Lethal and Sublethal Effects of Three Microbial Biocontrol Agents on Spodoptera litura and Its Natural Predator Rhynocoris kumarii

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Rearing of Insects
2.2. Microbial Biocontrol Agents and Bioassay for Assessment
2.3. Determining the Feeding Deterrence Index
2.4. Food Consumption
2.5. Rearing of Larvae with Entomopathogenic Microbes
2.6. Effect of Entomopathogenic Microbes on the Developmental Biology of the Predator
2.7. Statistical Analysis
3. Results
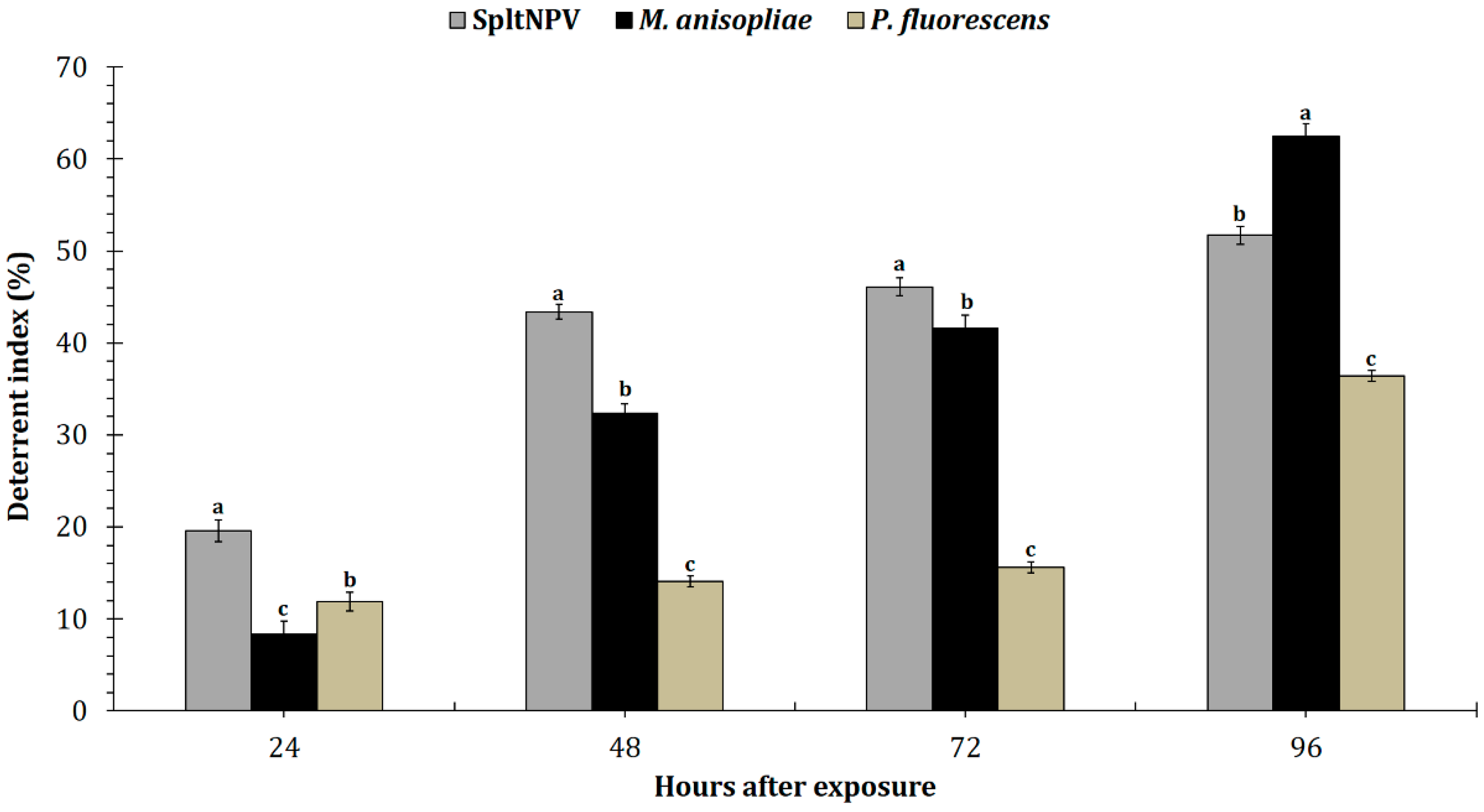
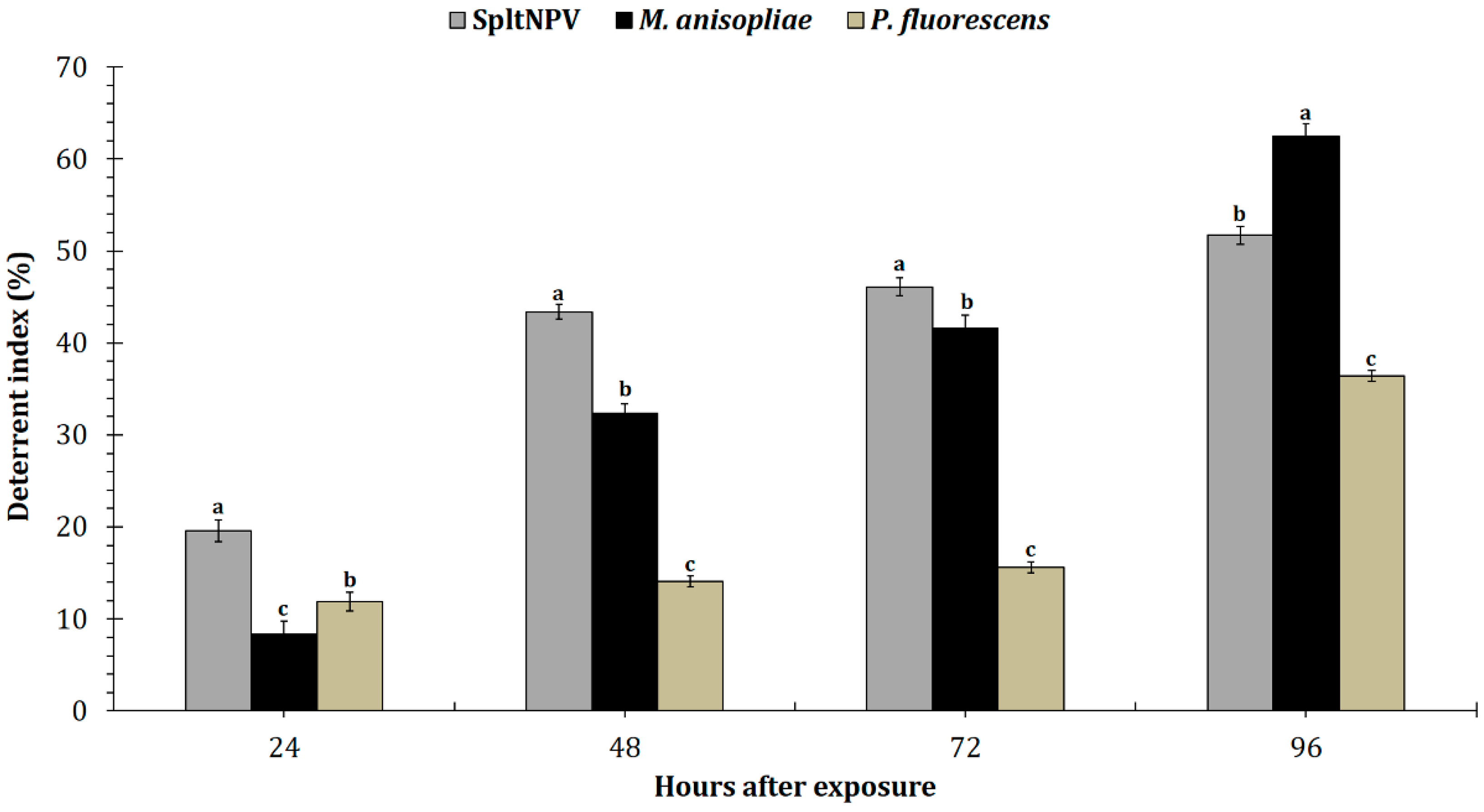
3.1. Food Deterrence Index
3.2. Food Energy Budget
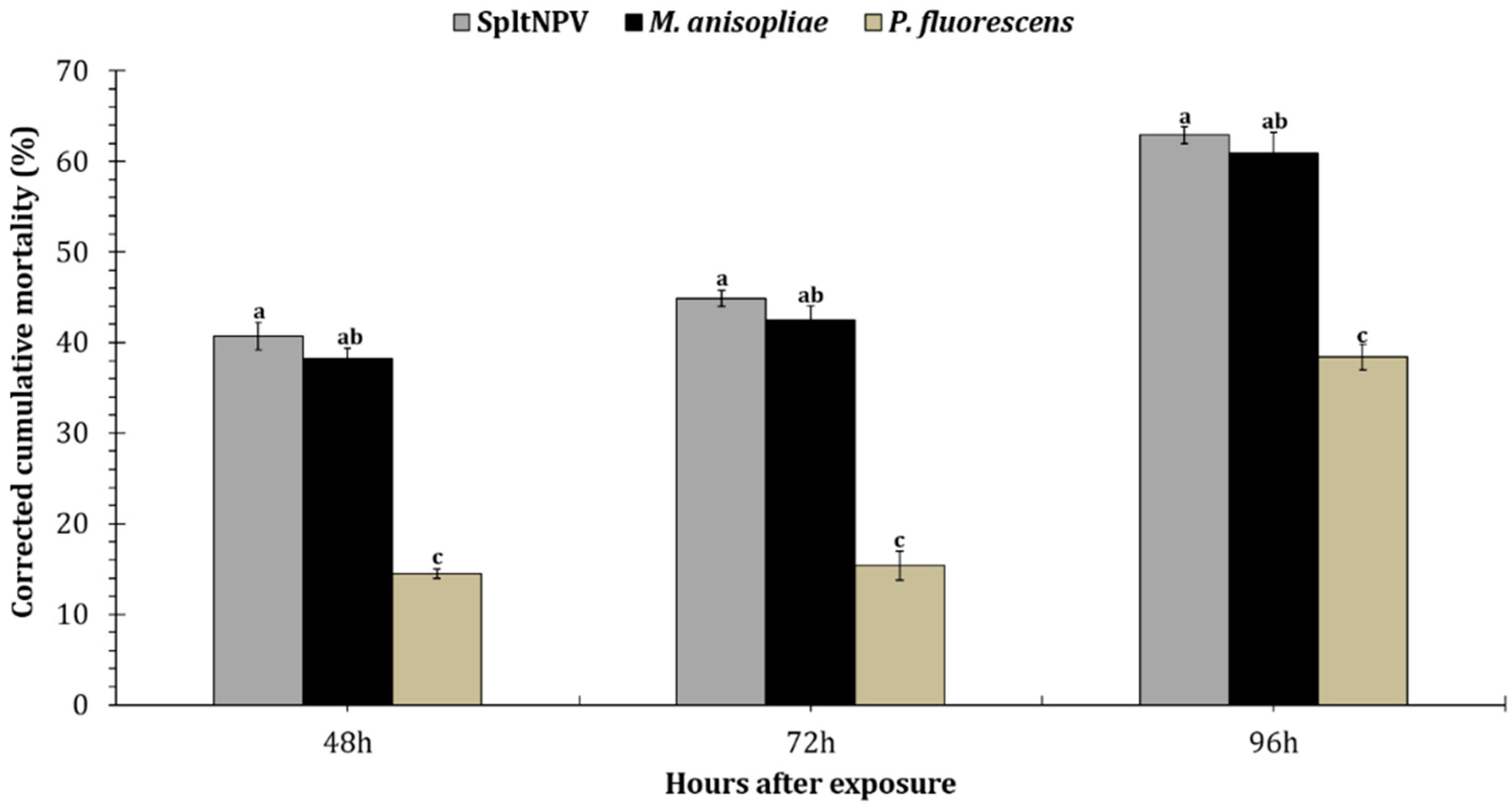
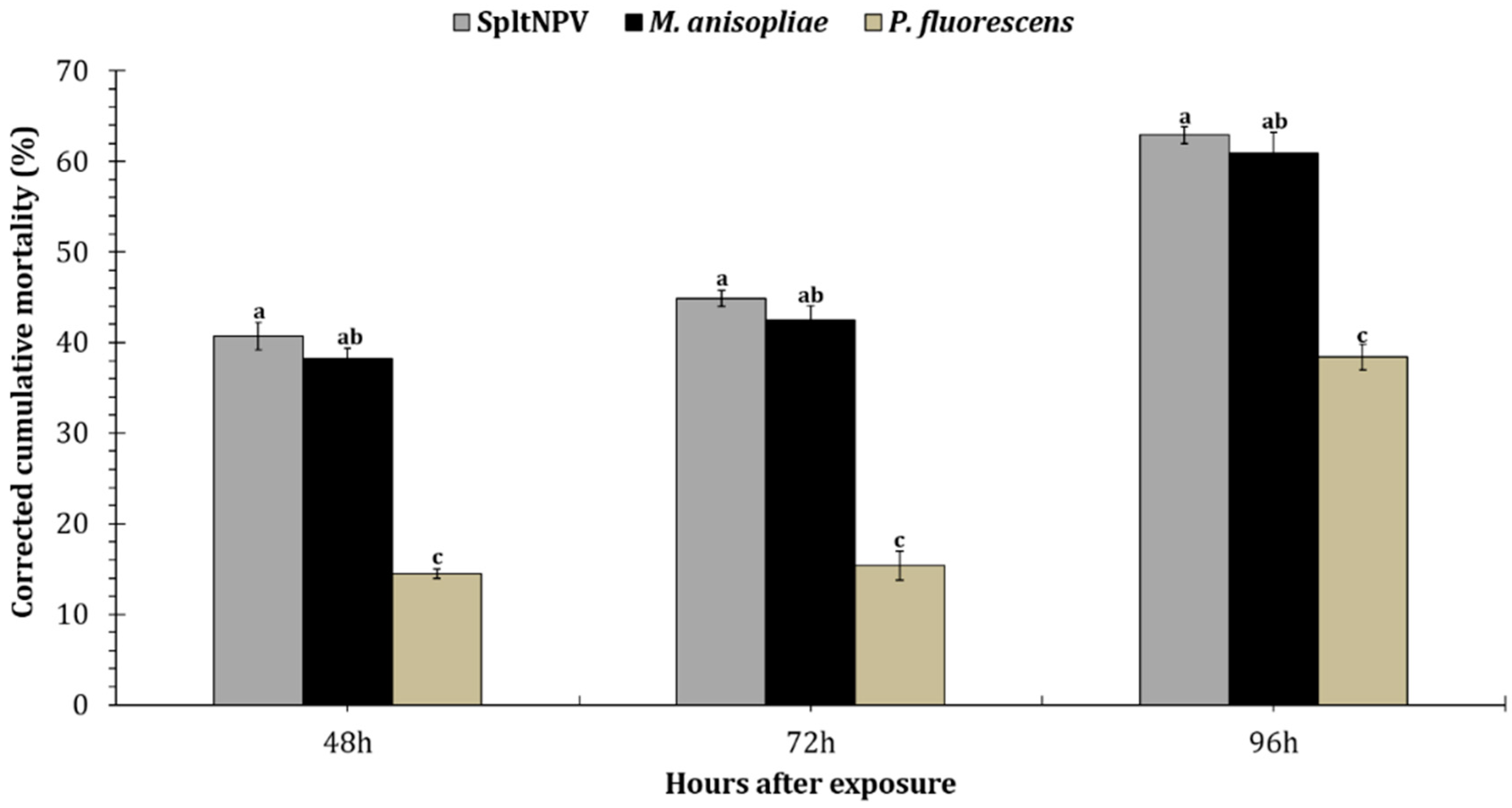
3.3. Larvicidal Activity of Entomopathogenic Microbes
3.4. Predation Behavior and Bioefficacy of R. kumarii
3.5. Predator Biology
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Noma, T.; Colunga-Garcia, M.; Brewer, M.; Landis, J.; Gooch, A. Oriental leafworm, Spodoptera litura. Michigan State University’s invasive species fact sheets. Feburary 2010. Available online: www.canr.msu.edu/ipm/uploads/files/Forecasting_invasion_risks/orientalLeafworm.pdf (accessed on 18 July 2018).
- Punithavalli, M.; Sharma, A.N.; Balaji, RM. Seasonality of the common cutworm Spodoptera litura in a soybean ecosystem. Phytoparasitica 2014, 42, 213–222. [Google Scholar] [CrossRef]
- Rao, M.S.; Manimanjari, D.; Rao, A.C.R.; Swathi, P.; Maheswari, M. Effect of climate change on Spodoptera litura Fab. on peanut: A life table approach. Crop Prot. 2014, 66, 98–106. [Google Scholar]
- Kranthi, K.R.; Jadhav, D.R.; Kranthi, S.; Wanjari, R.R.; Ali, R.R.; Russell, D.A. Insecticide resistance in five major insect pests of cotton in India. Crop Prot. 2002, 21, 449–460. [Google Scholar] [CrossRef]
- Gandhi, K.; Patil, R.H.; Srujana, Y. Field resistance of Spodoptera litura (Fab.) to conventional insecticides in India. Crop Prot. 2016, 88, 103–108. [Google Scholar]
- Abbas, N.; Shad, A.A.; Razaq, M. Fitness cost, cross resistance and realized heritability of resistance to imidacloprid in Spodoptera litura (Lepidoptera: Noctuidae). Pest. Biochem. Physiol. 2012, 103, 181–188. [Google Scholar] [CrossRef]
- Shad, S.; Sayyed, A.; Fazal, S.; Saleem, M.; Zaka, S.; Ali, M. Field evolved resistance to carbamates, organophosphates, pyrethroids, and new chemistry insecticides in Spodoptera litura Fab. (Lepidoptera: Noctuidae). J. Pest. Sci. 2012, 85, 153–162. [Google Scholar] [CrossRef]
- Yang, M.M.; Meng, L.L.; Yongan, Z.; Yu, Z.W.; Liang, J.Q.; Qing, H.W.; Jin, Y.D. Baculoviruses and insect pests control in China. Afr. J. Microbiol. Res. 2012, 6, 214–218. [Google Scholar] [CrossRef]
- Radja Commare, C.; Nandakumar, R.; Kandan, A.; Suresh, S.; Bharathi, M.; Raguchander, T.; Samiyappan, R. Pseudomonas fluorescens based bio-formulation for the management of sheath blight disease and leaf folder insect in rice. Crop Prot. 2002, 21, 671–677. [Google Scholar] [CrossRef]
- Hu, Q.B.; Ren, S.X.; An, X.C.; Qian, M.H. Insecticidal activity influence of destruxins on the pathogenicity of Paecilomyces javanicus against Spodoptera litura. J. Appl. Entomol. 2007, 131, 262–268. [Google Scholar] [CrossRef]
- Bhanu Prakash, G.V.; Padmaja, V.; Jami, S.K.; Kirti, P.B. Expression of chitinase genes of Metarhizium anisopliae isolates in lepidopteran pests and on synthetic media. J. Basic Microbiol. 2012, 52, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Mohana, S.A.; Dhandapani, N.; Shiv Sankar, H.D.; Muthukumar, P. Effect of gibberellic acid (GA3) and Pseudomonas fluorescens (migula) on the ovipositional behaviour of Plutella xylostella (L.) and Spodoptera litura (Fab.). Bull. Agric. Sci. 2006, 4, 55–58. [Google Scholar]
- Pechy-Tarr, M.; Bruck, D.J.; Maurhofer, M.; Fischer, E.; Vogne, C.; Henkels, M.D.; Donahue, K.M.; Grunder, J.; Loper, J.E.; Keel, C. Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ. Microbiol. 2008, 10, 2368–2386. [Google Scholar]
- Mohana, S.A.; Dhandapani, N. Gibberellic acid (GA3) and Pseudomonas fluorescens as components of biointensive pest management strategy against Plutella xylostella in cauliflower. Pest Manag. Hort. Ecosyst. 2009, 15, 1–8. [Google Scholar]
- Kumari, V.; Singh, N.P. Spodoptera litura nuclear polyhedrosis virus (NPV-S) as a component in integrated pest management (IPM) of Spodoptera litura (Fab.) on cabbage. J. Biopest. 2009, 2, 84–86. [Google Scholar]
- Nakai, M.; Nguyen, T.T.C. Field application of an insect virus in the Mekong Delta: Effects of a Vietnamese nucleopolyhedrovirus on Spodoptera litura (Lepidoptera: Noctuidae) and its parasitic natural enemies. Biocontrol Sci. Technol. 2005, 15, 443–453. [Google Scholar] [CrossRef]
- Singha, D.; Singha, B.; Dutta, B.K. Potential of Metarhizium anisopliae and Beauveria bassiana in the control of tea termite Microtermes obesi Holmgren in vitro and under field conditions. J. Pest Sci. 2011, 84, 69–75. [Google Scholar] [CrossRef]
- Fadare, T.A.; Amusa, N.A. Comparative efficacy of microbial and chosen insecticides on four major lepidopterous pests of cotton and their natural enemies. African J. Biotechnol. 2003, 2, 425–428. [Google Scholar]
- Gupta, R.K.; Amin, M.; Bali, K.; Monobrullah, M.; Jasrotia, P. Vertical transmission of sublethal granulovirus infection in the tobacco caterpillar Spodoptera litura. Phytoparasitica 2010, 38, 209–216. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Borgio, J.F. Virulence evaluation of entomopathogenic fungus Metarhizium anisopliae (Metsch.) Sorokin (Hyphomycetes) on seven insect pests. Indian J. Agric. Sci. 2010, 44, 195–200. [Google Scholar]
- Quesada-Moraga, E.; Carrasco-Díaz, J.A.; Santiago-Álvarez, C. Insecticidal and antifeedant activities of proteins secreted by entomopathogenic fungi against Spodoptera littoralis (Lepidoptera: Noctuiidae). J. Appl. Entomol. 2006, 130, 442–452. [Google Scholar] [CrossRef]
- Shapiro, M.; Robertson, J.L.; Webb, R.E. Effect of neem seed extract upon the gypsy moth (Lepidoptera: Lymantriidae) and its nuclear polyhedrosis virus. J. Econ. Entomol. 1994, 87, 356–360. [Google Scholar] [CrossRef]
- Jun, T.; Shohei, O.; Takayoshi, I.; Madoka, N.; Yasuhisa, K. Productivity and quality of polyhedral occlusion bodies of a nucleopolyhedrovirus harvested from Spodoptera litura (Lepidoptera: Noctuidae) larvae. Appl. Entomol. Zool. 2007, 42, 21–26. [Google Scholar]
- Rao, G.V.R.; Kumar, C.S.; Sireesha, K.; Kumar, P.L. Role of nucleopolyhedroviruses (NPVs) in the management of lepidopteran pests in Asia. In Biocontrol of Lepidopteran Pests, Soil Biology; Sree, K.S., Varma, A., Eds.; Springer International Publishing: Berlin, Germany, 2015; pp. 11–52. [Google Scholar]
- Petlamul, W.; Prasertsan, P. Evaluation of strains of Metarhizium anisopliae and Beauveria bassiana against Spodoptera litura on the basis of their virulence, germination rate, conidia production, radial growth and enzyme activity. Mycobiology 2012, 40, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Sowjanya Sree, K.; Padmaja, V.; Murthy, Y.L. Insecticidal activity of destruxin, a mycotoxin from Metarhizium anisopliae (Hypocreales), against Spodoptera litura (Lepidoptera: Noctuidae) larval stages. Pest Manag. Sci. 2008, 64, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan, R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pest and diseases. Crop Prot. 2001, 20, 1–11. [Google Scholar] [CrossRef]
- Roobakkumar, A.; Babu, A.; Vasantha Kumar, D.; Rahman, V.K.; Sarkar, S. Pseudomonas fluorescens as an efficient entomopathogen against Oligonychus coffeae Nietner (Acari: Tetranychidae) infesting tea. J. Entomol. Nematol. 2011, 3, 73–77. [Google Scholar]
- Al-Sohim, A.S.; Fouly, A.F. Biological effects of two bacterial isolates and mutants of Pseudomonas fluorescens on date palm red spider mite, Oligonychus afrasiaticus (Acari: Tertanychidae). Egyptian J. Biol. Pest Control 2015, 25, 513–518. [Google Scholar]
- Sahayaraj, K. Reduviids and their merits in biological control. In Basic and Applied Aspects of Biopesticides; Springer: New Delhi, India, 2014; pp. 195–214. [Google Scholar]
- Ambrose, D.P. Assassin Bugs; USA Science Publishers Inc.: Enfield, NH, USA, 1999; p. 337. [Google Scholar]
- Grundy, P.; Maelzer, D. Assessment of Pristhesancus plagipennis (Walker) (Hemiptera: Reduviidae) as an augmented biological control in cotton and soybean crops. Aust. J. Entomol. 2000, 39, 305–309. [Google Scholar] [CrossRef]
- Sahayaraj, K. Pest Control Mechanism of Reduviids; ABD Publisher: Jaipur, India, 2007; p. 240. [Google Scholar]
- Sahayaraj, K. Hunter reduviidae in pest management for plantation crops. Proceeding of National Seminar on Harmful/Beneficial Insects of Agricultural Importance, Calicut, India, 17–18 February 2011; pp. 42–51. [Google Scholar]
- Sahayaraj, K.; Kalidas, S.; Majesh, T. Stage preference and functional response of Rhynocoris longifrons (Stål) (Hemiptera: Reduviidae) on three hemipteran cotton pests. Braz. Arch. Biol. Technol. 2012, 55, 733–740. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Asha, A. Biological control potential evaluation of Rhynocoris kumarii Ambrose and Livingstone (Hemiptera: Reduviidae) on Aphis craccivora (Koch.) (Hemiptera: Aphididae). Indian J. Agric. Res. 2010, 44, 281–287. [Google Scholar]
- Claver, M.A.; Ambrose, D.P. Evaluation of Rhynocoris kumarii Ambrose & Livingstone (Hemiptera: Reduviidae) as a potential predator of some lepidopteran pests of cotton. J. Biol. Control 2001, 15, 15–20. [Google Scholar]
- Ambrose, D.P. Suppression of Helicoverpa armigera (Hubner) and Anomis flava (Fab.) infesting okra by the reduviid predator Rhynocoris kumarii Ambrose and Livingstone in field cages. Pest Manag. Hort. Ecosyst. 2000, 6, 32–35. [Google Scholar]
- Claver, M.A.; Ambrose, D.P. Impact of augmentative release of Rhynocoris kumarii Ambrose & Livingstone (Heteroptera: Reduviidae) on Dysdercus cingulatus (Fabricius) (Hemiptera: Pyrrhocoridae) population and damage on cotton. J. Biol. Control 2001, 15, 119–125. [Google Scholar]
- Claver, M.A.; Kalyanasundaram, M.; David, P.M.M.; Ambrose, D.P. Abundance of boll worm, flower beetle, predators and field colonization by Rhynocoris kumarii (Heteroptera: Reduvidae) following mulching and shelter provisioning in cotton. J. Appl. Entomol. 2003, 127, 383–388. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Ravi, C. Evaluation of reduviid predators and plant products against chosen groundnut pests. Arch. Phytopathol. Plant Prot. 2007, 40, 281–290. [Google Scholar] [CrossRef]
- Cooper, D.J. The role of predatory hemiptera in disseminating a nuclear polyhedrosis virus of Heliothis punctiger. Aust. J. Entomol. 1981, 20, 145–150. [Google Scholar] [CrossRef]
- Thungrabeab, M.; Tongma, S. Effect of entomopathogenic fungi, Beauveria bassiana (Balsam) and Metarhizium anisopliae (Metsch.) on non target insects. KMITL Sci. Technol. J. 2007, 7, 8–12. [Google Scholar]
- Sajap, A.S.; Kotuli, J.R.; Kadir, H.A.; Hussein, M.Y. Impact of prey infected by nuclear polyhedrosis virus on a predator, Sycanus leucomesus Walk. (Hemiptera: Reduviidae). J. Appl. Entomol. 1999, 123, 93–97. [Google Scholar] [CrossRef]
- Abbas, M.S.T.; Boucias, D.G. Interaction between nuclear polyhedrosis virus-infected Anticarsia gemmatalis (Lepidoptera: Noctuidae) larvae and a predator Podisus maculiventris (Say) (Hemiptera: Pentatomidae). Environ. Entomol. 1984, 13, 599–602. [Google Scholar] [CrossRef]
- George, P.J.E.; Ambrose, D.P. Insecticidal impact on the post-embryonic development of Rhynocoris kumarii Ambrose and Livingstone (Heteroptera: Reduviidae). J. Appl. Entomol. 1999, 123, 509–512. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Karthikraja, S. Effect of biopesticides on Rhynocoris marginatus (Fab.) (Heteroptera: Reduvidae). J. Biol. Control 2003, 17, 43–45. [Google Scholar]
- Sahayaraj, K.; Abitha, J.M.; Selvaraj, P. Side effects of selected biopesticides on reduviid predator Rhynocoris marginatus Fab. Entomol. Croat. 2003, 7, 43–50. [Google Scholar]
- Sahayaraj, K.; Lalitha, C.; Balasubramaniam, R. Biosafety of Metarhizium anisopliae (Metschnikoff) Sorokin on a reduviid predator Acanthaspis pedestris (Hemiptera: Reduviidae). Hexapoda 2008, 15, 46–48. [Google Scholar]
- Isman, M.B.; Koul, O.; Luczynski, A.; Kaminski, J. Insecticidal and antifeedant bioactivities of neem oils and their relationship to azadirachtin content. J. Agric. Food Chem. 1990, 38, 1406–1411. [Google Scholar] [CrossRef]
- Waldbauer, G.P. The consumption and utilization of food by insects. Adv. Insect Physiol. 1968, 5, 229–287. [Google Scholar]
- Zhong, W.; Ding, S.; Guo, H. The chitinase C gene PsChiC from Pseudomonas sp. and its synergistic effects on larvicidal activity. Gen. Mol. Biol. 2015, 38, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Sun, X. History and current status of development and use of viral insecticides in China. Viruses 2015, 7, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.E.; Nottingham, S.F.; Stephens, A.E. Food and water economy and its relation to growth in fifth-instar larvae of tobacco hornworm, Manduca sexta. J. Insect Physiol. 1985, 31, 119–127. [Google Scholar] [CrossRef]
- Inceoglu, A.B.; Kamita, S.G.; Hammock, B.D. Genetically modified baculoviruses: A historical overview and future outlook. Adv. Virus Res. 2006, 68, 323–360. [Google Scholar] [PubMed]
- Rath, S.S.; Prasad, B.C.; Sinha, B.R. Food utilization efficiency in fifth instar larvae of Antheraea mylitta (Lepidoptera: Saturniidae) infected with Nosema sp. and its effect on reproductive potential and silk production. J. Invertebr. Pathol. 2003, 83, 1–9. [Google Scholar] [CrossRef]
- Ruberson, J.R.; Young, S.Y.; Kring, T.J. Suitability of prey infected by nuclear polyhedrosis virus for development, survival, and reproduction of the predator Nabis roseipennis (Heteroptera: Nabidae). Environ. Entomol. 1991, 20, 1475–1479. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gani, M.; Jasrotia, P.; Srivastava, K. Development of the predator Eocanthecona furcellata on different proportions of nucleopolyhedrovirus infected Spodoptera litura larvae and potential for predator dissemination of virus in the field. BioControl 2013, 58, 543–552. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatments | FCI | RGR | AD | ECIF | ECDF |
|---|---|---|---|---|---|
| SpltNPV | 28.2 ± 2.7d | 11.1 ± 1.5d | 52.1 ± 2.0d | 42.9 ± 1.2d | 40.1 ± 2.5d |
| M. anisopliae | 32.9 ± 4.6c | 14.0 ± 0.3c | 57.2 ± 0.9c | 59.2 ± 2.4c | 51.2 ± 1.0c |
| P. fluorescens | 66.0 ± 3.4b | 17.9 ± 0.1b | 83.9 ± 3.8b | 65.7 ± 1.5b | 73.2 ± 2.8b |
| Control | 81.2 ± 4.8a | 20.4 ± 0.2a | 110.0 ± 5.0a | 73.8 ± 0.8a | 93.1 ± 3.5a |
| Exposure Period (h) | Treatments | R. kumarii Fourth Instar | R. kumarii Fifth Instar | R. kumarii Adult | |||
|---|---|---|---|---|---|---|---|
| HT | PR | HT | PR | HT | PR | ||
| 24 | Control | 170.8a | 1.2 | 82.0b | 1.0 | 61.8c | 1.0 |
| SpltNPV | 75.2c | 1.2 | 109.8a | 1.4 | 116.0a | 1.0 | |
| M. anisopliae | 79.3c | 1.2 | 71.4c | 1.4 | 94.6b | 1.0 | |
| P. fluorescens | 113.6b | 1.9 | 61.4d | 1.4 | 63.2c | 1.0 | |
| 48 | Control | 177.6a | 1.4 | 141.9c | 1.0 | 131.6c | 1.2 |
| SpltNPV | 163.4b | 1.0 | 188.2b | 2.2 | 208.6a | 1.2 | |
| M. anisopliae | 94.8c | 1.2 | 104.2d | 1.4 | 78.4d | 1.2 | |
| P. fluorescens | 57.8d | 1.2 | 228.5a | 2.8 | 207.8ab | 1.9 | |
| 72 | Control | 188.6c | 1.0 | 166.6c | 1.2 | 120.8b | 1.2 |
| SpltNPV | 134.3d | 1.2 | 172.4b | 1.4 | 115.2bc | 1.6 | |
| M. anisopliae | 219.4a | 1.4 | 196.8a | 1.8 | 142.2a | 1.4 | |
| P. fluorescens | 215.2b | 1.4 | 113.2d | 1.6 | 98.7d | 1.2 | |
| 96 | Control | 169.8d | 1.4 | 154.8b | 1.6 | 166.6c | 2.0 |
| SpltNPV | 199.2a | 1.0 | 176.2a | 1.6 | 229.5a | 2.9 | |
| M. anisopliae | 189.3b | 1.4 | 146.6c | 1.6 | 130.5d | 1.6 | |
| P. fluorescens | 177.2c | 1.6 | 125.4d | 1.6 | 221.8b | 2.9 | |
| R. kumarii Life Stages | Treatments | Food Consumption (mg) Hours after Starvation | |||
|---|---|---|---|---|---|
| 24 | 48 | 72 | 96 | ||
| Fourth instar | Control | 43.6 ± 2.7a | 46.2 ± 1.0a | 39.2 ± 3.4a | 39.2 ± 2.0b |
| SpltNPV | 8.2 ± 0.8c | 23.2 ± 0.7b | 23.3 ± 0.6d | 38.8 ± 1.2bc | |
| M. anisopliae | 14.1 ± 1.2b | 23.8 ± 2.6b | 38.4 ± 0.6ab | 23.6 ± 2.6d | |
| P. fluorescens | 5.7 ± 0.7d | 17.2 ± 1.3c | 29.2 ± 1.5c | 54.2 ± 0.6a | |
| Fifth instar | Control | 11.0 ± 1.6d | 15.2 ± 1.5d | 14.7 ± 1.0d | 10.8 ± 0.7d |
| SpltNPV | 21.3 ± 1.5bc | 36.3 ± 1.8ab | 22.6 ± 0.9b | 22.2 ± 0.6b | |
| M. anisopliae | 28.6 ± 2.0a | 31.6 ± 0.8c | 28.3 ± 2.0a | 29.8 ± 2.0a | |
| P. fluorescens | 22.5 ± 2.0b | 37.3 ± 0.9a | 18.5 ± 0.9c | 19.6 ± 1.5c | |
| Adults | Control | 10.2 ± 0.5bc | 14.8 ± 1.0c | 21.6 ± 3.4bc | 42.0 ± 0.2b |
| SpltNPV | 7.8 ± 0.3d | 29.2 ± 1.0b | 21.6 ± 1.0b | 53.6 ± 3.2a | |
| M. anisopliae | 11.9 ± 0.6a | 32.2 ± 2.1a | 40.8 ± 2.7a | 36.2 ± 1.5c | |
| P. fluorescens | 11.6 ± 1.2ab | 29.2 ± 1.6b | 20.8 ± 1.0bd | 30.8 ± 1.7d | |
| Treatments | R. kumarii Life Stages or Category | |||||
|---|---|---|---|---|---|---|
| Third Instar | Fourth Instar | Male | Female | Total Nymphal Development Period (Days) * | Sex Ratio (Female:Male) | |
| Control | 11.3 ± 0.8d (100) * | 11.3 ± 0.3d (100) | 26.0 ± 0.3a (100) | 28.0 ± 0.3a (100) | 60.0 ± 1.3d | 0.47 |
| SpltNPV | 14.0 ± 0.7a (100) | 15.0 ± 0.5bc (80.67) | 21.1 ± 0.2b (96.77) | 25.2 ± 0.3b (91.66) | 66.9 ± 2.3b | 0.45 |
| M. anisopliae | 13.0 ± 0.3bc (100) | 16.2 ± 0.8ab (100) | 16.4 ± 0.5d (90.23) | 19.0 ± 0.9d (80.89) | 64.6 ± 1.3c | 0.46 |
| P. fluorescens | 13.2 ± 0.2b (100) | 16.4 ± 0.2a (100) | 17.3 ± 0.6c (100) | 23.7 ± 0.7c (100) | 68.4 ± 0.9a | 0.52 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahayaraj, K.; Subash, N.; Allingham, R.W.; Kumar, V.; Avery, P.B.; Mehra, L.K.; McKenzie, C.L.; Osborne, L.S. Lethal and Sublethal Effects of Three Microbial Biocontrol Agents on Spodoptera litura and Its Natural Predator Rhynocoris kumarii. Insects 2018, 9, 101. https://doi.org/10.3390/insects9030101
Sahayaraj K, Subash N, Allingham RW, Kumar V, Avery PB, Mehra LK, McKenzie CL, Osborne LS. Lethal and Sublethal Effects of Three Microbial Biocontrol Agents on Spodoptera litura and Its Natural Predator Rhynocoris kumarii. Insects. 2018; 9(3):101. https://doi.org/10.3390/insects9030101
Chicago/Turabian StyleSahayaraj, Kitherian, Natarajan Subash, Robert W. Allingham, Vivek Kumar, Pasco B. Avery, Lucky K. Mehra, Cindy L. McKenzie, and Lance S. Osborne. 2018. "Lethal and Sublethal Effects of Three Microbial Biocontrol Agents on Spodoptera litura and Its Natural Predator Rhynocoris kumarii" Insects 9, no. 3: 101. https://doi.org/10.3390/insects9030101
APA StyleSahayaraj, K., Subash, N., Allingham, R. W., Kumar, V., Avery, P. B., Mehra, L. K., McKenzie, C. L., & Osborne, L. S. (2018). Lethal and Sublethal Effects of Three Microbial Biocontrol Agents on Spodoptera litura and Its Natural Predator Rhynocoris kumarii. Insects, 9(3), 101. https://doi.org/10.3390/insects9030101