Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant


Abstract

1. Introduction
2. Materials and Methods
2.1. Study Organism
2.2. Experimental Colonies
2.3. Retrieval of Intact Pupae
2.4. Activity of Pupal Extracts
2.5. Activity of Fipronil-Treated Ant Cadaver Extracts
2.6. Response Variables
2.7. Statistical Analysis
2.7.1. Activity of Pupal Extracts
2.7.2. Activity of Fipronil-Treated Ant Cadaver Extracts
3. Results
3.1. Pupa Retrieval Time
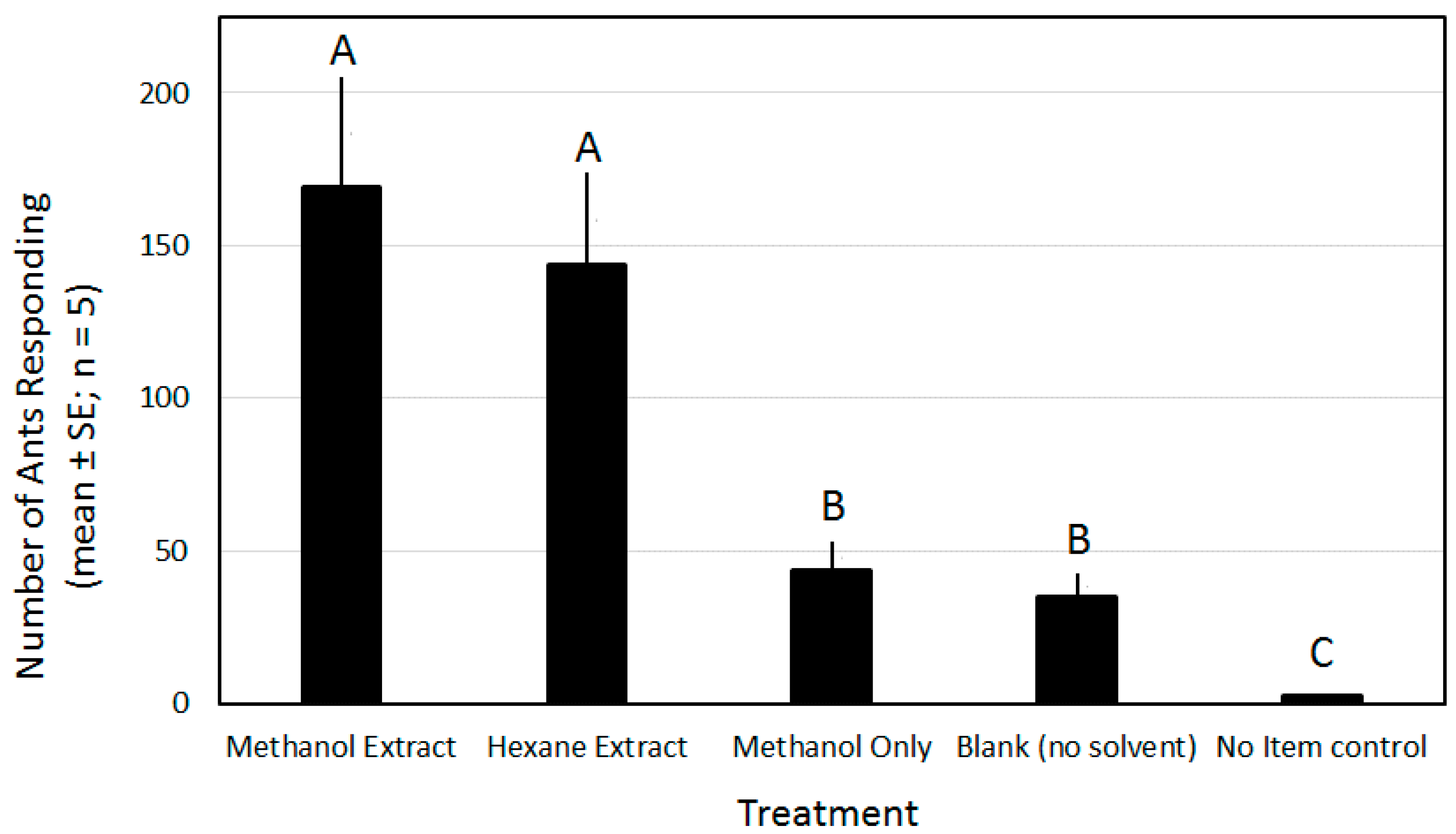
3.2. Activity of Pupal Extracts
3.3. Activity of Fipronil-Treated Ant Cadaver Extracts
4. Discussion
4.1. Pupa Retrieval Time
4.2. Activity of Pupal Extracts
4.3. Activity of Fipronil-Treated Ant Cadaver Extracts
Author Contributions
Funding
Conflicts of Interest
References
- Vega, S.J.; Rust, M.K. The Argentine ant—A significant invasive species in agricultural, urban and natural environments. Sociobiology 2001, 37, 3–25. [Google Scholar]
- Suarez, A.V.; Holway, D.A.; Tsutsui, N.D. Genetics and behavior of a colonizing species: The invasive Argentine ant. Am. Nat. 2008, 172, S72–S84. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, G.M.; Silverman, J. Queen acceptance and the complexity of nestmate discrimination in the Argentine ant. Behav. Ecol. Sociobiol. 2008, 62, 537–548. [Google Scholar] [CrossRef]
- Wiltz, B.A.; Suiter, D.R.; Gardner, W.A. Activity of bifenthrin, chlorfenapyr, fipronil, and thiamethoxam against Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2009, 102, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K.; Reierson, D.A.; Klotz, J.H. Pest management of Argentine ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2003, 38, 159–169. [Google Scholar] [CrossRef]
- Choe, D.H.; Rust, M.K. Horizontal transfer of insecticides in laboratory colonies of the Argentine ant (Hymenoptera: Formicidae). J. Econ. Entomol. 2008, 101, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Hooper-Bui, L.M.; Rust, M.K. Oral toxicity of abamectin, boric acid, fipronil, and hydramethylnon to laboratory colonies of Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2000, 93, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Hooper-Bui, L.M.; Kwok, E.S.C.; Buchholz, B.A.; Rust, M.K.; Eastmond, D.A.; Vogel, J.S. Insecticide transfer efficiency and lethal load in Argentine ants. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2015, 361, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.; Reierson, D.A.; Klotz, J.H. Delayed toxicity as a critical factor in the efficacy of aqueous baits for controlling Argentine ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2004, 97, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.; Reierson, D.A.; Klotz, J.H. Factors affecting the performance of bait toxicants for Argentine ants (Hymenoptera: Formicidae). In Proceedings of the 4th International Conference on Urban Pests, Charleston, SC, USA, 7–10 July 2002; Jones, S.C., Zhai, J., Robinson, W.H., Eds.; Pocahontas Press, Inc.: Blacksburg, VA, USA, 2002; pp. 115–120. [Google Scholar]
- Vega, S.Y.; Rust, M.K. Determining the foraging range and origin of resurgence after treatment of Argentine ant (Hymenoptera: Formicidae) in urban areas. J. Econ. Entomol. 2003, 96, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.H.; Rust, M.K.; Costa, H.S.; Reierson, D.A.; Kido, K. Strategies for controlling Argentine ants (Hymenoptera: Formicidae) with sprays and baits. J. Agric. Urban Entomol. 2002, 19, 85–94. [Google Scholar]
- Gan, J.; Bondarenko, S.; Oki, L.; Haver, D.; Li, J.X. Occurrence of fipronil and its biologically active derivatives in urban residential runoff. Environ. Sci. Technol. 2012, 46, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.Y.; Soeprono, A.; Rust, M.K.; Gan, J. Ant control efficacy of pyrethroids and fipronil on outdoor concrete surfaces. Pest Manag. Sci. 2014, 70, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, J.P.; Rouse, D.R.; Avants, J.K.; Garrison, A.W.; Delorenzo, M.E.; Chung, K.W.; Key, P.B.; Wilson, W.A.; Black, M.C. Toxicity of fipronil and its enantiomers to marine and freshwater non-targets. J. Environ. Sci. Health Part B 2007, 42, 471–480. [Google Scholar] [CrossRef]
- Pisa, L.W.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Downs, C.A.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; McField, M.; et al. Effects of neonicotinoids and fipronil on non-target invertebrates. Environ. Sci. Pollut. Res. 2015, 22, 68–102. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, L.; Rust, M.K.; Klotz, J.H.; Haver, D.; Kabashima, J.N.; Bondarenko, S.; Gan, J. Impact of ant control technologies on insecticide runoff and efficacy. Pest Manag. Sci. 2010, 66, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Wiltz, B.A.; Suiter, D.R.; Gardner, W.A. Deterrency and toxicity of essential oils to Argentine and Red Imported Fire ants (Hymenoptera: Formicidae). J. Entomol. Sci. 2007, 42, 239–249. [Google Scholar] [CrossRef]
- Guerra, M.D.; Suiter, D.R.; Scocco, C.M. Topical toxicity of nine essential oils to Camponotus pennsylvanicus (Hymenoptera: Formicidae). Sociobiology 2011, 58, 419–426. [Google Scholar]
- Scocco, C.M.; Suiter, D.R.; Gardner, W.A. Repellency of five essential oils to Linepithema humile (Hymenoptera: Formicidae). J. Entomol. Sci. 2012, 47, 150–159. [Google Scholar] [CrossRef]
- Greenberg, L.; Klotz, J.H. Argentine ant (Hymenoptera: Formicidae) trail pheromone enhances consumption of liquid sucrose solution. J. Econ. Entomol. 2000, 93, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Tsai, K.; Lopez, C.M.; Campbell, K. Pheromone-assisted techniques to improve the efficacy of insecticide sprays against Linepithema humile (Hymenoptera: Formicidae). J. Econ. Entomol. 2014, 107, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Peck, R.W.; Manning, L.M.; Stringer, L.D.; Cappadonna, J.; El-Sayed, A.M. Pheromone disruption of Argentine ant trail integrity. J. Chem. Ecol. 2008, 34, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Peck, R.W.; Stringer, L.D.; Snook, K.; Banko, P.C. Trail pheromone disruption of Argentine ant trail formation and foraging. J. Chem. Ecol. 2010, 36, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Stringer, L.D.; Corn, J.E. Argentine ant trail pheromone disruption is mediated by trail concentration. J. Chem. Ecol. 2011, 37, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Klotz, J.H.; Rust, M.K.; Greenberg, L.; Robertson, M.A. Developing low risk management strategies for Argentine ants (Hymenoptera: Formicidae). Sociobiology 2010, 55, 779–785. [Google Scholar]
- Buczkowski, G. Prey-baiting as a conservation tool: Selective control of invasive ants with minimal non-target effects. Insect Conserv. Divers. 2017, 10, 302–309. [Google Scholar] [CrossRef]
- Hughes, L.; Westoby, M. Capitula on stick insect eggs and elaiosomes on seeds: Convergent adaptations for burial by ants. Funct. Ecol. 1992, 6, 642–648. [Google Scholar] [CrossRef]
- Windsor, D.M.; Trapnell, D.W.; Amat, G. The egg capitulum of a Neotropical walkingstick, Calynda bicuspis, induces aboveground egg dispersal by the ponerine ant, Ectatomma ruidum. J. Insect Behav. 1996, 9, 353–367. [Google Scholar] [CrossRef]
- Brew, C.R.; O’Dowd, D.J.; Rae, I.D. Seed dispersal by ants: Behaviour-releasing compounds in elaiosomes. Oecologia 1989, 80, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.H.; Millar, J.G.; Rust, M.K. Chemical signals associated with life inhibit necrophoresis in Argentine ants. Proc. Natl. Acad. Sci. USA 2009, 106, 8251–8255. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS/STAT Software Version 14.1; SAS Institute Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Walsh, J.P.; Tschinkel, W.R. Brood recognition by contact pheromone in the red imported fire ant, Solenopsis invicta. Anim. Behav. 1974, 22, 695–704. [Google Scholar] [CrossRef]
- Bonavitacougourdan, A.; Clement, J.L.; Lange, C. Larvae recognition in the Camponotus vagus Scop ant—Larvae phenotypes of cuticular hydrocarbon patterns. C. R. Acad. Sci. Ser. Sci. Vie-Life Sci. 1988, 306, 299–305. [Google Scholar]
- Bonavitacougourdan, A.; Clement, J.L.; Lange, C. The role of cuticular hydrocarbons in recognition of larvae by workers of the ant Camponotus vagus—Changes in the chemical signature in response to social-environment (Hymenoptera: Formicidae). Sociobiology 1989, 16, 49–74. [Google Scholar]
- Van Wilgenburg, E.; Sulc, R.; Shea, K.J.; Tsutsui, N.D. Deciphering the chemical basis of nestmate recognition. J. Chem. Ecol. 2010, 36, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Shimoji, H.; Fujiki, Y.; Yamaoka, R.; Tsuji, K. Egg discrimination by workers in Diacamma sp from Japan. Insectes Soc. 2012, 59, 201–206. [Google Scholar] [CrossRef]
- Helantera, H.; Martin, S.J.; Ratnieks, F.L.W. Recognition of nestmate eggs in the ant Formica fusca is based on queen derived cues. Curr. Zool. 2014, 60, 131–136. [Google Scholar] [CrossRef]
- Krasnec, M.O.; Breed, M.D. Colony-specific cuticular hydrocarbon profile in Formica argentea ants. J. Chem. Ecol. 2013, 39, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sainz-Borgo, C.; Cabrera, A.; Hernandez, J.V. Nestmate recognition in the ant Odontomachus bauri (Hymenoptera: Formicidae). Sociobiology 2011, 58, 701–718. [Google Scholar]
- Bigley, W.S.; Vinson, S.B. Characterization of a brood pheromone isolated from the sexual brood of the imported fire ant, Solenopsis invicta. Ann. Entomol. Soc. Am. 1975, 68, 301–304. [Google Scholar] [CrossRef]
- Tschinkel, W.R. Nestmate and brood recognition. In The Fire Ants; Belknap: Cambridge, MA, USA, 2006; pp. 263–268. ISBN 0-674-02207-6. [Google Scholar]
- Wiltz, B.A.; Suiter, D.; Gardner, W.; Berisford, C.W. A novel delivery method for ant (Hymenoptera: Formicidae) toxicants. Midsouth Entomol. 2010, 3, 79–88. [Google Scholar]
- Glancey, B.M.; Stringer, C.E.; Craig, C.H.; Bishop, P.M.; Martin, B.B. Pheromone may induce brood tending in the fire ant, Solenopsis saevissima. Nature 1970, 226, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.L.; Beattie, A.J.; Bollenbacher, W.E. Evidence for diglycerides as attractants in an ant-seed interaction. J. Chem. Ecol. 1979, 5, 335–344. [Google Scholar] [CrossRef]
- Buczkowski, G. The Trojan horse approach for managing invasive ants: A study with Asian needle ants, Pachycondyla chinensis. Biol. Invasions 2016, 18, 507–515. [Google Scholar] [CrossRef]
- Buczkowski, G.; Mothapo, N.P.; Wossler, T.C. Let them eat termites—Prey-baiting provides effective control of Argentine ants, Linepithema humile, in a biodiversity hotspot. J. Appl. Entomol. 2018, 142, 504–512. [Google Scholar] [CrossRef]
- Pulliainen, U.; Bos, N.; d’Errorre, P.; Sundström, L. Caste-dependent brood retrieval by workers in the ant Formica exsecta. Anim. Behav. 2018, 140, 151–159. [Google Scholar] [CrossRef]
- Mahler, B.J.; Van Metre, P.C.; Wilson, J.T.; Musgrove, M.; Zaugg, S.D.; Burkhardt, M.R. Fipronil and its degradates in indoor and outdoor dust. Environ. Sci. Technol. 2009, 43, 5665–5670. [Google Scholar] [CrossRef] [PubMed]
- Hayasaka, D.; Kuwayama, N.; Takeo, A.; Ishida, T.; Mano, H.; Inoue, M.N.; Nagai, T.; Sanchez-Bayo, F.; Goka, K.; Sawahata, T. Different acute toxicity of fipronil baits on invasive Linepithema humile supercolonies and some non-target ground arthropods. Ecotoxicology 2015, 24, 1221–1228. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Hour 1 | Hour 2 | Hour 24 |
|---|---|---|---|
| Ant Cadavers | 26.1 ± 2.6 a | 27.8 ± 2.2 a | 30.0 ± 2.2 a |
| 0.001% fipronil + cadavers | 26.8 ± 2.6 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.01% fipronil + cadavers | 26.8 ± 2.6 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.10% fipronil + cadavers | 25.9 ± 2.5 a | 27.8 ± 2.2 a | 30.0 ± 2.2 a |
| 0.001% fipronil + cadaver extract + wicks | 27.7 ± 2.7 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.01% fipronil + cadaver extract + wicks | 29.6 ± 2.8 a | 30.0 ± 2.3 a | 30.0 ± 2.2 a |
| 0.10% fipronil + cadaver extract + wicks | 25.9 ± 2.5 a | 26.5 ± 2.2 a | 27.0 ± 2.1 a |
| 0.001% fipronil + wicks | 0.7 ± 0.3 b | 0.8 ± 0.4 c | 7.0 ± 1.1 c |
| 0.01% fipronil + wicks | 1.5 ± 0.5 b | 2.0 ± 0.6 bc | 4.8 ± 0.9 c |
| 0.10% fipronil + wicks | 1.7 ± 0.5 b | 2.7 ± 0.7 b | 13.7 ± 1.5 b |
| Treatment | Percent Mortality (Mean ± SE) |
|---|---|
| Ant cadavers | 15.1 ± 4.3 g |
| 0.001% fipronil + cadavers | 19.3 ± 1.2 f |
| 0.01% fipronil + cadavers | 58.9 ± 7.3 b |
| 0.10% fipronil + cadavers | 84.9 ± 4 a |
| 0.001% fipronil + cadaver extract + wicks | 14.0 ± 2.1 g |
| 0.01% fipronil + cadaver extract + wicks | 13.5 ± 1.5 g |
| 0.10% fipronil + cadaver extract + wicks | 43.9 ± 7.9 d |
| 0.001% fipronil + wicks | 18.3 ± 6.2 f |
| 0.01% fipronil + wicks | 25.9 ± 5.1 e |
| 0.10% fipronil + wicks | 51.9 ± 5.9 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gochnour, B.M.; Suiter, D.R.; Davis, J.W.; Huang, Q. Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects 2018, 9, 171. https://doi.org/10.3390/insects9040171
Gochnour BM, Suiter DR, Davis JW, Huang Q. Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects. 2018; 9(4):171. https://doi.org/10.3390/insects9040171
Chicago/Turabian StyleGochnour, Benjamin M., Daniel R. Suiter, Jerry W. Davis, and Qingguo Huang. 2018. "Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant" Insects 9, no. 4: 171. https://doi.org/10.3390/insects9040171
APA StyleGochnour, B. M., Suiter, D. R., Davis, J. W., & Huang, Q. (2018). Use of an Argentine Ant, Linepithema humile, Semiochemical to Deliver an Acute Toxicant. Insects, 9(4), 171. https://doi.org/10.3390/insects9040171