Climatic Variation of Supercooling Point in the Linden Bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae)


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Organism
2.2. Experimental Design
2.3. Climatic Data
2.4. Statistical Analysis
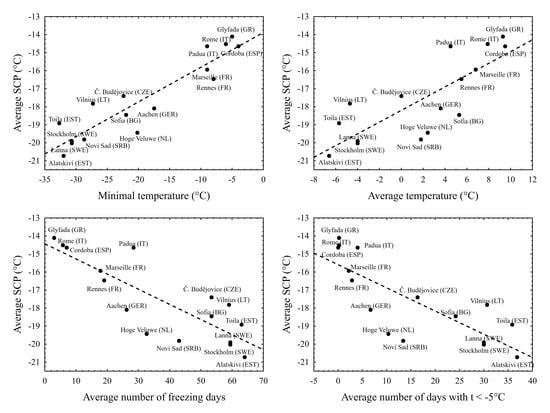
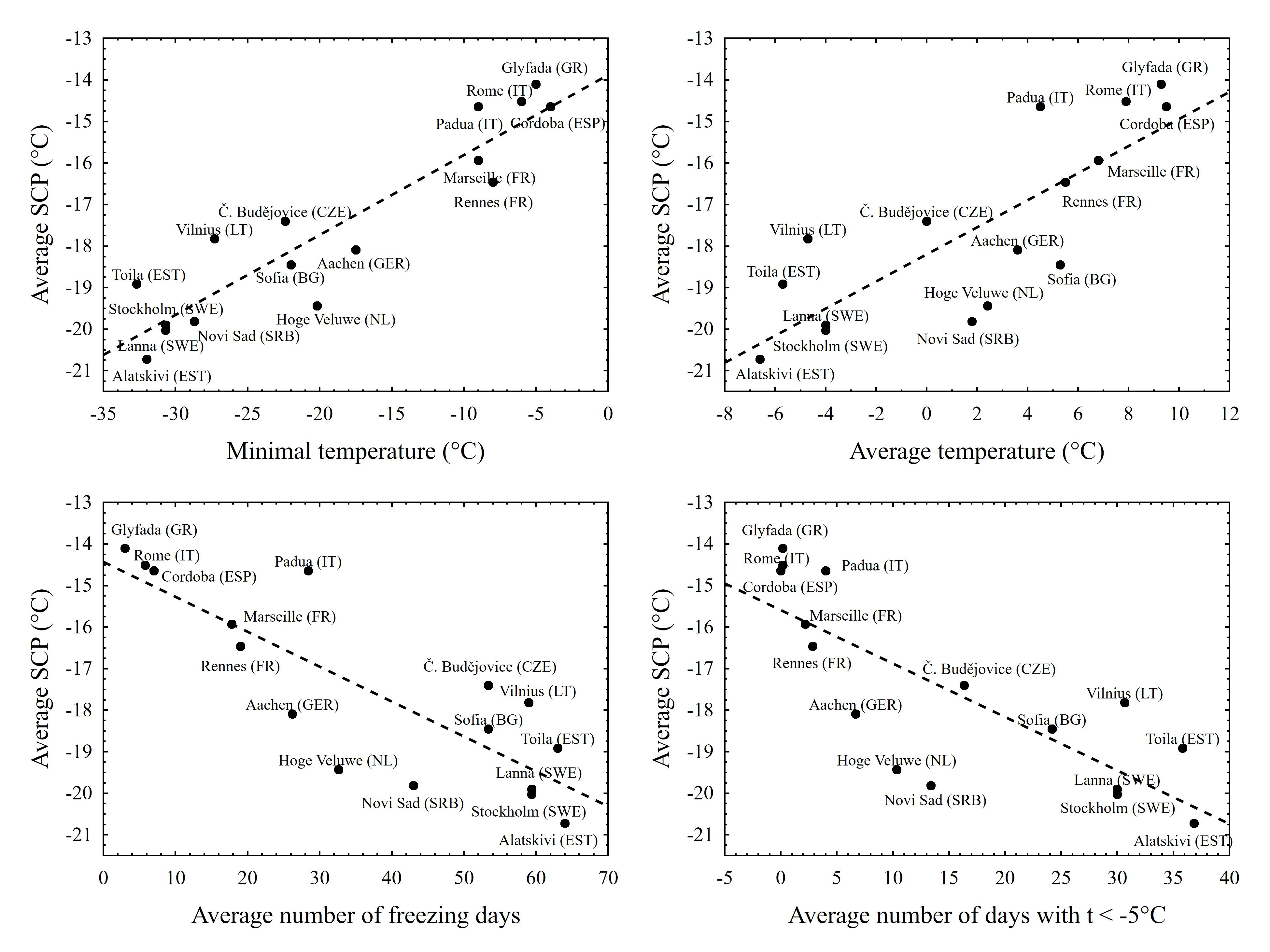
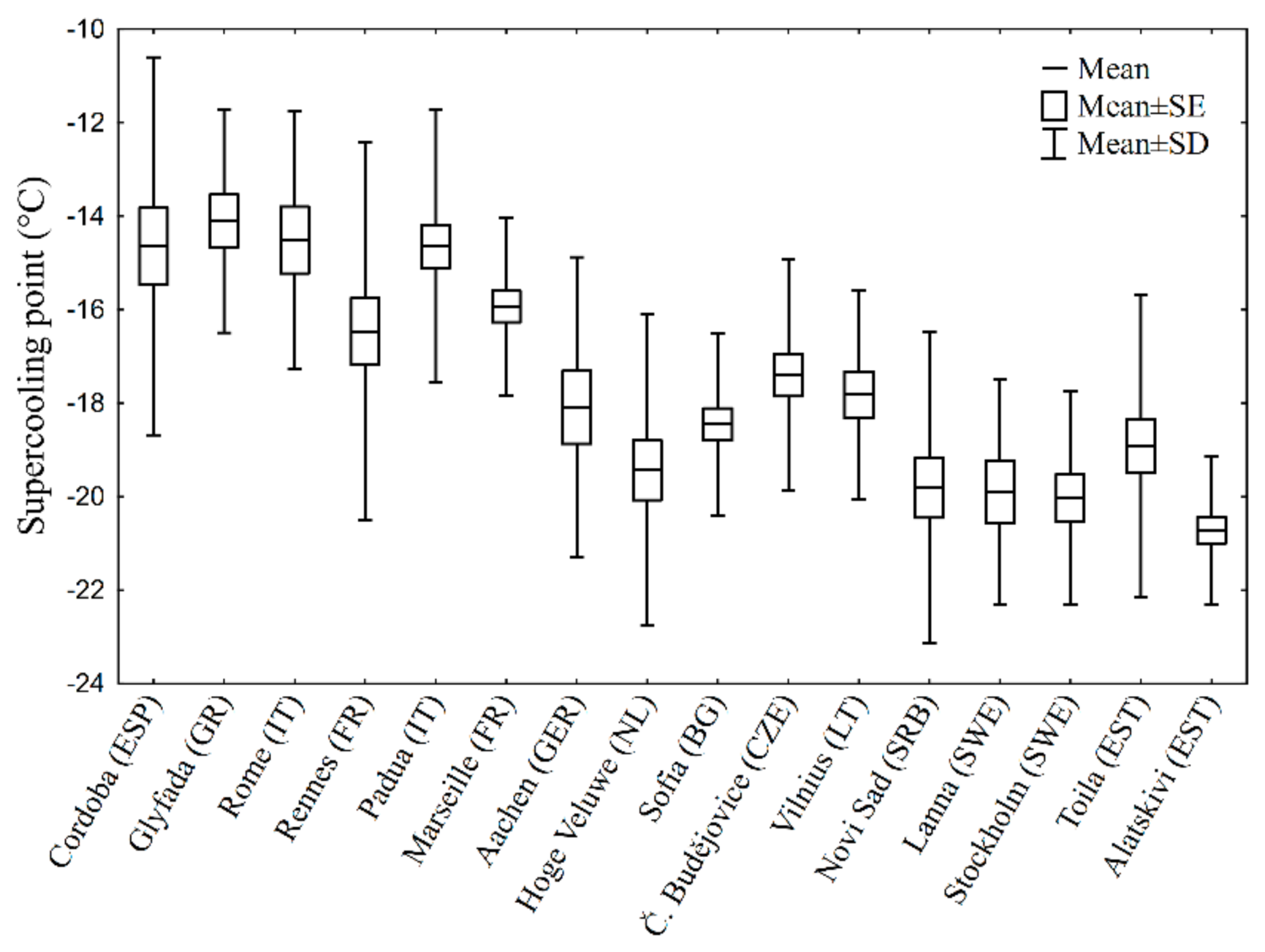
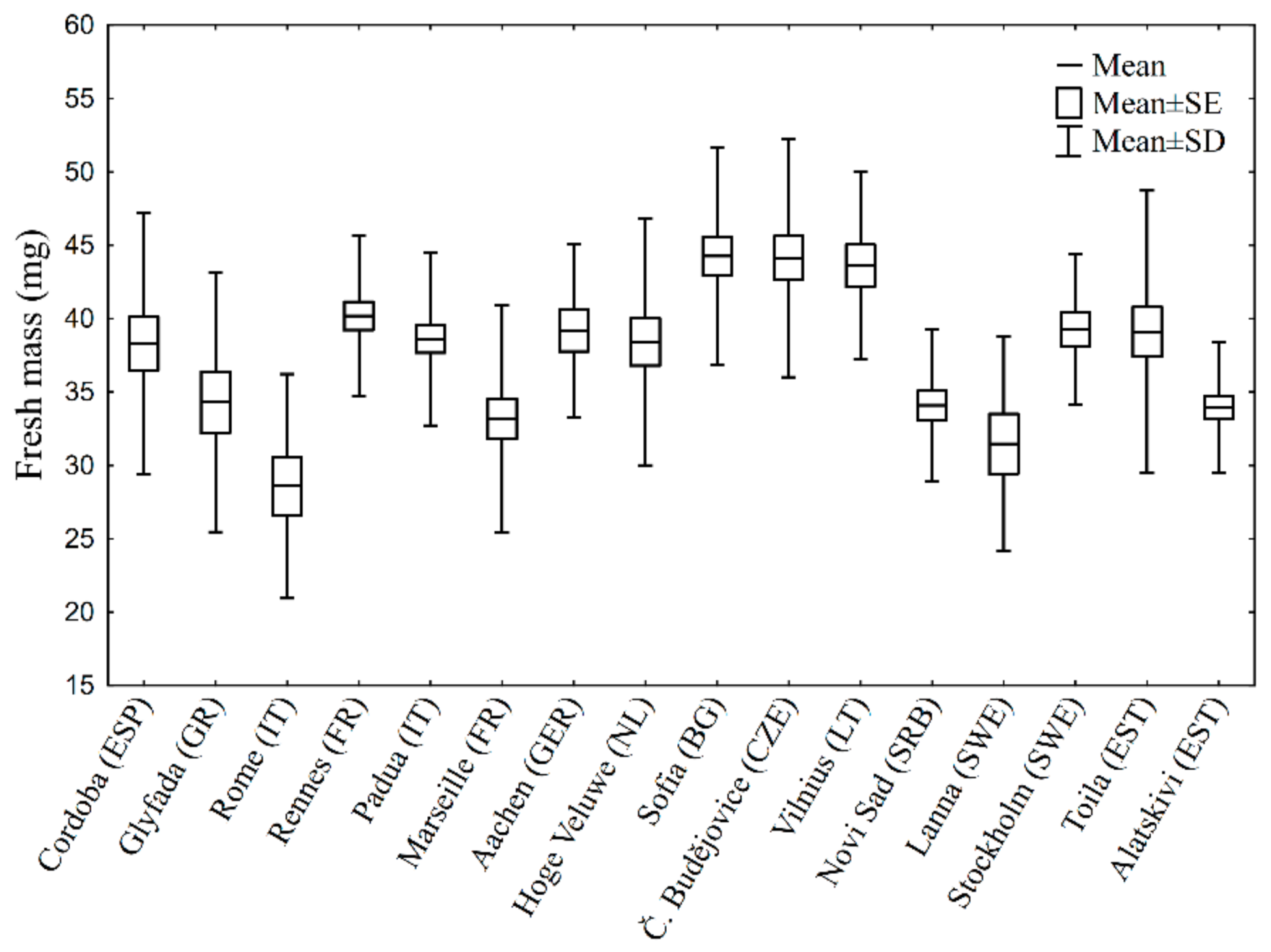
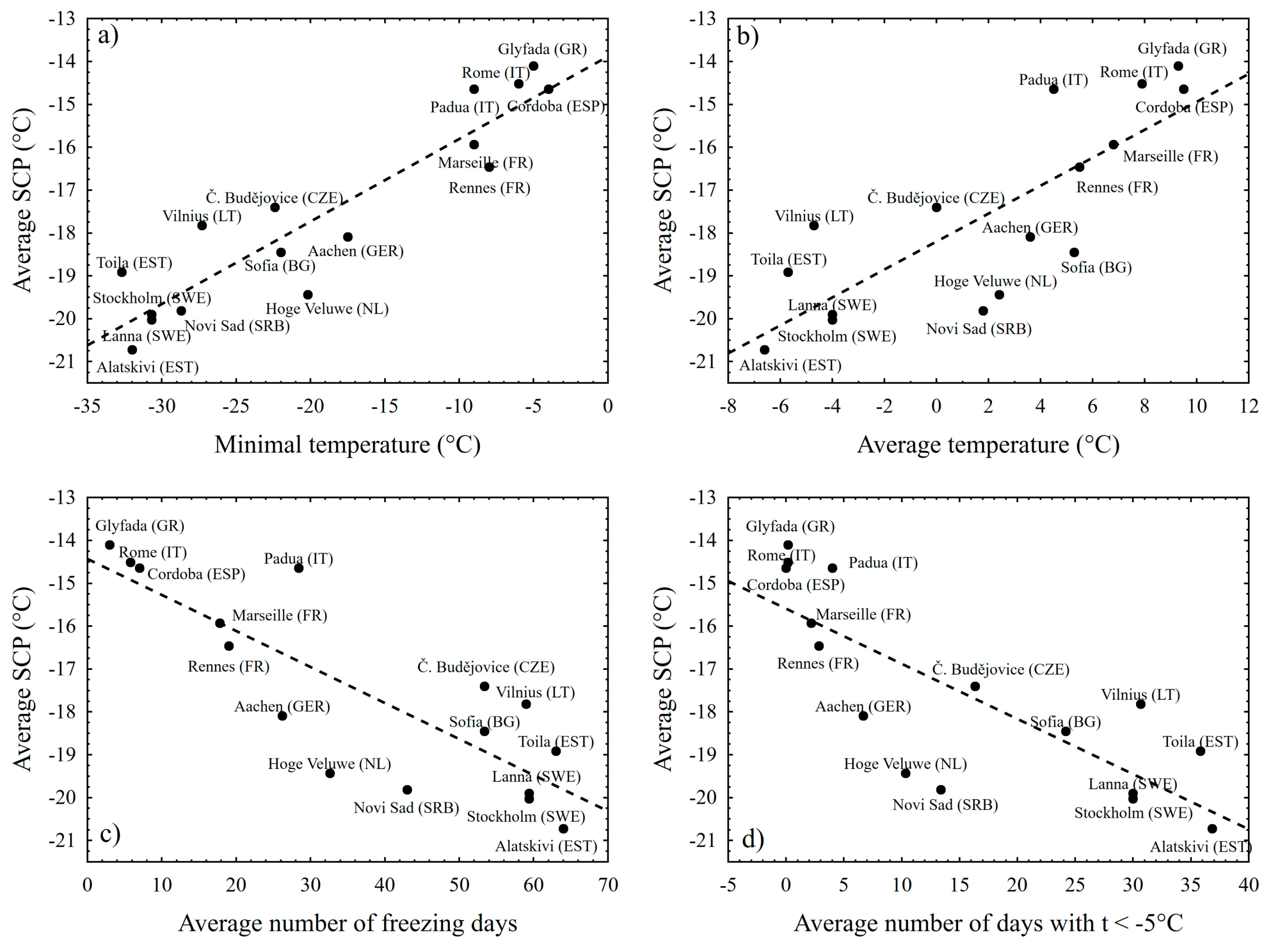
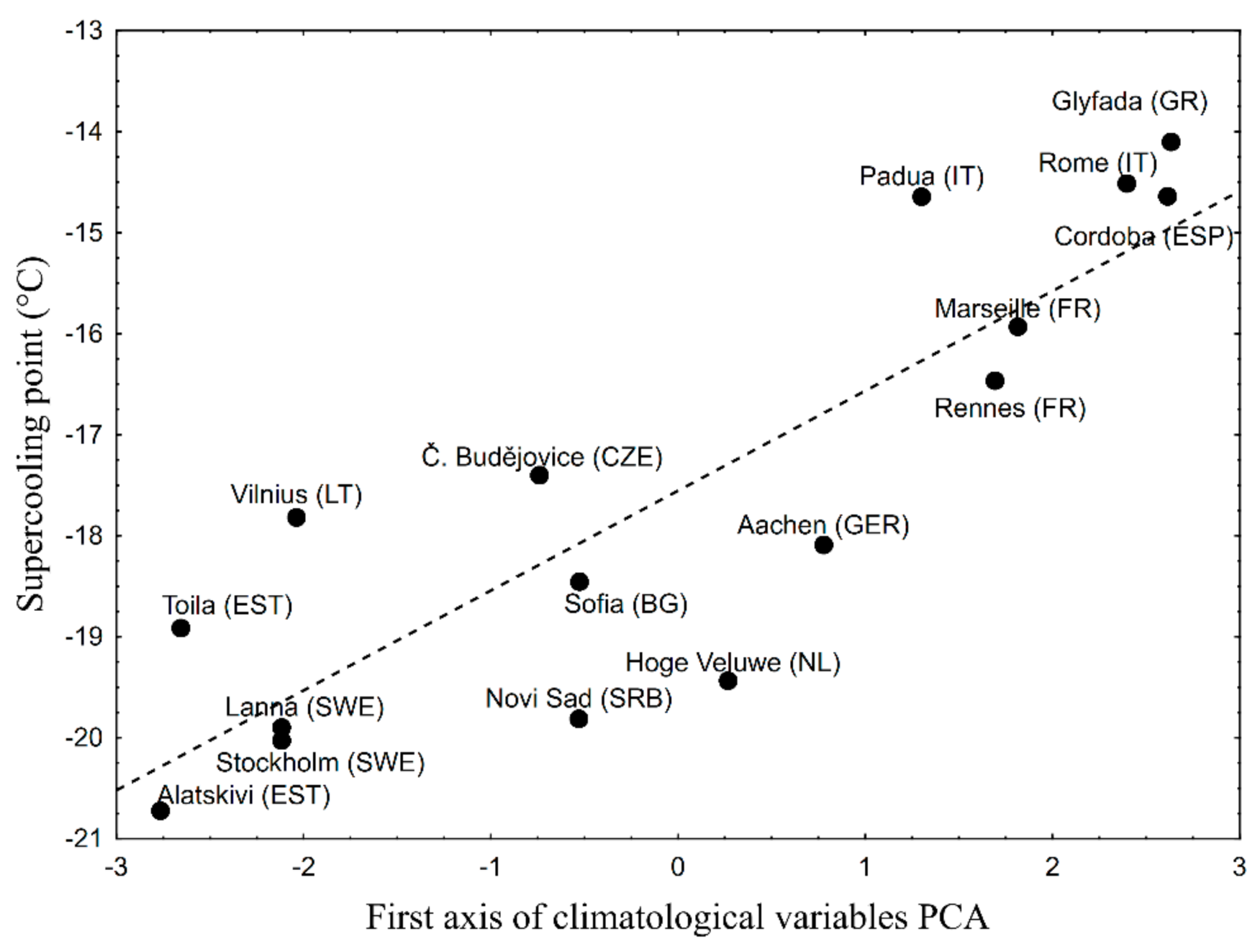
3. Results
4. Discussion
Limitation of this Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leather, S.R.; Walters, K.F.A.; Bale, J.S. The Ecology of Insect Overwintering; Cambridge University Press: Cambridge, UK, 1993; p. 255. [Google Scholar]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.S.; Worland, M.R. How insects survive the cold: Molecular mechanisms—A review. J. Comp. Physiol. B 2008, 178, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B.; Storey, J.M. Insect cold hardiness: Metabolic, gene, and protein adaptation. Can. J. Zool. 2012, 90, 456–475. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Lee, R.E. Low Temperature Biology of Insects; Cambridge University Press: Cambridge, UK, 2010; p. 390. [Google Scholar]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J.; Chown, S.L. Climatic variability and hemispheric differences in insect cold tolerance: Support from southern Africa. Funct. Ecol. 2005, 19, 214–221. [Google Scholar] [CrossRef]
- Ditrich, T. Supercooling point is an individually fixed metric of cold tolerance in Pyrrhocoris apterus. J. Therm. Biol. 2018, 74, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Košťál, V.; Šimek, P. Overwintering strategy in Pyrrhocoris apterus (Heteroptera): The relations between life-cycle, chill tolerance and physiological adjustments. J. Insect Physiol. 2000, 46, 1321–1329. [Google Scholar] [CrossRef]
- Nedvěd, O.; Hodková, M.; Brunnhofer, V.; Hodek, I. Simultaneous measurement of low-temperature survival and supercooling in a sample of insects. Cryoletters 1995, 16, 108–113. [Google Scholar]
- Hodková, M.; Hodek, I. Temperature regulation of supercooling and gut nucleation in relation to diapause of Pyrrhocoris apterus (Heteroptera). Cryobiology 1997, 34, 70–79. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.T. Cold and heat tolerance of drosophilid flies with reference to their latitudinal distributions. Oecologia 2004, 140, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Kellermann, V.; Loeschcke, V.; Hoffmann, A.A.; Kristensen, T.N.; Flojgaard, C.; David, J.R.; Svenning, J.C.; Overgaard, J. Phylogenetic constraints in key functional traits behind species’ climate niches: Patterns of desiccation and cold resistance across 95 drosophila species. Evolution 2012, 66, 3377–3389. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J.; Woods, A. How to assess Drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- David, J.R.; Gibert, P.; Moreteau, B.; Gilchrist, G.W.; Huey, R.B. The fly that came in from the cold: Geographic variation of recovery time from low-temperature exposure in Drosophila subobscura. Funct. Ecol. 2003, 17, 425–430. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Williams, C.M.; Terblanche, J.S. Variation in thermal performance among insect populations. Physiol. Biochem. Zool. 2012, 85, 594–606. [Google Scholar] [CrossRef] [PubMed]
- Crozier, L. Winter warming facilitates range expansion: Cold tolerance of the butterfly Atalopedes campestris. Oecologia 2003, 135, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Kalushkov, P.; Nedvěd, O. Cold hardiness of Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae) from central and southern europe. Eur. J. Entomol. 2000, 97, 149–153. [Google Scholar] [CrossRef]
- Xie, H.C.; Li, D.S.; Zhang, H.G.; Mason, C.E.; Wang, Z.Y.; Lu, X.; Cai, W.Z.; He, K.L. Seasonal and geographical variation in diapause and cold hardiness of the asian corn borer, Ostrinia furnacalis. Insect Sci. 2015, 22, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, L.E.; Lardies, M.A.; Bozinovic, F. Interpopulational variation in recovery time from chill coma along a geographic gradient: A study in the common woodlouse, Porcellio laevis. J. Insect Physiol. 2005, 51, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- Socha, R. Pyrrhocoris apterus (Heteroptera)—An experimental model species: A review. Eur. J. Entomol. 1993, 90, 241–286. [Google Scholar]
- Renault, D.; Salin, C.; Vannier, G.; Vernon, P. Survival at low temperatures in insects: What is the ecological significance of the supercooling point? Cryoletters 2002, 23, 217–228. [Google Scholar] [PubMed]
- Pivarciova, L.; Vaněčková, H.; Provaznik, J.; Wu, B.C.H.; Pivarci, M.; Pecková, O.; Bazalová, O.; Cada, S.; Kment, P.; Kotwica-Rolinska, J.; et al. Unexpected geographic variability of the free running period in the linden bug Pyrrhocoris apterus. J. Biol. Rhythm. 2016, 31, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Root, T.L.; Willig, M.R. Impacts of extreme weather and climate on terrestrial biota. Bull. Am. Meteorol. Soc. 2000, 81, 443–450. [Google Scholar] [CrossRef]
- Bale, J.S. Insect cold hardiness: A matter of life and death. Eur. J. Entomol. 1996, 93, 369–382. [Google Scholar]
- Numata, H.; Saulich, A.H.; Volkovich, T.A. Photoperiodic responses of the linden bug, Pyrrhocoris apterus, under conditions of constant-temperature and under thermoperiodic conditions. Zool. Sci. 1993, 10, 521–527. [Google Scholar]
- Saunders, D.S. A diapause induction termination asymmetry in the photoperiodic responses of the linden bug, Pyrrhocoris apterus and an effect of near-critical photoperiods on development. J. Insect Physiol. 1983, 29, 399–405. [Google Scholar] [CrossRef]
- Šlachta, M.; Vambera, J.; Zahradníčková, H.; Košťál, V. Entering diapause is a prerequisite for successful cold-acclimation in adult Graphosoma lineatum (Heteroptera: Pentatomidae). J. Insect Physiol. 2002, 48, 1031–1039. [Google Scholar] [CrossRef]
- Denlinger, D.L. Relationship between cold hardiness and diapause. In Insects at Low Temperature; Lee, R.E., Denlinger, D.L., Eds.; Springer: Boston, MA, USA, 1991; pp. 174–198. ISBN 978-1-4757-0190-6. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | GPS | Generation | Sampling Date | Climatic Data Source | n |
|---|---|---|---|---|---|
| Aachen (GER) | 50°49′56.8″ N | F7 | 06/2014 | The Royal Netherlands Meteorological Institute | 17 |
| 06°03′01.6″ E | |||||
| Alatskivi (EST) | 58°35′31.7″ N | F31 | 06/2010 | The Estonian Environment Agency | 32 |
| 27°07′29.8″ E | |||||
| Cordoba (ESP) | 37°53′21.5″ N | F17 | 08/2012 | Weather Underground | 24 |
| 04°47′51.7″ W | |||||
| Č. Budějovice (CZE) | 48°59′37.3″ N | F3 | 08/2014 | Czech Hydrometeorological Institute | 30 |
| 14°32′30.1″ E | |||||
| Glyfada (GR) | 37°52′46.1″ N | F33 | 08/2009 | Helenic National Meteorological Service | 18 |
| 23°46′06.2″ E | |||||
| Hoge Veluwe (NL) | 52°04′57.7″ N | F19 | 06/2012 | The Royal Netherlands Meteorological Institute | 27 |
| 05°50′00.2″ E | |||||
| Lanna (SWE) | 59°12′00.0″ N | F36 | 04/2010 | Weather Underground | 13 |
| 18°09′00.0″ E | |||||
| Marseille (FR) | 43°17′35.9″ N | F58 | 06/2004 | Weather Underground | 32 |
| 05°21′35.6″ E | |||||
| Novi Sad (SRB) | 45°15′41.4″ N | F30 | 06/2011 | Republic Hydrometeorological Service of Serbia | 27 |
| 19°51′19.4″ E | |||||
| Padua (IT) | 45°24′34.8″ N | F4 | 07/2016 | Weather Underground | 40 |
| 11°53′53.3″ E | |||||
| Rennes (FR) | 48°06′51.8″ N | F3 | 05/2016 | Weather Underground | 32 |
| 1°38′08.3″ W | |||||
| Rome (IT) | 41°55′52.8″ N | F4 | 06/2016 | Weather Underground | 15 |
| 12°28′54.4″ E | |||||
| Sofia (BG) | 42°42′00.0″ N | F27 | 07/2011 | Weather Underground | 32 |
| 23°19′12.0″ E | |||||
| Stockholm (SWE) | 59°12′00.0″ N | F36 | 04/2010 | Weather Underground | 20 |
| 18°09′00.0″ E | |||||
| Toila (EST) | 59°24′57.6″ N | F33 | 06/2010 | The Estonian Environment Agency | 32 |
| 27°31′02.3″ E | |||||
| Vilnius (LT) | 54°41′41.9″ N | F0 | 10/2015 | Lithuanian Hydrometeorological Service | 20 |
| 25°15′34.8″ E |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ditrich, T.; Janda, V.; Vaněčková, H.; Doležel, D. Climatic Variation of Supercooling Point in the Linden Bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Insects 2018, 9, 144. https://doi.org/10.3390/insects9040144
Ditrich T, Janda V, Vaněčková H, Doležel D. Climatic Variation of Supercooling Point in the Linden Bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Insects. 2018; 9(4):144. https://doi.org/10.3390/insects9040144
Chicago/Turabian StyleDitrich, Tomáš, Václav Janda, Hana Vaněčková, and David Doležel. 2018. "Climatic Variation of Supercooling Point in the Linden Bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae)" Insects 9, no. 4: 144. https://doi.org/10.3390/insects9040144
APA StyleDitrich, T., Janda, V., Vaněčková, H., & Doležel, D. (2018). Climatic Variation of Supercooling Point in the Linden Bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Insects, 9(4), 144. https://doi.org/10.3390/insects9040144