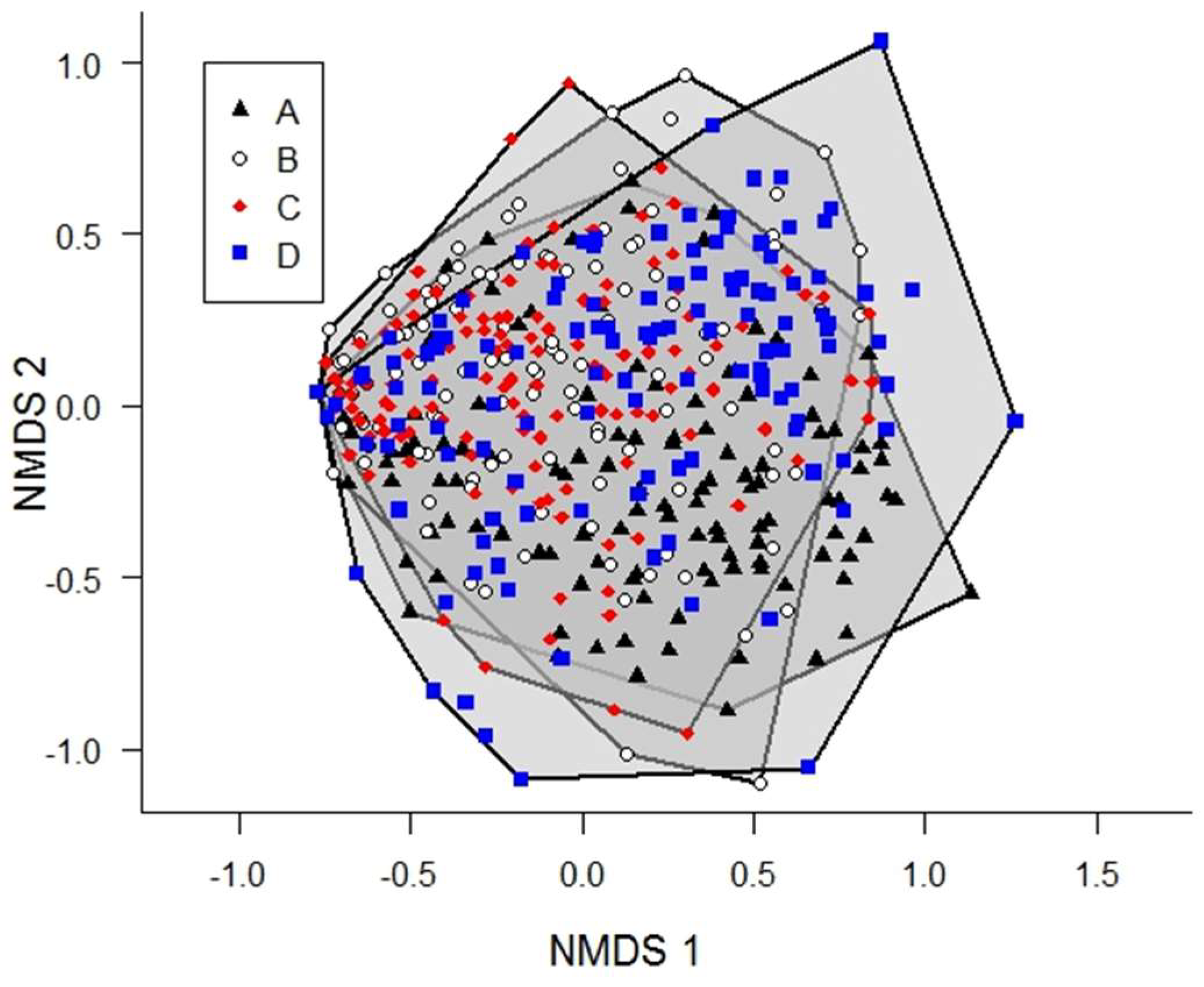
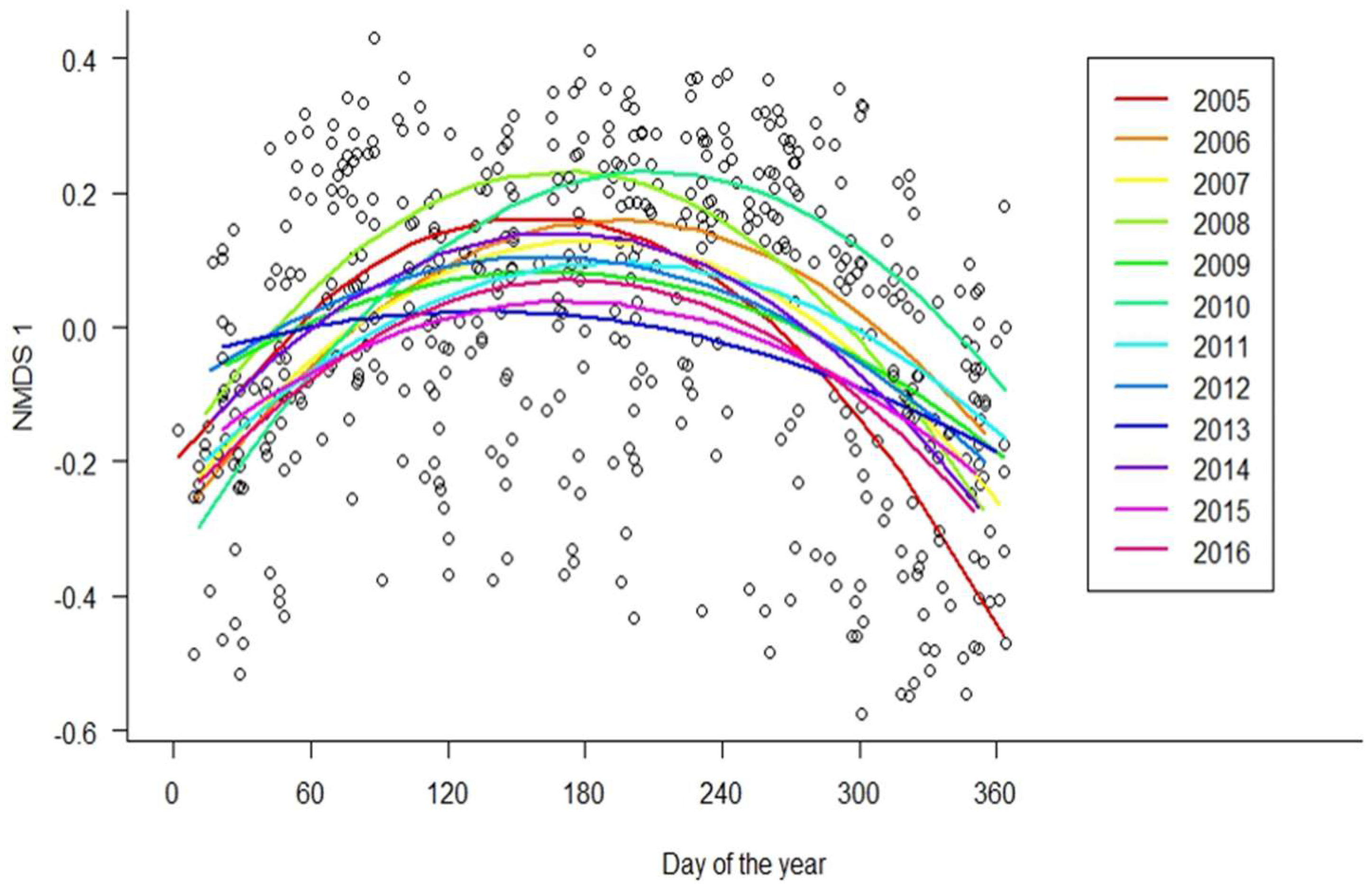
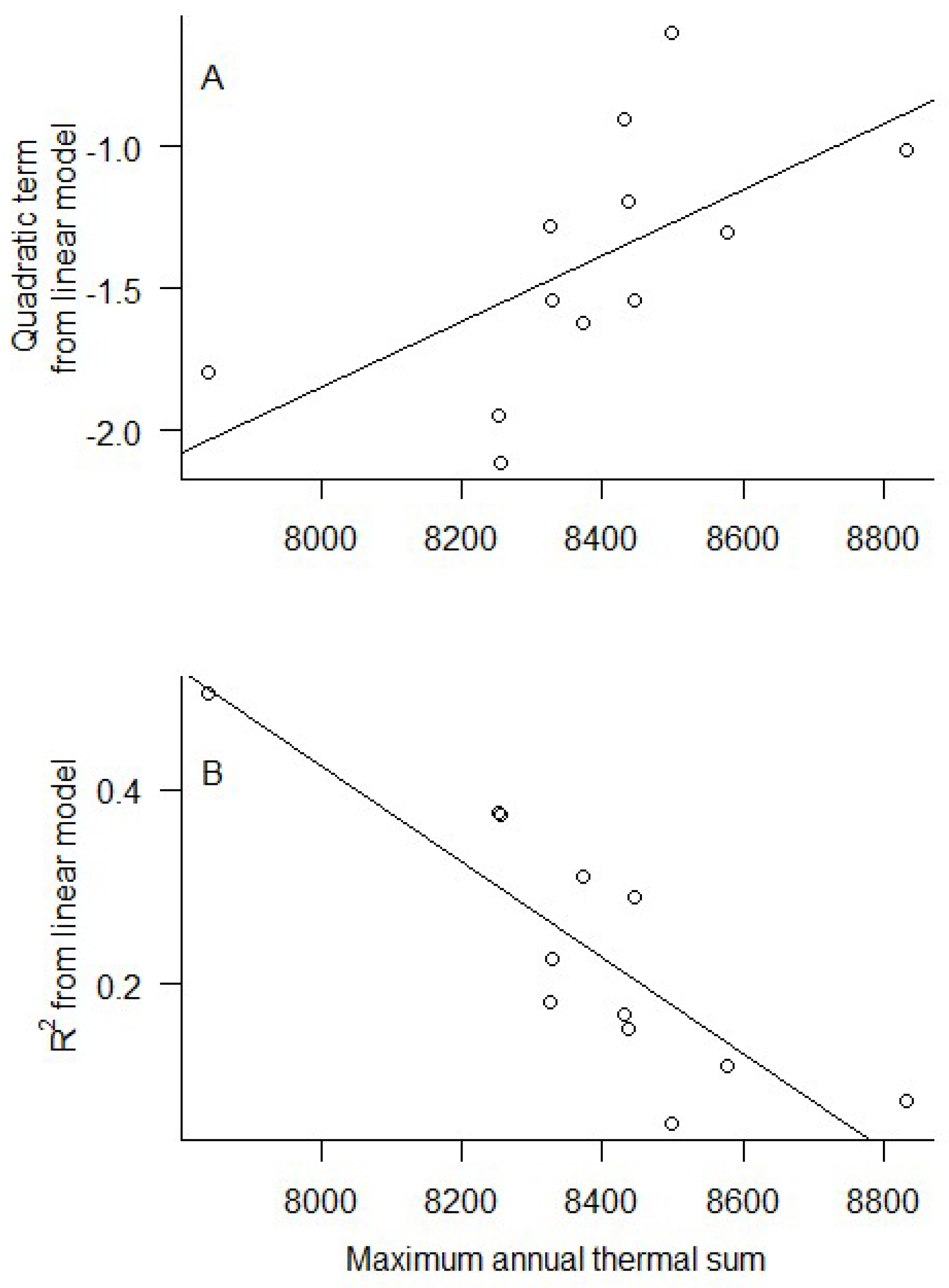
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L. Biological consequences of global warming: Is the signal already apparent? Trends Ecol. Evol. 2000, 15, 56–61. [Google Scholar] [CrossRef]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will Climate Change Promote Alien Plant Invasions. In Biological Invasions; Nentwig, D.W., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-77375-7. [Google Scholar]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. In Annual Review of Ecology Evolution and Systematics; Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Alan Pounds, J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.; Miller-Rushing, A.J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3101–3112. [Google Scholar] [CrossRef] [PubMed]
- Cayton, H.L.; Haddad, N.M.; Gross, K.; Diamond, S.E.; Ries, L. Do growing degree days predict phenology across butterfly species? Ecology 2015, 96, 1473–1479. [Google Scholar] [CrossRef]
- Kimball, S.; Angert, A.L.; Huxman, T.E.; Venable, D.L. Contemporary climate change in the Sonoran Desert favors cold-adapted species. Glob. Change Biol. 2010, 16, 1555–1565. [Google Scholar] [CrossRef]
- Guo, H.; Yang, H.; Mockler, T.C.; Lin, C. Regulation of flowering time by Arabidopsis photoreceptors. Science 1998, 279, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.M.; Miller-Rushing, A.J.; Inouye, D.W. Changes in snowmelt date and summer precipitation affect the flowering phenology of Erythronium grandiflorum (glacier lily; Liliaceae). Am. J. Bot. 2010, 97, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Harte, J.; Taylor, K.J. Subalpine meadow flowering phenology responses to climate change: integrating experimental and gradient methods. Ecol. Monogr. 2003, 73, 69–86. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B. Global warming and flowering times in Thoreau’s concord: A community perspective. Ecology 2008, 89, 332–341. [Google Scholar] [CrossRef] [PubMed]
- CaraDonna, P.J.; Iler, A.M.; Inouye, D.W. Shifts in flowering phenology reshape a subalpine plant community. Proc. Natl. Acad. Sci. 2014, 111, 4916–4921. [Google Scholar] [CrossRef] [PubMed]
- Gezon, Z.J.; Inouye, D.W.; Irwin, R.E. Phenological change in a spring ephemeral: implications for pollination and plant reproduction. Glob. Change Biol. 2016, 22, 1779–1793. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.S.; Etterson, J.R.; Galloway, L.F. Artificial selection shifts flowering phenology and other correlated traits in an autotetraploid herb. Heredity 2007, 99, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.T.; Inouye, D.W.; McKinney, A.M.; Colautti, R.I.; Mitchell-Olds, T. Phenotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change. Proc. R. Soc. B Biol. Sci. 2012, 279, 3843–3852. [Google Scholar] [CrossRef] [PubMed]
- McKinney, A.M.; CaraDonna, P.J.; Inouye, D.W.; Barr, B.; Bertelsen, C.D.; Waser, N.M. Asynchronous changes in phenology of migrating Broad-tailed Hummingbirds and their early-season nectar resources. Ecology 2012, 93, 1987–1993. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.; Inouye, D.W.; Thomson, J.D. Flowering phenology in subalpine meadows: Does climate variation influence community co-flowering patterns? Ecology 2010, 91, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Bonebrake, T.C.; Ponisio, L.C.; Boggs, C.L.; Ehrlich, P.R. More than just indicators: A review of tropical butterfly ecology and conservation. Biol. Conserv. 2010, 143, 1831–1841. [Google Scholar] [CrossRef]
- Inouye, D.W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 2008, 89, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Altermatt, F. Tell me what you eat and I’ll tell you when you fly: diet can predict phenological changes in response to climate change. Ecol. Lett. 2010, 13, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.H. Why mountain passes are higher in the tropics. Am. Nat. 1967, 101, 233–249. [Google Scholar] [CrossRef]
- Diamond, S.E.; Chick, L.; Penick, C.A.; Nichols, L.M.; Cahan, S.H.; Dunn, R.R.; Ellison, A.M.; Sanders, N.J.; Gotelli, N.J. Heat tolerance predicts the importance of species interaction effects as the climate changes. Integr. Comp. Biol. 2017, 57, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Pollard, E. A method of assessing the abundance of butterflies in Monks Wood National Nature Reserve in 1973. Ent. Gaz. 1975, 26, 79–88. [Google Scholar]
- Pollard, E. A method for assessing changes in the abundance of butterflies. Biol. Conserv. 1977, 12, 115–134. [Google Scholar] [CrossRef]
- Smith, B.; Wilson, J.B. A consumer’s guide to evenness indices. Oikos 1996, 76, 70–82. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 12 September 2018).
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0. [Google Scholar]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q) AIC (c). Available online: https://cran.r-project.org/web/packages/AICcmodavg/index.html (accessed on 12 September 2018).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Dai, B.; Grothendieck, G.; Green, P. lme4: Linear Mixed-Effects Models using “Eigen” and S4. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 12 September 2018).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest: Tests in Linear Mixed Effects Models. Available online: https://cran.r-project.org/web/packages/lmerTest/index.html (accessed on 12 September 2018).
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Lefcheck, J. piecewiseSEM: Piecewise structural equation modeling. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Wickham, H. RStudio scales: Scale Functions for Visualization. Available online: https://cran.r-project.org/web/packages/scales/index.html (accessed on 8 May 2018).
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2016. [Google Scholar]
- Cavanaugh, K.C.; Kellner, J.R.; Forde, A.J.; Gruner, D.S.; Parker, J.D.; Rodriguez, W.; Feller, I.C. Poleward expansion of mangroves is a threshold response to decreased frequency of extreme cold events. Proc. Natl. Acad. Sci. 2014, 111, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Holle, B.V.; Wei, Y.; Nickerson, D. Climatic Variability Leads to Later Seasonal Flowering of Floridian Plants. PLOS ONE 2010, 5, e11500. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Change Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Stålhandske, S.; Gotthard, K.; Leimar, O. Winter chilling speeds spring development of temperate butterflies. J. Anim. Ecol. 2017, 86, 718–729. [Google Scholar] [CrossRef]
- Donnelly, A.; Caffarra, A.; O’Neill, B.F. A review of climate-driven mismatches between interdependent phenophases in terrestrial and aquatic ecosystems. Int. J. Biometeorol. 2011, 55, 805–817. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | K | AICc | ΔAICc | wi | cwi | Log L |
|---|---|---|---|---|---|---|
| doy + doy2 | 4 | −162.46 | 0 | 0.73 | 0.73 | 85.27 |
| doy + precipitation + doy2 | 5 | −160.43 | 2.03 | 0.27 | 1 | 85.28 |
| Precipitation × doy2 | 4 | −95.4 | 67.06 | 0 | 1 | 51.74 |
| Precipitation | 3 | −75.01 | 87.45 | 0 | 1 | 40.53 |
| doy | 4 | −73.88 | 88.58 | 0 | 1 | 40.98 |
| doy2 | 3 | −70.71 | 91.76 | 0 | 1 | 38.38 |
| precipitation + precipitation2 | 3 | −56.8 | 105.66 | 0 | 1 | 31.43 |
| Model | K | AICc | ΔAICc | wi | cwi | log L |
|---|---|---|---|---|---|---|
| route | 5 | −378.46 | 0 | 0.54 | 0.54 | 194.29 |
| effort + route | 6 | −376.42 | 2.04 | 0.19 | 0.74 | 194.3 |
| route + temperature | 6 | −376.41 | 2.05 | 0.19 | 0.93 | 194.29 |
| effort + route + temperature | 7 | −374.36 | 4.1 | 0.07 | 1 | 194.3 |
| effort | 5 | −148.75 | 229.71 | 0 | 1 | 79.44 |
| effort + temperature | 6 | −146.7 | 231.77 | 0 | 1 | 79.44 |
| temperature | 5 | −145.55 | 232.91 | 0 | 1 | 77.84 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gezon, Z.J.; Lindborg, R.J.; Savage, A.; Daniels, J.C. Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures. Insects 2018, 9, 174. https://doi.org/10.3390/insects9040174
Gezon ZJ, Lindborg RJ, Savage A, Daniels JC. Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures. Insects. 2018; 9(4):174. https://doi.org/10.3390/insects9040174
Chicago/Turabian StyleGezon, Zachariah J., Rebekah J. Lindborg, Anne Savage, and Jaret C. Daniels. 2018. "Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures" Insects 9, no. 4: 174. https://doi.org/10.3390/insects9040174
APA StyleGezon, Z. J., Lindborg, R. J., Savage, A., & Daniels, J. C. (2018). Drifting Phenologies Cause Reduced Seasonality of Butterflies in Response to Increasing Temperatures. Insects, 9(4), 174. https://doi.org/10.3390/insects9040174