Abstract
Entomopathogenic nematodes from the two genera Steinernema and Heterorhabditis are widely used as biological agents against various insect pests and represent a promising alternative to replace pesticides. Efficacy and biocontrol success can be enhanced through improved understanding of their biology and ecology. Many endogenous and environmental factors influence the survival of nematodes following application, as well as their transmission success to the target species. The aim of this paper is to give an overview of the major topics currently considered to affect transmission success of these biological control agents, including interactions with insects, plants and other members of the soil biota including conspecifics.
1. Introduction
Entomopathogenic nematodes (EPNs) represent a guild of soil-inhabiting nematodes capable of infecting a wide range of insects. Their free-living infective juveniles (IJs) penetrate insect hosts through natural openings or through the cuticle [1] and release their symbiotic bacteria into the hemocoel. The host is usually rapidly killed by the ensuing septicemia, and its cadaver is converted from insect to nematode biomass, mediated by the action of the bacteria in digesting host tissues into a nutritious soup for the nematodes. The bacteria also protect the cadaver resource against competitors, by producing bacteriocins, antimicrobials and other antibiotics [2,3]. IJs develop to adults and give rise to several successive generations until the resources become scarce. New IJs are produced and emerge from the cadaver, dispersing and seeking fresh hosts to infect.
The two families Heterorhabditidae and Steinernematidae are widely studied, for two reasons: firstly, they both show great potential as inundative biological agents against numerous insect pests, in addition to or in replacement of chemical pesticides, and secondly, due to their short lifespan and ease of lab culture, they are increasingly used as model organisms in fundamental research into symbiosis and parasitism, inter alia. So far, about 90 species of Steinernematidae and 20 species of Heterorhabditidae have been described. However, only a handful of species are commercially produced for use in biological control [4], and these species (particularly Steinernema carpocapsae, Steinernema feltiae and Heterorhabditis bacteriophora) are also the ones that are most intensively researched [4,5,6,7,8]. Significant developments are being made towards increasing efficacy through strain selection and improved methods of production [9,10], formulation [11,12,13,14,15] and application [16,17,18,19]. Research on the biology and ecology of EPNs, such as mechanisms of infection and factors influencing their survival and behavior, underpins these developments and also helps make better predictions of their field performance.
EPN transmission strategies have been shaped by natural selection to optimize fitness of the nematodes themselves, and are of intrinsic interest to parasitologists and evolutionary ecologists. In this paper, we review factors currently considered to impact EPN transmission success, particularly those that impact on biocontrol, with emphasis on recent developments and literature.
2. Trait Diversity
Apart from some noticeable exceptions, such as Steinernema scapterisci which presents a narrow host range [20], most EPN species are capable of infecting a wide range of different insect species—at least when tested in the laboratory. However, the susceptibility of insect pests varies depending on EPN species. Although EPNs are already widely used in inundative biological pest control programs, a substantial part of current research effort concerns tests against additional insect pests, including groups as diverse as Dipteran, Lepidopteran, Isopteran, Hemipteran, Hymenopteran or Coleopteran species. Table 1 presents examples of recent studies (2014–present). Although some EPN species have been shown to be efficient against a large number of different pest species (e.g., H. bacteriophora or S. carpocapsae, see Table 1), several different species of nematodes are usually compared in order to identify the best species for the proposed purpose (e.g., [21,22,23,24,25]). Native species are often considered [23,25,26], as they are expected to be adapted to local conditions, ideally including the pest itself. Use of native species is also preferred for the purpose of limiting environmental risks. In addition, nematodes belonging to the same species but originating from different isolates can also drastically vary in their efficacy [27]. For instance, the mortality of Bactrocera dorsalis fruit flies induced by Heterorhabditis taysearae ranged from 51.2 to 96.1% depending on the isolate, despite all isolates originating from Benin [28]. Considering such important variation, finding the most efficient species/isolate is thus particularly important, and is quite a difficult task considering the large number of EPN species already described. Moreover, new species are regularly discovered. Within the five last years, at least two Heterorhabditidae and eight Steinernematidae have been described (Table 2), while already-known species were detected in new geographical locations [29,30]. More rarely, new species of bacteria are also described, such as Photorhabdus heterorhabditis, a symbiont of Heterorhabditis zealandica [31]. The laboratory screening step is of most use in rejecting species and strains with low virulence, but because many factors other than virulence are crucial to field success, it is advisable to bring more than one virulent strain to field testing. While screening different species and isolates is an important and practical step in a biocontrol program, the underlying reasons for species and strain specific differences in efficacy are rarely elucidated (Figure 1). These can include traits related to nematode behavior and virulence [32], while the bacterial symbiont is an important and often overlooked determinant of success of the nematode-bacterial complex. In the past, it was considered that each nematode species was associated with a particular symbiont species or subspecies [33], but it is becoming clear that certain species, particularly of Heterorhabditis, can associate with more than one symbiont (e.g., [34,35]). In nature, switching between symbionts with different properties can allow a nematode to effectively extend its niche. For instance, in desiccating conditions, Heterorhabditis downesi IJs had a higher reproductive success when they were associated with the bacteria Photorhabdus temperata subsp. cinerea compared to those associated with P. temperata subsp. temperata [35]. Reassociating nematodes with compatible but non-native symbionts is one possible route to strain improvement. However, any gain in virulence must be offset against a potential loss in other traits of the symbiosis, such as reproductive capacity [36]. Steinernema feltiae recombined with symbiont Xenorhabdus bovienni from different sources performed significantly better with the native symbiont and closely related strains than with those from more divergent sources [37].

Table 1.
Recent studies (2014—present) investigating the effect of entomopathogenic nematodes against insect pests.
Table 2.
Recently discovered species of entomopathogenic nematodes.
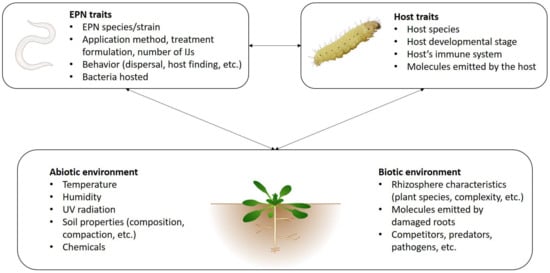
Figure 1.
Factors influencing EPNs survival and efficacy.
Promising results from laboratory experiments on the efficacy of EPNs against insect pests frequently do not translate into success in field and greenhouse, as shown for example in tests of EPNs against the African black beetle Heteronychus arator [38] and the carob moth Ectomyelois ceratoniae [39].
This is not surprising, given the complexity of the soil environment with many interacting biotic and abiotic factors impacting nematode survival and host finding (Figure 1). Entomopathogenic nematodes used as biocontrol agents are usually applied in high numbers [40], but only a fraction of these succeed in finding a host. A dramatic decline in IJ numbers has been documented following the application of nematodes [41], which has been attributed to the detrimental effect of UV light and desiccation at the soil surface. Studies have shown inter-specific variation in nematodes’ tolerance to UV [42] and to desiccation [43,44,45,46,47]. Adequate environmental moisture is essential for IJ survival and movement [48,49,50]. Although it is often difficult to control this parameter in the field, one solution resides in the addition of adjuvants [21,26,51,52,53], in particular in the case of folial application of EPNs that induce a higher desiccation stress [54,55]. For instance, both the survival rates and infectivity of Steinernema species against two Lepidopteran pests were increased by the use of a surfactant in the nematode suspension [56]. Similarly, the negative effect of UV on nematodes can be reduced by using certain UV protectants [55]. In contrast, other application methods can also be used rather than nematode suspensions, such as nematode-infected cadavers [16] or pre-infected live insect hosts [19], both of which gave promising results. A more technically advanced alternative is the use of nematode-filled capsules. While lacking the natural IJ dispersal factors that may be present in insect cadavers [57,58], the use of capsules has the advantage of being able to include attractants and feeding stimulants for the target pest, and this approach has been successfully trialed against Diabrotica virgifera virgifera in maize [59]. Attracting the pest to the nematodes, rather than the reverse, reduces the importance of several of the key elements in the nematodes’ natural transmission strategy (dispersal, host finding, and many of the interactions with soil biota), relying more on virulence for the target pest and also compatibility with the formulation.
Nematodes moving into the soil will be protected from UV and partly from desiccation, especially since IJs can adapt their vertical distribution according to the soil moisture and their own tolerance [60,61]. However, they remain vulnerable to other mortality factors, such as temperature extremes or predation, as well as starvation, resulting in a more gradual decline in their numbers over the weeks following the application. As for desiccation tolerance, nematodes have different thermal limits and optima depending on their species [43,44,62]. Temperature is probably the most important factor impacting EPN efficacy [23,25,38,48,51,63,64]. Temperature can also be used as a tool to improve EPN efficacy. For instance, conditioning S. carpocapsae and Heterorhabditis megidis for three weeks at 9 °C increased their efficacy against black vine weevils Otiorhynchus sulcatus [65].
3. Dispersal and Host Finding
The number of IJs exiting a host can range from tens of thousands to hundreds of thousands [101], while billions of them are applied to control pests [40]. We assume that each individual IJ has been shaped by natural selection to locate a suitable host in which to complete its life cycle, though strains may since have been artificially selected for improved biocontrol [102]. Even strains that have been bred for improved biocontrol performance and produced under conditions that are as uniform as possible consist of individual IJs, each with its unique history of gene x environment interactions [103]. IJs’ behavior changes as they age. The temporal change that receives most attention is the decline in “quality”, mostly measured as infectivity, as IJs age [104]. IJs are non-feeding and rely on a finite quantity of energy reserves that are critical for their survival and infection success. In particular, lipids and glycogen contents have been linked to infectivity in several nematode species [105,106,107]. To increase their chances of finding a suitable host, IJs employ a variety of strategies, and two types of model can be used to help conceptualize this [108]. Hierarchical models, developed for other parasites, divide the transmission process into a set of steps that may include dispersal, host-habitat location, host location and host acceptance, with each phase (phases are not necessarily distinct) characterized by responses to specific stimuli [108,109]. While each of these processes is studied in EPNs, the temporal phasing of them has received less attention, but we can assume that rapidly after their emergence from an insect host or their application as biocontrol agents, IJs move away in an initial dispersal phase characterized by random movements as well as potential responses to environmental signals that might bring them to their hosts’ habitat [109]. Dispersal of IJs is stimulated by factors associated with the spent cadaver [57]. Among the factors responsible are ascarosides, a diverse class of signaling molecules that regulate development and social behaviors in Caenorhabditis elegans and more widely among nematodes [58]. Ascaroside #9 accumulated in nematode-infected cadavers and in bioassays caused S. feltiae IJs to become more active and move away from the source location [58]. Once in the host’s habitat, nematodes rely on different types of signals, mostly chemicals [110] (although vibrations or temperature gradients might also be used [111,112,113]), bringing them to the area of the habitat that is modified by the presence of the host (host’s “active space”) [109], and ultimately to the host itself.
A second conceptual model involves a distinction between EPN species as ambush or cruise foragers. Heterorhabditis species are characterized as cruisers, moving actively through the soil in search of their hosts, while the behavior of Steinernema species varies from cruisers to ambushers; ambushers mostly remain at the surface of the soil and lift their body into the air (nictation) or exhibit jumping behavior to attach to passing insects [114,115,116]. The behavior of H. bacteriophora and S. carpocapsae was recently studied in mesocosms [117]. In accordance with the cruiser-ambush theory, H. bacteriophora was more efficient at infecting non-mobile hosts (Galleria mellonella larvae maintained in cages) compared to mobile hosts, while the contrary was observed for S. carpocapsae [117]. Despite their foraging strategy, ambusher species also need to disperse. In another mesocosm experiment, a majority of S. carpocapsae were found to ambush near the source cadaver, while a majority of H. bacteriophora dispersed away from it [118]. However, around 4% of S. carpocapsae IJs exhibited a “sprinter” behavior, dispersing faster than the fastest H. bacteriophora and reaching 30–61 cm (in comparison, only 2% of H. bacteriophora IJs reached that far) [118]. The two different strategies are likely based on trade-offs with other parameters. Indeed, although S. carpocapsae responded promptly to the artificial selection of the fastest dispersers, the resulting enhanced dispersal was associated with reduced reproduction and nictation abilities [119]. The distinction between species based on foraging strategy should not be over-relied on for accurately predicting field success of a species. For example, S. carpocapsae, which is classified as an ambusher, was able to infect large pine weevils (Hylobius abietis) under the bark of tree roots as far as 30 cm deep in the soil [120]. In this and similar examples, the nematodes may be using tree roots as “routeways” to facilitate movement through soil.
Several factors are known to influence IJs dispersal, such as the vegetation [118,121] or the soil properties, in terms of composition and compaction [122]. Along with the presence of appropriate hosts, the impact of such factors on EPN survival and dispersal might explain the distribution of nematodes in habitats differing in plant communities or soil parameters [123,124]. Some insects can also serve as phoretic dispersers, such as the beetle Calosoma granulatum, recently shown to be capable of transporting Heterorhabditis amazonensis over long distances (over 40 cm) [125]. Recently, attention has been focused on the dispersal of IJs in groups, rather than as individuals [126,127]. Both Heterorhabditis indica (cruiser-type forager) and S. carpocapsae (ambusher-type forager) performed aggregative movement patterns, including in their dispersal phase [128]. Such group movements might increase the survival of IJs, through reduced evaporation rates in desiccating conditions, or through protection against predators (effect of dilution). It could also ensure a better infection success with the penetration of several IJs in a host, where the probability of finding a suitable mate would also be increased [128], though if nematodes regularly travel and mate with individuals from the same natal host, this should increase the risk of inbreeding. It is currently unknown to what extent group movement is a product of physical forces acting on the nematodes, or involves some form of chemical signaling.
Eventually, the random dispersal of IJs might bring them into zones in which signals from the host or its habitat will be used for directing movements. In addition to the universally produced CO2 which is attractive for IJs of all EPN species [129], many specific host-derived odorants that stimulate host-seeking behavior by IJs have been identified and shown to be differentially attractive to different EPN species [110]. Recently it was shown that Steinernema diaprepesi and H. indica were attracted not only to beetle frass, but also to the sex pheromones of their weevil host [130]. Entomopathogenic nematodes widely use root-feeding insects as hosts. Consequently, stimuli initiating active search from IJs originate not only from the host itself, but also from roots, as indicators of likely host habitat, and more particularly damaged roots, indicative of the presence of the host. Although not specific to the host species, following a CO2 gradient is likely to bring nematodes into the rhizosphere, where potential hosts might then be detected based on their own chemicals or on chemicals emitted by the plant. While roots themselves are attractive, roots that are wounded by the feeding of herbivores emit volatiles or blends that specifically attract EPNs, a form of signaling that benefits both plants and nematodes [131,132]. This effect has been demonstrated for several species of plant, including recently for carrot, vine, fig and sugarcane [133,134,135]. In order to directly examine the foraging behavior of nematodes, Li et al. used pluronic gel, a transparent polymer allowing 3D observation of nematodes inside the gel. In accordance with previous results, they found that IJs preferentially aggregated around the wounded parts of roots [136]. In addition to the release of attractive compounds, roots might also provide nematodes with physical “routeways”, facilitating their progression into the soil [137]. Moreover, the complexity of the root architecture can interact with the herbivore-induced plant volatiles to impact the foraging behavior of nematodes. Using artificial model-roots with different degrees of complexity and connectivity, Demarta et al. found that host-finding by H. megidis was facilitated by low root complexity. However, the addition of a synthetic root volatile, (E)-β-caryophyllene, changed this pattern and favored the nematodes foraging on the most complex model-roots [138]. The effect of (E)-β-caryophyllene on the behavior of H. megidis was also found to depend on the type of soil, while the diffusion of the compound in the soil depended on its humidity [139]. Aboveground stimulation can also influence the recruitment of EPNs in the soil. In particular, the stimulation of the salicylic acid pathway, a plant defense pathway, led to the attraction of S. diaprepesi nematodes in the absence of their hosts [50]. Filgueiras et al. also found a strong effect of elicitors of plant defense applied on leaves of corn seedlings on the recruitment of H. amazonensis nematodes and suggested that treating crops with elicitors might be a good strategy to increase the success of the biological control of EPNs [140].
Recent evidence suggests that the response of nematodes to volatile compounds such as (E)-β-caryophyllene or pinene could be modified by their experience (previous exposure to the compound) [141]. Intriguingly, the behavior of H. indica individuals that were not previously exposed to α-pinene depended on interspecific relationships; although naive H. indica alone were repelled by α-pinene, they exhibited a preference for this compound when they were associated to previously-exposed S. diaprepesi, [141].
4. Infection
Having located a host, IJs may choose to infect or not, depending on its suitability. Since IJs of different species respond differently to different species of insect [110], some amount of host selection may already be accomplished during the host-seeking phase. Apart from some specialist species [20], most EPNs can infect a wide range of different host species, at various developmental stages, with substantial differences in susceptibility to nematodes both between species and between developmental stages of the same species. Insect larvae and pupae are often found to be more susceptible to EPN infection than adults [25,85,142]. Such differences in susceptibility can result from various mechanisms, such as differences in behavior (in particular higher activity levels and avoidance behaviors in adults), immune system or physical barriers to nematode penetration. For instance, the low susceptibility of certain pupae, which was also observed in other studies [49,77], could be due to a lack of natural entry routes for nematodes, as well as a tougher cuticle [76]. Susceptibility can also vary among different larval instars, as was observed in armyworms and mosquitoes [72,88]. Interestingly, large pine weevils (H. abietis) infected by H. downesi or S. carpocapsae as pupae died as adults, suggesting that IJs can infect pupae, survive through their host metamorphosis and kill the adults [142]. In social insects, variation in susceptibility was observed among different castes. While Steinernema karii induced a higher mortality in workers of the termite species Coptotermes formosanus compared to soldiers [82], the opposite effect was observed for the two other termite species Macrotermes bellicosus and Trinervitermes occidentalis exposed to H. indica and Heterorhabditis sonorensi [83]. Finally, although EPNs are typically thought of as infecting and killing live insect hosts, IJs can also act as scavengers. For instance, although their progeny was lower than that recorded in live hosts, both Steinernema kraussei and H. megidis were able to reproduce in freeze-killed G. mellonella larvae [143]. The ability to utilize additional resources for reproduction may facilitate persistence of EPN populations in soil post application.
Differences in the success of nematode infection documented above can be partly due to the response of the host’s immune system to the nematode-bacteria complex. The immune response of insects to EPNs and the mechanisms used by nematodes and bacteria to evade or defeat it have been extensively studied in recent years (see the review by Eleftherianos et al. [144]). Following the penetration of IJs into the insect host through natural openings and their establishment in the hemolymph, the IJs then release their symbiotic bacteria. These are either from the Photorhabdus genus for Heterorhabditis species, or from the Xenorhabdus genus for Steinernema nematodes. The subsequent release of toxic and immunosuppressive compounds by the bacteria cause death of the host by septicemia [145,146]. Axenic nematodes, especially Steinernema species, are also capable of causing the death of their hosts [146,147], in particular through the release of a venom containing a high proportion of proteases (serine carboxypeptidases, trypsins, eukaryotic aspartyl proteases, zinc carboxypeptidases) and protease inhibitors [147]. However, the association between nematodes and bacteria is important in the virulence of the complex [148]. In an experiment where nematodes from different Steinernema species were isolated from their symbiotic bacteria, combinations of nematodes with “foreign” bacteria resulted in a reduction of the virulence of the complex against G. mellonella larvae, as well as a reduction in nematode progeny [149].
Following host penetration, the release of bacteria by nematodes is usually delayed in the host by 30 min for Heterorhabditis species and several hours for Steinernema nematodes [150]. There is thus a possibility for the insect to neutralize its parasite before the bacterial challenge. Insects’ reactions to nematode infection include the melanization and encapsulation of the parasites with hemocytes. Many immune factors have been shown to vary in the hemolymph of the host following the entry of nematodes, including both humoral and cellular responses. For instance, many studies documented modifications through time in the hemocyte counts [151,152,153] and the enzyme activity of the host such as proteases and phenoloxidase [152,153,154,155]. Nematodes and their symbiotic bacteria can both inhibit their hosts’ immune system, suppressing the melanization process and depressing the antimicrobial response [146,153,156]. Since IJs are applied in high numbers, it is likely that at least some hosts get infected by numerous nematodes. Such multiple infections, although leading to competition between nematodes (see below), lower the risk of them being overcome by the host’s immune system. However, Kenney and Eleftherianos suggested that the success of infection of EPNs used in biocontrol programs could be improved by developing nematode-bacteria complexes that produce proteins necessary for evasion of the host’s immune system, or nematodes with a short delay in the ejection of their bacterial symbionts [157].
5. Interaction with the Biotic and Abiotic Environment
As shown above, environmental parameters such as temperature, humidity, types of vegetation and soil properties can affect nematode survival and virulence, while infection also depends on complex interactions between IJs, their symbiotic bacteria and the host (Figure 1). However, other abiotic parameters and biotic interactions are particularly worthy of interest, especially when considering nematodes as biocontrol agents.
Nematodes applied inundatively to soil can affect other species, including non-target hosts or potential competitors such as indigenous nematodes [158]. Studies investigating the environmental risks linked to the use of EPNs generally re-affirm the low impact of this form of biocontrol on non-target species [159]. For instance, non-target species were unaffected by the use of H. bacteriophora applied against corn rootworm larvae in maize [160]. The effect on non-target species will depend on the biology of these species, and laboratory studies can help identify susceptible species and formulate recommendations for risk reduction, if considered necessary [161]. Special consideration needs to be given to ecologically important species, and potential impact on their roles in ecosystems. However, as for tests against target species, laboratory tests of susceptibility of important non-targets may not accurately predict impact in the field. Recently, bumble bees (Bombus terrestris) were reported to suffer 80% mortality when exposed in the laboratory to soil treated with EPNs at recommended field concentrations [162]. Moreover, IJs were able to proliferate in the bee cadavers, which the authors suggested may represent a potential threat for other members of their colony [162]. However, factors such as the preferred habitat for bumble bee nests, the depth and structure of the nests, and behavior of infected individuals [163] or their hygienic colony mates [164], will likely present significant ecological barriers to EPN infection of bumble bees. Safety may also be examined in terms of persistence of EPNs in the treated area. In a field experiment, S. carpocapsae was detected in the soil up to two years after application, but not after four years, and its presence was positively correlated with that of its large pine weevil host [165], an optimistic result regarding potential environmental risk.
EPNs used for biocontrol are also likely to interact with other organisms, especially those belonging to the trophic network of their hosts. Within the few days following infection, host cadavers release scavenger deterrent factors (SDF), chemical compounds produced by nematodes’ symbiotic bacteria that deter potential scavengers and thus protect the host cadaver [166]. A wide range of species, including various arthropods [166,167] and birds [168], are known to be sensitive to such factors, avoiding infected cadavers or promptly rejecting them after an attempt to consume them. Recently, laboratory tests on three cyprinid fish species showed they were also sensitive to SDFs, rejecting G. mellonella larvae killed by Heterorhabditis or Steinernema species, and preferentially feeding on freeze-killed mosquito (Aedes aegypti) larvae compared to nematode-killed ones [169]. Tests on the carabid beetle Carabus granulatum, a predator of the lepidopteran host of H. amazonensis, showed that larvae and adults both avoided feeding on infected larvae when they had a choice [170]. In forced feeding experiments, the consumption of infected larvae led to a high mortality of the predator [170]. However, not all insects are repelled by nematode-infected hosts. For instance, while crickets and springtails avoided Steinernema-killed insects, consumption was observed for ants, cockroaches, mites and earwigs [171]. In addition, springtails Sinella curviseta and Folsomia candida were found to directly consume IJs [171]. As suggested by the authors, the scavenger and predator effect of soil arthropods could have a top-down regulatory impact on nematodes.
Nematodes can also be in direct competition with other organisms. In the case of the carabid C. granulatum cited above [170], the predator is not only a threat for the integrity of the host cadaver, and thus for the reproduction success of nematodes; IJs and beetles compete for the same resources, that is, living prey or hosts. Consequently, the inundative use of EPNs could decrease the number of insect prey available for predators, with consequences on their populations. Parasitoid insects also compete for the same resource, as well as being susceptible to infection by nematodes themselves. Despite negative interactions demonstrated in the laboratory, this is less often documented in the field [172,173]. For example, in field trials, the use of nematodes against Caribbean fruit flies Anastrepha suspense did not reduce the density of their emerging parasitoid Diachasmimorpha longicaudata [174]. The most direct competitors of EPNs might be nematodes themselves, either indigenous entomopathogenic nematodes or free-living nematodes (FLN). For instance, FLNs from the genus Oscheius show scavenger behavior and are capable of reproducing in freeze-killed as well as already infected insect cadavers [175]. Competition with Oscheius species in host cadavers resulted in a substantial reduction in the number of EPN progeny, with inter-specific differences in their ability to overcome intraguild competition [175,176]. A more recent study found contradictory results with no evidence of competition between two Oscheius species and EPNs [143]. Apart from competition for resources, a direct interspecific killing behavior has also been documented among four Steinernema species, as well as intra-specific killing among males [177,178]. Because of their negative interaction on other nematodes, EPNs can be used to control pest species. Many studies already documented the efficacy of EPNs against pest nematodes (reviewed by Kenney and Eleftherianos [157]). For instance, several species of EPNs, and particularly their bacteria, proved to be efficient against the root-knot nematode Meloidogyne incognita, a parasite of tomato plants [179,180,181]. More complex and unexpected interactions between EPNs or their bacteria and other organisms are reported, such as the induction by EPNs of systemic resistance in tomatoes against insect pests and bacterial diseases [182].
The plant host can also affect the susceptibility of insects feeding on it. For instance, Steinernema riobrave nematodes showed a reduced virulence and reproduction on Helicoverpa zea larvae reared on tobacco plants compared to those feeding on tomato and eggplants [183]. There is an increasing use of GMO plants in crops, many of them producing their own insecticidal proteins. In a recent study, no significant effects on the virulence, reproduction and host preference of H. bacteriophora were found when diamondback moth (Plutella xylostella) was reared on broccoli leaves genetically modified to produce CrylAc proteins [184]. Additional levels continue to be added to the inter-organismal interactions in which EPNs are implicated—most recently, the insect host’s endosymbionts. For example, the presence of Wolbachia endosymbionts promoted the survival of Drosophila melanogaster infected with S. carpocapsae through effects on the insect’s immune response [185].
6. Conclusions and Future Directions
The use of EPNs against pests is particularly timely given the negative impact of the use of chemical pesticides. Applied research largely focuses on improving their use on a large scale, through improved nematode culture methods, storage, treatment formulations and application methods. Spreading billions of nematodes in places where they are not always native represents a potential environmental risk that deserves attention. In this respect, long-term studies combining laboratory experiments followed by field tests are particularly relevant. In addition, a better understanding of the biology and ecology of EPNs is likely to give new directions in their use against pests. It is now getting easier to access large datasets of information through proteomic and transcriptomic studies. For instance, such approaches were used to describe nematode infection. The venom proteome of S. carpocapsae IJs was recently analyzed using mass spectrometry, highlighting the presence of 472 venom proteins including numerous proteases and protease inhibitors, as well as toxin-related proteins potentially linked to the suppression of host immune system [147]. In addition, the host immune response could be better understood with transcriptomic analyses following nematodes infection [186,187,188]. Genetic analyses, including sequencing, are particularly useful to better understand the diversity of nematodes [189,190] and their symbiotic bacteria [191,192], and to study their phylogenetic relationships [193]. Such phylogenies can help understand the evolution of certain traits. For instance, Blackburn et al. reconstituted the phylogenetic relationships of 18 strains of Photorhabdus and also tested their virulence against two hosts. Results showed a general evolutionary trend towards an increase in virulence in Photorhabdus [194]. Finally, genomic and proteomic data provide a large number of genes that are potential candidates for the improvement of EPNs, from the survival of IJs in certain conditions to an increase transmission and killing efficacy. Although artificial selection solely based on phenotypic traits can lead to improved nematodes [195], understanding the mechanisms might lead to better efficacy, especially on traits where artificial selection is complicated [196].
Author Contributions
Both authors participated in the writing of this review
Funding
This work was supported by the European Union Seventh Framework Programme (grant agreement no 612713—BIOCOMES Project).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Poinar, G.O. Nematodes for Biological Control of Insects; CRC Press, Inc.: Boca Raton, CA, USA, 1979. [Google Scholar]
- Akhurst, R.J. Antibiotic activity of Xenorhabdus spp., bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditidae and Steinernematidae. J. Gen. Microbiol. 1982, 128, 3061–3065. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B. Entomopathogenic bacteria as a source of secondary metabolites. Curr. Opin. Chem. Biol. 2009, 13, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Campos-Herrera, R. Nematode Pathogenesis of Insects and Other Pests; Springer International Publishing: Berlin, Germany, 2015; ISBN 978-3-319-18265-0. [Google Scholar]
- Grewal, P.S.; Ehlers, R.U.; Shapiro-Ilan, D.I. Nematodes as Biocontrol Agents; CAB International: Wallingford, UK, 2005. [Google Scholar]
- Gaugler, R.R. Entomopathogenic Nematology; CABI: Wallingford, UK, 2002. [Google Scholar]
- Laznik, Ž.; Trdan, S. Entomopathogenic nematodes (Nematoda: Rhabditida) in Slovenia: From tabula rasa to implementation into crop production systems. In Insecticides—Advances in Integrated Pest Management; Perveen, F., Ed.; InTech: Rijeka, Croatia, 2011; pp. 627–656. [Google Scholar]
- Leite, L.G.; Shapiro-Ilan, D.I.; Hazir, S.; Jackson, M.A. Effect of inoculum age and physical parameters on in vitro culture of the entomopathogenic nematode Steinernema feltiae. J. Helminthol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Testa, A.M.; Shields, E.J. Low labor “in vivo” mass rearing method for entomopathogenic nematodes. Biol. Control 2017, 106, 77–82. [Google Scholar] [CrossRef]
- Kim, J.; Jaffuel, G.; Turlings, T.C.J. Enhanced alginate capsule properties as a formulation of entomopathogenic nematodes. BioControl 2015, 60, 527–535. [Google Scholar] [CrossRef]
- Guo, W.; Yan, X.; Han, R. Adapted formulations for entomopathogenic nematodes, Steinernema and Heterorhabditis spp. Nematology 2017, 19, 587–596. [Google Scholar] [CrossRef]
- Cortés-Martínez, C.I.; Lewis, E.E.; Ruiz-Vega, J.; Martínez-Gutiérrez, G.A. Mechanical production of pellets for the application of entomopathogenic nematodes: Effect of pre-acclimation of Steinernema glaseri on its survival time and infectivity against Phyllophaga vetula. Biocontrol Sci. Technol. 2017, 27, 940–951. [Google Scholar] [CrossRef]
- Kagimu, N.; Ferreira, T.; Malan, A.P. The attributes of survival in the formulation of entomopathogenic nematodes utilised as insect biocontrol agents. Afr. Entomol. 2017, 25, 275–291. [Google Scholar] [CrossRef]
- Cruz-Martínez, H.; Ruiz-Vega, J.; Matadamas-Ortíz, P.T.; Cortés-Martínez, C.I.; Rosas-Diaz, J. Formulation of entomopathogenic nematodes for crop pest control—A review. Plant Prot. Sci. 2017, 53, 15–24. [Google Scholar] [CrossRef]
- Bai, G.Y.; Xu, H.; Fu, Y.Q.; Wang, X.Y.; Shen, G.S.; Ma, H.K.; Feng, X.; Pan, J.; Gu, X.S.; Guo, Y.Z.; Ruan, W.B.; Shapiro-Ilan, D.I. A comparison of novel entomopathogenic nematode application methods for control of the chive gnat, Bradysia odoriphaga (Diptera: Sciaridae). J. Econ. Entomol. 2016, 109, 2006–2013. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Mizell, R.F.; Horton, D.L. Curative control of the peachtree borer using entomopathogenic nematodes. J. Nematol. 2016, 48, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Mizell, R.F.; Horton, D.L.; Zaid, A. Field suppression of the peachtree borer, Synanthedon exitiosa, using Steinernema carpocapsae: Effects of irrigation, a sprayable gel and application method. Biol. Control 2015, 82, 7–12. [Google Scholar] [CrossRef]
- Gumus, A.; Karagoz, M.; Shapiro-Ilan, D.; Hazir, S. A novel approach to biocontrol: Release of live insect hosts pre-infected with entomopathogenic nematodes. J. Invertebr. Pathol. 2015, 130, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Sepulveda, C.; Dillman, A.R. Infective juveniles of the entomopathogenic nematode Steinernema scapterisci are preferentially activated by cricket tissue. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, N.; Karimi, J.; Hosseini, M.; Goldani, M.; Campos-Herrera, R. Pathogenicity of two species of entomopathogenic nematodes against the greenhouse whitefly, Trialeurodes vaporariorum (Hemiptera: Aleyrodidae), in laboratory and greenhouse experiments. J. Nematol. 2015, 47, 60–66. [Google Scholar] [PubMed]
- Azarnia, S.; Abbasipour, H.; Saeedizadeh, A.; Askarianzadeh, A. Laboratory assay of entomopathogenic nematodes against clearwing moth (Lepidoptera: Sesiidae) larvae. J. Entomol. Sci. 2018, 53, 62–69. [Google Scholar] [CrossRef]
- Kepenekci, İ.; Hazir, S.; Özdem, A. Evaluation of native entomopathogenic nematodes for the control of the European cherry fruit fly Rhagoletis cerasi L. (Diptera: Tephritidae) larvae in soil. Turkish J. Agric. For. 2015, 39, 74–79. [Google Scholar] [CrossRef]
- Manzoor, M.; Ahmad, J.N.; Sharif, M.Z.; Majeed, D.; Kiran, H.; Jafir, M.; Ali, H. Comparative effectiveness of entomopathogenic nematodes against red palm weevil (Rhynchophorus ferrugineus) in Pakistan. J. Entomol. Zool. Stud. JEZS 2017, 5, 756–760. [Google Scholar]
- Odendaal, D.; Addison, M.F.; Malan, A.P. Entomopathogenic nematodes for the control of the codling moth (Cydia pomonella L.) in field and laboratory trials. J. Helminthol. 2016, 90, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Odendaal, D.; Addison, M.F.; Malan, A.P. Control of diapausing codling moth, Cydia pomonella (Lepidoptera: Tortricidae) in wooden fruit bins, using entomopathogenic nematodes (Heterorhabditidae and Steinernematidae). Biocontrol Sci. Technol. 2016, 26, 1504–1515. [Google Scholar] [CrossRef]
- Laznik, Ž.; Tóth, T.; Lakatos, T.; Vidrih, M.; Trdan, S. The activity of three new strains of Steinernema feltiae against adults of Sitophilus oryzae under laboratory conditions. J. Food Agric. Environ. 2010, 8, 150–154. [Google Scholar]
- Godjo, A.; Zadji, L.; Decraemer, W.; Willems, A.; Afouda, L. Pathogenicity of indigenous entomopathogenic nematodes from Benin against mango fruit fly (Bactrocera dorsalis) under laboratory conditions. Biol. Control 2018, 117, 68–77. [Google Scholar] [CrossRef]
- Yan, X.; Waweru, B.; Qiu, X.; Hategekimana, A.; Kajuga, J.; Li, H.; Edgington, S.; Umulisa, C.; Han, R.; Toepfer, S. New entomopathogenic nematodes from semi-natural and small-holder farming habitats of Rwanda. Biocontrol Sci. Technol. 2016, 26, 820–834. [Google Scholar] [CrossRef]
- Gökçe, C.; Erbaş, Z.; Yilmaz, H.; Demirbağ, Z.; Demir, İ. A new entomopathogenic nematode species from Turkey, Steinernema websteri (Rhabditida: Steinernematidae), and its virulence. Turkish J. Biol. 2015, 39, 167–174. [Google Scholar] [CrossRef]
- Ferreira, T.; van Reenen, C.A.; Endo, A.; Tailliez, P.; Pagès, S.; Spröer, C.; Malan, A.P.; Dicks, L.M.T. Photorhabdus heterorhabditis sp. nov., a symbiont of the entomopathogenic nematode Heterorhabditis zealandica. Int. J. Syst. Evol. Microbiol. 2014, 64, 1540–1545. [Google Scholar] [CrossRef] [PubMed]
- Griffin, C.T. Perspectives on the behavior of entomopathogenic nematodes from dispersal to reproduction: Traits contributing to nematode fitness and biocontrol efficacy. J. Nematol. 2012, 44, 177–184. [Google Scholar] [PubMed]
- Adams, B.J.; Fodor, A.; Koppenhöfer, H.S.; Stackebrandt, E.; Patricia Stock, S.; Klein, M.G. Biodiversity and systematics of nematode-bacterium entomopathogens. Biol. Control 2006, 37, 32–49. [Google Scholar] [CrossRef]
- Kazimierczak, W.; Lis, M.; Skrzypek, T.; Kreft, A. Comparison of the methods applicable for the pathogenicity assessment of entomopathogenic nematodes. BioControl 2018, 63, 289–298. [Google Scholar] [CrossRef]
- Maher, A.M.D.; Asaiyah, M.A.M.; Brophy, C.; Griffin, C.T. An entomopathogenic nematode extends its niche by associating with different symbionts. Microb. Ecol. 2017, 73, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Stuart, R.J.; McCoy, C.W. Targeted improvement of Steinernema carpocapsae for control of the pecan weevil, Curculio caryae (Horn) (Coleoptera: Curculionidae) through hybridization and bacterial transfer. Biol. Control 2005, 34, 215–221. [Google Scholar] [CrossRef]
- Murfin, K.E.; Lee, M.M.; Klassen, J.L.; McDonald, B.R.; Larget, B.; Forst, S.; Stock, S.P.; Currie, C.R.; Goodrich-Blair, H. Xenorhabdus bovienii strain diversity impacts coevolution and symbiotic maintenance with Steinernema spp. nematode hosts. mBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.; Wilson, D.J.; Rodgers, A.; Gerard, P.J. Developing a strategy for using entomopathogenic nematodes to control the African black beetle (Heteronychus arator) in New Zealand pastures and investigating temperature constraints. Biol. Control 2016, 93. [Google Scholar] [CrossRef]
- Memari, Z.; Karimi, J.; Kamali, S.; Goldansaz, S.H.; Hosseini, M. Are entomopathogenic nematodes effective biological control agents against the carob moth, Ectomyelois ceratoniae? J. Nematol. 2016, 48, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Han, R.; Dolinksi, C. Entomopathogenic nematode production and application technology. J. Nematol. 2012, 44, 206–217. [Google Scholar] [PubMed]
- Smits, P.H. Post-application persistence of entomopathogenic nematodes. Biocontrol Sci. Technol. 1996, 6, 379–388. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Hazir, S.; Lete, L. Viability and virulence of entomopathogenic nematodes exposed to ultraviolet radiation. J. Nematol. 2015, 47, 184–189. [Google Scholar] [PubMed]
- Shapiro-Ilan, D.I.; Brown, I.; Lewis, E.E. Freezing and desiccation tolerance in entomopathogenic nematodes: Diversity and correlation of traits. J. Nematol. 2014, 46, 27–34. [Google Scholar] [PubMed]
- Ulu, T.C.; Susurluk, I.A. Heat and desiccation tolerances of Heterorhabditis bacteriophora strains and relationships between their tolerances and some bioecological characteristics. Invertebr. Surviv. J. 2014, 11, 4–10. [Google Scholar]
- Menti, H.; Wright, D.J.; Perry, R.N. Desiccation survival of populations of the entomopathogenic nematodes Steinernema feltiae and Heterorhabditis megidis from Greece and the UK. J. Helminthol. 1997, 71, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.N.; Perry, R.N.; Wright, D.J. Desiccation survival and water contents of entomopathogenic nematodes, Steinernema spp. (Rhabditida: Steinernematidae). Int. J. Parasitol. 1997, 27, 61–70. [Google Scholar] [CrossRef]
- Piggott, S.J.; Liu, Q.Z.; Glazer, I.; Wright, D.J. Does osmoregulatory behaviour in entomopathogenic nematodes predispose desiccation tolerance? Nematology 2002, 4, 483–487. [Google Scholar] [CrossRef]
- Baimey, H.; Zadji, L.; Afouda, L.; Moens, M.; Decraemer, W. Influence of pesticides, soil temperature and moisture on entomopathogenic nematodes from southern Benin and control of underground termite nest populations. Nematology 2015, 17, 1057–1069. [Google Scholar] [CrossRef]
- Langford, E.A.; Nielsen, U.N.; Johnson, S.N.; Riegler, M. Susceptibility of Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae), to entomopathogenic nematodes. Biol. Control 2014, 69, 34–39. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Willett, D.S.; Junior, A.M.; Pareja, M.; El-Borai, F.E.; Dickson, D.W.; Stelinski, L.L.; Duncan, L.W.; Moino, A.; Pareja, M.; et al. Stimulation of the salicylic acid pathway aboveground recruits entomopathogenic nematodes belowground. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, V.M.; Beck, B.K.; Berckmoes, E.; Moerkens, R.; Wittemans, L.; De Vis, R.; Nuyttens, D.; Casteels, H.F.; Maes, M.; Tirry, L.; et al. Efficacy of entomopathogenic nematodes against larvae of Tuta absoluta in the laboratory. Pest Manag. Sci. 2016, 72, 1702–1709. [Google Scholar] [CrossRef] [PubMed]
- Portman, S.L.; Krishnankutty, S.M.; Reddy, G.V.P. Entomopathogenic nematodes combined with adjuvants presents a new potential biological control method for managing the wheat stem sawfly, Cephus cinctus (Hymenoptera: Cephidae). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Dito, D.F.; Shapiro-Ilan, D.I.; Dunlap, C.A.; Behle, R.W.; Lewis, E.E. Enhanced biological control potential of the entomopathogenic nematode, Steinernema carpocapsae, applied with a protective gel formulation. Biocontrol Sci. Technol. 2016, 26, 835–848. [Google Scholar] [CrossRef]
- Mason, J.M.; Matthews, G.A.; Wright, D.J. Screening and selection of adjuvants for the spray application of entomopathogenic nematodes against a foliar pest. Crop Prot. 1998, 17, 463–470. [Google Scholar] [CrossRef]
- Wright, D.J.; Peters, A.; Schroer, S.; Fife, J.P. Application technology. In Nematodes as Biocontrol Agents; Grewal, P., Ehlers, R.-U., Shapiro-Ilan, D.I., Eds.; CABI: Wikiwand, UK, 2005; pp. 91–106. [Google Scholar]
- Noosidum, A.; Satwong, P.; Chandrapatya, A.; Lewis, E.E. Efficacy of Steinernema spp. plus anti-desiccants to control two serious foliage pests of vegetable crops, Spodoptera litura F. and Plutella xylostella L. Biol. Control 2016, 97, 48–56. [Google Scholar] [CrossRef]
- Shapiro, D.I.; Glazer, I. Comparison of entomopathogenic nematode infectivity from infected hosts versus aqueous suspension. Environ. Entomol. 1996, 25, 1455–1461. [Google Scholar] [CrossRef]
- Kaplan, F.; Alborn, H.T.; von Reuss, S.H.; Ajredini, R.; Ali, J.G.; Akyazi, F.; Stelinski, L.L.; Edison, A.S.; Schroeder, F.C.; Teal, P.E. Interspecific nematode signals regulate dispersal behavior. PLoS ONE 2012, 7, e38735. [Google Scholar] [CrossRef] [PubMed]
- Hiltpold, I.; Hibbard, B.E.; French, B.W.; Turlings, T.C.J. Capsules containing entomopathogenic nematodes as a Trojan horse approach to control the western corn rootworm. Plant Soil 2012, 358, 11–25. [Google Scholar] [CrossRef]
- Salame, L.; Glazer, I. Stress avoidance: Vertical movement of entomopathogenic nematodes in response to soil moisture gradient. Phytoparasitica 2015, 43, 647–655. [Google Scholar] [CrossRef]
- El-Borai, F.; Killiny, N.; Duncan, L.W. Concilience in entomopathogenic nematode responses to water potential and their geospatial patterns in Florida. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.P.; Malan, A.P.; Terblanche, J.S. Divergent thermal specialisation of two South African entomopathogenic nematodes. PeerJ 2015, 3, e1023. [Google Scholar] [CrossRef] [PubMed]
- Kamali, S.; Karimi, J.; Koppenhöfer, A.M. New insight into the management of the tomato leaf miner, Tuta absoluta (Lepidoptera: Gelechiidae) with entomopathogenic nematodes. J. Econ. Entomol. 2018, 111, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Youngman, R.R.; Kok, L.T.; Laub, C.A.; Pfeiffer, D.G. Interaction between entomopathogenic nematodes and entomopathogenic fungi applied to third instar southern masked chafer white grubs, Cyclocephala lurida (Coleoptera: Scarabaeidae), under laboratory and greenhouse conditions. Biol. Control 2014, 76, 65–73. [Google Scholar] [CrossRef]
- Guy, A.; Gaffney, M.; Kapranas, A.; Griffin, C.T. Conditioning the entomopathogenic nematodes Steinernema carpocapsae and Heterorhabditis megidis by pre-application storage improves efficacy against black vine weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae) at low and moderate temperatures. Biol. Control 2017. [Google Scholar] [CrossRef]
- Kajuga, J.; Hategekimana, A.; Yan, X.; Waweru, B.W.; Li, H.; Li, K.; Yin, J.; Cao, L.; Karanja, D.; Umulisa, C.; et al. Management of white grubs (Coleoptera: Scarabeidae) with entomopathogenic nematodes in Rwanda. Egypt. J. Biol. Pest Control 2018, 28. [Google Scholar] [CrossRef]
- Demir, S.; Karagoz, M.; Hazir, S.; Kaya, H.K. Evaluation of entomopathogenic nematodes and their combined application against Curculio elephas and Polyphylla fullo larvae. J. Pest Sci. 2015, 88, 163–170. [Google Scholar] [CrossRef]
- Guo, W.; Yan, X.; Zhao, G.; Chen, J.; Han, R. Efficacy of entomopathogenic Steinernema and Heterorhabditis nematodes against Holotrichia oblita. J. Pest Sci. 2015, 88, 359–368. [Google Scholar] [CrossRef]
- Kapranas, A.; Malone, B.; Quinn, S.; Mc Namara, L.; Williams, C.D.; O’Tuama, P.; Peters, A.; Griffin, C.T.; O’Tuama, P.; Peters, A.; et al. Efficacy of entomopathogenic nematodes for control of large pine weevil, Hylobius abietis: Effects of soil type, pest density and spatial distribution. Pest Sci. 2016, 90, 495–505. [Google Scholar] [CrossRef]
- Antwi, F.B.; Reddy, G.V.P. Efficacy of entomopathogenic nematodes and sprayable polymer gel against crucifer flea beetle (Coleoptera: Chrysomelidae) on canola. J. Econ. Entomol. 2016, 109, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Sáenz-Aponte, A. Susceptibility variation to different entomopathogenic nematodes in Strategus aloeus L (Coleoptera: Scarabaeidae). Springerplus 2015, 4, 620. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, D.D.O.; Gomes, V.M.; Dolinski, C.; Souza, R.M. Potential of entomopathogenic nematodes as biocontrol agents of immature stages of Aedes aegypti. Nematoda 2015, 2, e092015. [Google Scholar] [CrossRef]
- Wu, H.; Gong, Q.; Fan, K.; Sun, R.; Xu, Y.; Zhang, K. Synergistic effect of entomopathogenic nematodes and thiamethoxam in controlling Bradysia odoriphaga Yang and Zhang (Diptera: Sciaridae). Biol. Control 2017, 111, 53–60. [Google Scholar] [CrossRef]
- Edmunds, C.V.; Wilding, C.S.; Rae, R. Susceptibility of Chironomus plumosus larvae (Diptera: Chironomidae) to entomopathogenic nematodes (Rhabditidae: Steinernematidae and Heterorhabditidae): Potential for control. Eur. J. Entomol. 2017, 114, 526–532. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Audsley, N. Further screening of entomopathogenic fungi and nematodes as control agents for Drosophila suzukii. Insects 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Garriga, A.; Morton, A.; Garcia-del-Pino, F. Is Drosophila suzukii as susceptible to entomopathogenic nematodes as Drosophila melanogaster? J. Pest Sci. 2018, 91, 789–798. [Google Scholar] [CrossRef]
- Hübner, A.; Englert, C.; Herz, A. Effect of entomopathogenic nematodes on different developmental stages of Drosophila suzukii in and outside fruits. BioControl 2017, 62, 669–680. [Google Scholar] [CrossRef]
- Archana, M.; D’Souza, P.E.; Patil, J. Efficacy of entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) on developmental stages of house fly, Musca domestica. J. Parasit. Dis. 2017, 41, 782–794. [Google Scholar] [CrossRef] [PubMed]
- de Souza Rodrigues Leal, L.C.; de Oliveira Monteiro, C.M.; de Mendonca, A.E.; Bittencourt, V.R.E.P.; Bittencourt, A.J.; Rodrigues, L.C.; de Oliveira, C.; de Mendonca, A.E.; Pinheiro, V.R.; Bittencourt, A.J. Potential of entomopathogenic nematodes of the genus Heterorhabditis for the control of Stomoxys calcitrans (Diptera: Muscidae). Brazilian J. Vet. Parasitol. 2017, 26, 451–456. [Google Scholar] [CrossRef]
- Berkvens, N.; Van Vaerenbergh, J.; Maes, M.; Beliën, T.; Viaene, N. Entomopathogenic nematodes fail to parasitize the woolly apple aphid Eriosoma lanigerum as their symbiotic bacteria are suppressed. J. Appl. Entomol. 2014, 138, 644–655. [Google Scholar] [CrossRef]
- Le Vieux, P.D.; Malan, A.P. An overview of the vine mealybug (Planococcus ficus) in South African vineyards and the use of entomopathgenic nematodes as potential biocontrol agent. S. Afr. J. Enol. Vitic. 2013, 34, 108–118. [Google Scholar]
- Wagutu, G.K.; Kang’ethe, L.N.; Waturu, C.N. Efficacy of entomopathogenic nematode (Steinernema karii) in control of termites (Coptorermes formosanus). J. Agric. Sci. Technol. 2017, 18, 55–64. [Google Scholar]
- Zadji, L.; Baimey, H.; Afouda, L.; Moens, M.; Decraemer, W. Comparative susceptibility of Macrotermes bellicosus and Trinervitermes occidentalis (Isoptera: Termitidae) to entomopathogenic nematodes from Benin. Nematology 2014, 16, 719–727. [Google Scholar] [CrossRef]
- Odendaal, D.; Addison, M.F.; Malan, A.P. Evaluation of above-ground application of entomopathogenic nematodes for the control of diapausing codling moth (Cydia pomonella L.) under natural conditions. Afr. Entomol. 2016, 24, 61–74. [Google Scholar] [CrossRef]
- Ramos-Rodríguez, O.; Campbell, J.F.; Ramaswamy, S.B. Pathogenicity of three species of entomopathogenic nematodes to some major stored-product insect pests. J. Stored Prod. Res. 2006, 42, 241–252. [Google Scholar] [CrossRef]
- Bolaños, T.A.; Gutiérrez, G.A.M.; Tinoco, C.E.; Cortés-martínez, C.I.; Sánchez, D.M. Loss caused by fruit worm and its treatment with entomopathogenic nematodes in fruits of Physalis ixocarpa Guenee. Int. J. Appl. Microbiol. Biotechnol. Res. 2016, 4, 60–65. [Google Scholar]
- Sunanda, B.S.; Jeyakumar, P.; Jacob, V. V Bioefficacy of different formulations of entomopathogenic nematode Steinernema carpocapsae against Diamond back moth (Plutella xylostella) infesting Cabbage (Brassica oleracea var. capitata). J. Biopestic. 2014, 7, 210–215. [Google Scholar]
- Safdar, H.; Javed, N.; Khan, S.A.; Arshad, M. Reproduction potential of entomopathogenic nematodes on armyworm (Spodoptera litura). Pak. J. Zool. 2018, 50, 1–4. [Google Scholar] [CrossRef]
- Karabörklü, S.; Ayvaz, A.; Yilmaz, S.; Azizoglu, U.; Akbulut, M. Native entomopathogenic nematodes isolated from Turkey and their effectiveness on pine processionary moth, Thaumetopoea wilkinsoni Tams. Int. J. Pest Manag. 2015, 61, 3–8. [Google Scholar] [CrossRef]
- Salari, E.; Karimi, J.; Sadeghi-Nameghi, H.; Hosseini, M. Efficacy of two entomopathogenic nematodes Heterorhabditis bacteriophora and Steinernema carpocapsae for control of the leopard moth borer Zeuzera pyrina (Lepidoptera: Cossidae) larvae under laboratory conditions. Biocontrol Sci. Technol. 2015, 25, 260–275. [Google Scholar] [CrossRef]
- Malan, A.P.; Knoetze, R.; Tiedt, L. Heterorhabditis noenieputensis n. sp. (Rhabditida: Heterorhabditidae), a new entomopathogenic nematode from South Africa. J. Helminthol. 2014, 88, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Shahina, F.; Tabassum, K.A.; Salma, J.; Mehreen, G.; Knoetze, R. Heterorhabditis pakistanense n. sp. (Nematoda: Heterorhabditidae) a new entomopathogenic nematode from Pakistan. J. Helminthol. 2017, 91, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, S.; Khanum, T.A.; Ali, S.; Solangi, G.S.; Gulsher, M.; Javed, S. Steinernema balochiense n. sp. (Rhabditida: Steinernematidae) a new entomopathogenic nematode from Pakistan. Zootaxa 2015, 3904, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Çimen, H.; Půža, V.; Nermuť, J.; Hatting, J.; Ramakuwela, T.; Faktorová, L.; Hazir, S. Steinernema beitlechemi n. sp., a new entomopathogenic nematode (Nematoda: Steinernematidae) from South Africa. Nematology 2016, 18, 439–453. [Google Scholar] [CrossRef]
- Çimen, H.; Puza, V.; Nermut, J.; Hatting, J.; Ramakuwela, T.; Hazir, S. Steinernema biddulphi n. sp., a new entomopathogenic nematode (Nematoda: Steinernematidae) from South Africa. J. Nematol. 2016, 48, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Çimen, H.; Lee, M.M.; Hatting, J.; Hazir, S.; Stock, S.P. Steinernema innovationi n. sp. (Panagrolaimomorpha: Steinernematidae), a new entomopathogenic nematode species from South Africa. J. Helminthol. 2015, 89, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Mráček, Z.; Nermuť, J. Steinernema poinari sp. n. (Nematoda: Steinernematidae) a new entomopathogenic nematode from the Czech Republic. Zootaxa 2014, 3760, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Půža, V.; Nermut, J.; Mráček, Z.; Gengler, S.; Haukeland, S. Steinernema pwaniensis n. sp., a new entomopathogenic nematode (Nematoda: Steinernematidae) from Tanzania. J. Helminthol. 2017, 91, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Gorgadze, O.; Lortkhipanidze, M.; Ogier, J.-C.; Tailliez, P.; Burjanadze, M. Steinernema tbilisiensis sp. n. (Nematoda: Steinernematidae)—A new species of entomopathogenic nematode from Georgia. J. Agric. Sci. Technol. A 2015, 5, 264–276. [Google Scholar] [CrossRef]
- Stock, S.P. Steinernema tophus sp. n. (Nematoda: Steinernematidae), a new entomopathogenic nematode from South Africa. Zootaxa 2014, 3821, 337–353. [Google Scholar]
- Shapiro-Ilan, D.I.; Gaugler, R. Production technology for entomopathogenic nematodes and their bacterial symbionts. J. Ind. Microbiol. Biotechnol. 2002, 28, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Sumaya, N.H.; Gohil, R.; Okolo, C.; Addis, T.; Doerfler, V.; Ehlers, R.U.; Molina, C. Applying inbreeding, hybridization and mutagenesis to improve oxidative stress tolerance and longevity of the entomopathogenic nematode Heterorhabditis bacteriophora. J. Invertebr. Pathol. 2018, 181, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Ow, M.C.; Borziak, K.; Nichitean, A.M.; Dorus, S.; Hall, S.E. Early experiences mediate distinct adult gene expression and reproductive programs in Caenorhabditis elegans. PLoS Genet. 2018, 14, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Grewal, P.S.; Peters, A. Formulation and quality. In Nematodes as Biocontrol Agents; Grewal, P.S., Ehlers, R.U., Shapiro-Ilan, D.I., Eds.; CAB International: Wallingford, UK, 2005; pp. 79–90. [Google Scholar]
- Patel, M.N.; Stolinski, M.; Wright, D.J. Neutral lipids and the assessment of infectivity in entomopathogenic nematodes: Observations on four Steinernema species. Parasitology 1997, 114, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.N.; Wright, D.J. Glycogen: Its importance in the infectivity of aged juveniles of Steinernema carpocapsae. Parasitology 1997, 114, 591–596. [Google Scholar] [PubMed]
- Menti, H.; Wright, D.J.; Perry, R.N. Infectivity of populations of the entomopathogenic nematodes Steinernema feltiae and Heterorhabditis megidis in relation to temperature, age and lipid content. Nematology 2000, 2, 515–521. [Google Scholar] [CrossRef]
- Lewis, E.E.; Campbell, J.; Griffin, C.; Kaya, H.; Peters, A. Behavioral ecology of entomopathogenic nematodes. Biol. Control 2006, 38, 66–79. [Google Scholar] [CrossRef]
- Griffin, C.T. Behaviour and population dynamics of entomopathogenic nematodes following application. In Nematode Pathogenesis of Insects and Other Pests: Ecology and Applied Technologies for Sustainable Plant and Crop Protection; Springer: Berlin, Germany, 2015; pp. 57–95. ISBN 9783319182667. [Google Scholar]
- Dillman, A.R.; Guillermin, M.L.; Ha, J.; Kim, B.; Sternberg, P.W.; Hallem, E.A. Olfaction shapes host—Parasite interactions in parasitic nematodes. Proc. Natl. Acad. Sci. USA 2012, 109, E2324–E2333. [Google Scholar] [CrossRef] [PubMed]
- Torr, P.; Heritage, S.; Wilson, M.J. Vibrations as a novel signal for host location by parasitic nematodes. Int. J. Parasitol. 2004, 34, 997–999. [Google Scholar] [CrossRef] [PubMed]
- Burman, M.; Pye, A.E. Neoaplectana carpocapsae: Movements of nematode populations on a thermal gradient. Exp. Parasitol. 1980, 49, 258–265. [Google Scholar] [CrossRef]
- Byers, J.A.; Poinar, G.O. Location of insect hosts by the nematode, Neoaplectana carpocapsae, in response to temperature. Behaviour 1982, 79, 1–10. [Google Scholar] [CrossRef]
- Campbell, J.F.; Gaugler, R. Nictation behaviour and its ecological implications in the host search strategies of entomopathogenic nematodes (Hererorhabditidae and Steinernematidae). Behaviour 1993, 126, 155–169. [Google Scholar] [CrossRef]
- Campbell, J.F.; Kaya, H.K. How and why a parasitic nematode jumps. Nature 1999, 397, 485–486. [Google Scholar] [CrossRef]
- Campbell, J.F.; Kaya, H.K. Variation in entomopathogenic nematode (Steinernematidae and Heterorhabditidae) infective-stage jumping behaviour. Nematology 2002, 4, 471–482. [Google Scholar] [CrossRef]
- Bal, H.K.; Grewal, P.S. Lateral dispersal and foraging behavior of entomopathogenic nematodes in the absence and presence of mobile and non-mobile hosts. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.K.; Taylor, R.A.J.; Grewal, P.S. Ambush foraging entomopathogenic nematodes employ “sprinters” for long-distance dispersal in the absence of hosts. J. Parasitol. 2014, 100, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.K.; Michel, A.P.; Grewal, P.S. Genetic selection of the ambush foraging entomopathogenic nematode, Steinernema carpocapsae for enhanced dispersal and its associated trade-offs. Evol. Ecol. 2014, 28, 923–939. [Google Scholar] [CrossRef]
- Kapranas, A.; Malone, B.; Quinn, S.; O’Tuama, P.; Peters, A.; Griffin, C.T.; Tuama, P.O.; Peters, A.; Griffin, C.T. Optimizing the application method of entomopathogenic nematode suspension for biological control of large pine weevil Hylobius abietis. BioControl 2017, 62, 659–667. [Google Scholar] [CrossRef]
- Bal, H.K.; Acosta, N.; Cheng, Z.; Grewal, P.S.; Hoy, C.W. Effect of habitat and soil management on dispersal and distribution patterns of entomopathogenic nematodes. Appl. Soil Ecol. 2017, 121, 48–59. [Google Scholar] [CrossRef]
- Kapranas, A.; Maher, A.M.D.; Griffin, C.T. The influence of organic matter content and media compaction on the dispersal of entomopathogenic nematodes with different foraging strategies. Parasitology 2017, 144, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Jaffuel, G.; Blanco-Pérez, R.; Büchi, L.; Mäder, P.; Fließbach, A.; Charles, R.; Degen, T.; Turlings, T.C.J.; Campos-Herrera, R. Effects of cover crops on the overwintering success of entomopathogenic nematodes and their antagonists. Appl. Soil Ecol. 2017, 114, 62–73. [Google Scholar] [CrossRef]
- Abate, B.A.; Slippers, B.; Wingfield, M.J.; Malan, A.P.; Hurley, B.P. Diversity of entomopathogenic nematodes and their symbiotic bacteria in south African plantations and indigenous forests. Nematology 2018. [Google Scholar] [CrossRef]
- Ramos Mertz, N.; Agudelo, E.J.G.; Sales, F.S.; Rohde, C.; Moino, A. Phoretic dispersal of the entomopathogenic nematode Heterorhabditis amazonensis by the beetle Calosoma granulatum. Phytoparasitica 2014, 42, 179–187. [Google Scholar] [CrossRef]
- El-Borai, F.E.; Campos-Herrera, R.; Stuart, R.J.; Duncan, L.W. Substrate modulation, group effects and the behavioral responses of entomopathogenic nematodes to nematophagous fungi. J. Invertebr. Pathol. 2011, 106, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Lewis, E.E.; Schliekelman, P. Aggregative group behavior in insect parasitic nematode dispersal. Int. J. Parasitol. 2014, 44, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.B.; Shapiro-Ilan, D.; Lewis, E.E.; Kaplan, F.; Alborn, H.; Gu, X.H.; Schliekelman, P. Movement patterns in entomopathogenic nematodes: Continuous vs. temporal. J. Invertebr. Pathol. 2018, 151, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Hallem, E.A.; Dillman, A.R.; Hong, A.V.; Zhang, Y.; Yano, J.M.; Demarco, S.F.; Sternberg, P.W. A sensory code for host seeking in parasitic nematodes. Curr. Biol. 2011, 21, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Rivera, M.J.; Martini, X.; Khrimian, A.; Stelinski, L. A weevil sex pheromone serves as an attractant for its entomopathogenic nematode predators. Chemoecology 2017, 27, 199–206. [Google Scholar] [CrossRef]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 2005, 434, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Turlings, T.C.J.; Hiltpold, I.; Rasmann, S. The importance of root-produced volatiles as foraging cues for entomopathogenic nematodes. Plant Soil 2012, 358, 51–60. [Google Scholar] [CrossRef]
- Laznik, Ž.; Trdan, S. Attraction behaviors of entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) to synthetic volatiles emitted by insect damaged potato tubers. J. Pest Sci. 2016, 42, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Le Vieux, P.D.; Malan, A.P. Prospects for using entomopathogenic nematodes to control the vine mealybug, Planococcus ficus, in South African vineyards. S. Afr. J. Enol. Vitic. 2015, 36, 59–70. [Google Scholar] [CrossRef]
- Tonelli, M.; Peñaflor, M.F.G.V.; Leite, L.G.; Silva, W.D.; Martins, F.; Bento, J.M.S. Attraction of entomopathogenic nematodes to sugarcane root volatiles under herbivory by a sap-sucking insect. Chemoecology 2016, 26, 59–66. [Google Scholar] [CrossRef]
- Li, C.; Wang, Y.; Hu, Y.; Hua, C.; Wang, C. Three dimensional study of wounded plant roots recruiting entomopathogenic nematodes with Pluronic gel as a medium. Biol. Control 2015, 89, 68–74. [Google Scholar] [CrossRef]
- Ennis, D.E.; Dillon, A.B.; Griffin, C.T. Simulated roots and host feeding enhance infection of subterranean insects by the entomopathogenic nematode Steinernema carpocapsae. J. Invertebr. Pathol. 2010, 103, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Demarta, L.; Hibbard, B.E.; Bohn, M.O.; Hiltpold, I. The role of root architecture in foraging behavior of entomopathogenic nematodes. J. Invertebr. Pathol. 2014, 122, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Chiriboga, M.X.; Campos-Herrera, R.; Jaffuel, G.; Röder, G.; Turlings, T.C.J. Diffusion of the maize root signal (E)-β-caryophyllene in soils of different textures and the effects on the migration of the entomopathogenic nematode Heterorhabditis megidis. Rhizosphere 2017, 3, 53–59. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Willett, D.S.; Pereira, R.V.; de Sabino, P.H.S.; Junior, A.M.; Pareja, M.; Dickson, D.W. Parameters affecting plant defense pathway mediated recruitment of entomopathogenic nematodes. Biocontrol Sci. Technol. 2017, 27, 833–843. [Google Scholar] [CrossRef]
- Willett, D.S.; Alborn, H.T.; Duncan, L.W.; Stelinski, L.L. Social Networks of Educated Nematodes. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.D.; Dillon, A.B.; Ennis, D.E.; Hennessy, R.; Griffin, C.T. Differential susceptibility of pine weevil, Hylobius abietis (Coleoptera: Curculionidae), larvae and pupae to entomopathogenic nematodes and death of adults infected as pupae. BioControl 2015, 60, 537–546. [Google Scholar] [CrossRef]
- Blanco-Pérez, R.; Bueno-Pallero, F.Á.; Neto, L.; Campos-Herrera, R. Reproductive efficiency of entomopathogenic nematodes as scavengers. Are they able to fight for insect’s cadavers? J. Invertebr. Pathol. 2017, 148. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Shokal, U.; Yadav, S.; Kenney, E.; Maldonado, T. Insect immunity to entomopathogenic nematodes and their mutualistic bacteria. Curr. Top. Microbiol. Immunol. 2017, 402, 123–156. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Gao, A.; Li, B.; Wang, M.; Shan, L. Two symbiotic bacteria of the entomopathogenic nematode Heterorhabditis spp. against Galleria mellonella. Toxicon 2017, 127, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Peña, J.M.; Carrillo, M.A.; Hallem, E.A. Variation in the susceptibility of Drosophila to different entomopathogenic nematodes. Infect. Immun. 2015, 83, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Macchietto, M.; Chang, D.; Barros, M.M.; Baldwin, J.; Mortazavi, A.; Dillman, A.R. Activated entomopathogenic nematode infective juveniles release lethal venom proteins. PLoS Pathog. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bisch, G.; Pagès, S.; McMullen, J.G.; Stock, S.P.; Duvic, B.; Givaudan, A.; Gaudriault, S. Xenorhabdus bovienii CS03, the bacterial symbiont of the entomopathogenic nematode Steinernema weiseri, is a non-virulent strain against lepidopteran insects. J. Invertebr. Pathol. 2015, 124, 15–22. [Google Scholar] [CrossRef] [PubMed]
- McMullen, J.G.; Peterson, B.F.; Forst, S.; Blair, H.G.; Stock, S.P. Fitness costs of symbiont switching using entomopathogenic nematodes as a model. BMC Evol. Biol. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Cowles, R.S.; Cowles, E.A.; Gaugler, R.; Cox-Foster, D.L. Relationship between the successful infection by entomopathogenic nematodes and the host immune response. Int. J. Parasitol. 2007, 37, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Alvandi, J.; Karimi, J.; Dunphy, G.B. Cellular reactions of the white grub larvae, Polyphylla adspersa, against entomopathogenic nematodes. Nematology 2014, 16, 1047–1058. [Google Scholar] [CrossRef]
- Rahatkhah, Z.; Karimi, J.; Ghadamyari, M.; Brivio, M.F. Immune defenses of Agriotes lineatus larvae against entomopathogenic nematodes. BioControl 2015, 60, 641–653. [Google Scholar] [CrossRef]
- Lalitha, K.; Karthi, S.; Vengateswari, G.; Karthikraja, R.; Perumal, P.; Shivakumar, M.S. Effect of entomopathogenic nematode of Heterorhabditis indica infection on immune and antioxidant system in lepidopteran pest Spodoptera litura (Lepidoptera: Noctuidae). J. Parasit. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.A.M.; Taha, M.A.; Salem, H.H.A. Changes in enzyme activities in Agrotis ipsilon (Lepidoptera, noctuidae) as a response to entomopathogenic nematode infection. Int. J. Adv. Res. 2015, 3, 111–118. [Google Scholar]
- Ebrahimi, L.; Shiri, M.; Dunphy, G.B. Effect of entomopathogenic nematode, Steinernema feltiae, on survival and plasma phenoloxidase activity of Helicoverpa armigera (Hb) (Lepidoptera: Noctuidae) in laboratory conditions. Egypt. J. Biol. Pest Control 2018, 28, 12. [Google Scholar] [CrossRef]
- Binda-Rossetti, S.; Mastore, M.; Protasoni, M.; Brivio, M.F. Effects of an entomopathogen nematode on the immune response of the insect pest red palm weevil: Focus on the host antimicrobial response. J. Invertebr. Pathol. 2016, 133, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Kenney, E.; Eleftherianos, I. Entomopathogenic and plant pathogenic nematodes as opposing forces in agriculture. Int. J. Parasitol. 2016, 46, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Helmberger, M.S.; Shields, E.J.; Wickings, K.G. Ecology of belowground biological control: Entomopathogenic nematode interactions with soil biota. Appl. Soil Ecol. 2017, 121, 201–213. [Google Scholar] [CrossRef]
- Harvey, C.D.; Williams, C.D.; Dillon, A.B.; Griffin, C.T. Inundative pest control: How risky is it? A case study using entomopathogenic nematodes in a forest ecosystem. For. Ecol. Manag. 2016, 380, 242–251. [Google Scholar] [CrossRef]
- Babendreier, D.; Jeanneret, P.; Pilz, C.; Toepfer, S. Non-target effects of insecticides, entomopathogenic fungi and nematodes applied against western corn rootworm larvae in maize. J. Appl. Entomol. 2015, 139, 457–467. [Google Scholar] [CrossRef]
- Metwally, H.M.; Sabry, A.K.H.; Gaber, N.M. Biosafety of different entomopathogenic nematodes species on some insects natural enemies. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 1878–1883. [Google Scholar]
- Dutka, A.; McNulty, A.; Williamson, S.M. A new threat to bees? Entomopathogenic nematodes used in biological pest control cause rapid mortality in Bombus terrestris. PeerJ 2015, 3, e1413. [Google Scholar] [CrossRef] [PubMed]
- Shorter, J.R.; Rueppell, O. A review on self-destructive defense behaviors in social insects. Insectes Soc. 2012, 59, 1–10. [Google Scholar] [CrossRef]
- Munday, Z.; Brown, M.J.F. Bring out your dead: Quantifying corpse removal in Bombus terrestris, an annual eusocial insect. Anim. Behav. 2018, 138, 51–57. [Google Scholar] [CrossRef]
- Harvey, C.D.; Griffin, C.T. Local host-dependent persistence of the entomopathogenic nematode Steinernema carpocapsae used to control the large pine weevil Hylobius abietis. BioControl 2016, 61, 185–193. [Google Scholar] [CrossRef]
- Gulcu, B.; Hazir, S.; Kaya, H.K. Scavenger deterrent factor (SDF) from symbiotic bacteria of entomopathogenic nematodes. J. Invertebr. Pathol. 2012, 110, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Kaya, H.K.; Heungens, K.; Goodrich-Blair, H. Response of ants to a deterrent factor(s) produced by the symbiotic bacteria of entomopathogenic nematodes. Appl. Environ. Microbiol. 2002, 68, 6202–6209. [Google Scholar] [CrossRef] [PubMed]
- Fenton, A.; Magoolagan, L.; Kennedy, Z.; Spencer, K.A. Parasite-induced warning coloration: A novel form of host manipulation. Anim. Behav. 2011, 81, 417–422. [Google Scholar] [CrossRef]
- Raja, R.K.; Aiswarya, D.; Gulcu, B.; Raja, M.; Perumal, P.; Sivaramakrishnan, S.; Kaya, H.K.; Hazir, S. Response of three cyprinid fish species to the Scavenger Deterrent Factor produced by the mutualistic bacteria associated with entomopathogenic nematodes. J. Invertebr. Pathol. 2017, 143, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Mertz, N.R.; Agudelo, E.J.G.; Sales, F.S.; Moino Junior, A. Effects of entomopathogenic nematodes on the predator Calosoma granulatum in the laboratory. J. Insect Behav. 2015, 28, 312–327. [Google Scholar] [CrossRef]
- Ulug, D.; Hazir, S.; Kaya, H.K.; Lewis, E. Natural enemies of natural enemies: The potential top-down impact of predators on entomopathogenic nematode populations. Ecol. Entomol. 2014, 39, 462–469. [Google Scholar] [CrossRef]
- Dillon, A.B.; Moore, C.P.; Downes, M.J.; Griffin, C.T. Evict or infect? Managing populations of the large pine weevil, Hylobius abietis, using a bottom-up and top-down approach. For. Ecol. Manag. 2008, 255, 2634–2642. [Google Scholar] [CrossRef]
- Mbata, G.N.; Shapiro-Ilan, D.I. Compatibility of Heterorhabditis indica (Rhabditida: Heterorhabditidae) and Habrobracon hebetor (Hymenoptera: Braconidae) for biological control of Plodia interpunctella (Lepidoptera: Pyralidae). Biol. Control 2010, 54, 75–82. [Google Scholar] [CrossRef]
- Heve, W.K.; El-Borai, F.E.; Carrillo, D.; Duncan, L.W. Increasing entomopathogenic nematode biodiversity reduces efficacy against the Caribbean fruit fly Anastrepha suspensa: Interaction with the parasitoid Diachasmimorpha longicaudata. J. Pest Sci. 2018, 91, 799–813. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Půža, V.; Jaffuel, G.; Blanco-Pérez, R.; Čepulyte-Rakauskiene, R.; Turlings, T.C.J. Unraveling the intraguild competition between Oscheius spp. nematodes and entomopathogenic nematodes: Implications for their natural distribution in Swiss agricultural soils. J. Invertebr. Pathol. 2015, 132, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Campos-Herrera, R.; Jaffuel, G.; Chiriboga, X.; Blanco-Pérez, R.; Fesselet, M.; Půža, V.; Mascher, F.; Turlings, T.C.J. Traditional and molecular detection methods reveal intense interguild competition and other multitrophic interactions associated with native entomopathogenic nematodes in Swiss tillage soils. Plant Soil 2015, 389, 237–255. [Google Scholar] [CrossRef]
- O’Callaghan, K.M.; Zenner, A.N.R.L.; Hartley, C.J.; Griffin, C.T. Interference competition in entomopathogenic nematodes: Male Steinernema kill members of their own and other species. Int. J. Parasitol. 2014, 44, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Kapranas, A.; Maher, A.M.D.; Griffin, C.T. Higher relatedness mitigates mortality in a nematode with lethal male fighting. J. Evol. Biol. 2016, 29, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Javed, N.; Kamran, M.; Abbas, H.; Safdar, A.; Ul Haq, I. Management of Meloidogyne incognita race 1 through the use of entomopathogenic nematodes in tomato. Pak. J. Zool. 2016, 48, 763–768. [Google Scholar]
- Kepenekci, I.; Hazir, S.; Lewis, E.E. Evaluation of entomopathogenic nematodes and the supernatants of the in vitro culture medium of their mutualistic bacteria for the control of the root-knot nematodes Meloidogyne incognita and M. arenaria. Pest Manag. Sci. 2016, 72, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Aatif, H.M.; Javed, N.; Khan, S.A.; Ahmed, S.; Raheel, M. Virulence of entomopathogenic nematodes against Meloidogyne incognita for invasion, development and reproduction at different application times in brinjal roots. Int. J. Agric. Biol. 2015, 17, 995–1000. [Google Scholar] [CrossRef]
- An, R.; Orellana, D.; Phelan, L.P.; Cañas, L.; Grewal, P.S. Entomopathogenic nematodes induce systemic resistance in tomato against Spodoptera exigua, Bemisia tabaci and Pseudomonas syringae. Biol. Control 2016, 93, 24–29. [Google Scholar] [CrossRef]
- Hazir, S.; Shapiro-Ilan, D.I.; Hazir, C.; Leite, L.G.; Cakmak, I.; Olson, D. Multifaceted effects of host plants on entomopathogenic nematodes. J. Invertebr. Pathol. 2016, 135, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S.; Olmstead, D.; Tian, J.-C.; Collins, H.L.; Shelton, A.M. Tri-trophic studies using Cry1Ac-resistant Plutella xylostella demonstrate no adverse effects of Cry1Ac on the entomopathogenic nematode, Heterorhabditis bacteriophora. J. Econ. Entomol. 2014, 107, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Frazer, J.; Banga, A.; Pruitt, K.; Harsh, S.; Jaenike, J.; Eleftherianos, I. Endosymbiont-based immunity in Drosophila melanogaster against parasitic nematode infection. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Daugherty, S.; Shetty, A.C.; Eleftherianos, I. RNAseq analysis of the drosophila response to the entomopathogenic nematode Steinernema. G3 Genes Genomes Genet. 2017, 7, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Vadnal, J.; Ratnappan, R.; Keaney, M.; Kenney, E.; Eleftherianos, I.; O’Halloran, D.; Hawdon, J.M. Identification of candidate infection genes from the model entomopathogenic nematode Heterorhabditis bacteriophora. BMC Genomics 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Arefin, B.; Kucerova, L.; Dobes, P.; Markus, R.; Strnad, H.; Wang, Z.; Hyrsl, P.; Zurovec, M.; Theopold, U. Genome-wide transcriptional analysis of Drosophila larvae infected by entomopathogenic nematodes shows involvement of complement, recognition and extracellular matrix proteins. J. Innate Immun. 2014, 6, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Půža, V.; Chundelová, D.; Nermuť, J.; Žurovcová, M.; Mráček, Z. Intra-individual variability of ITS regions in entomopathogenic nematodes (Steinernematidae: Nematoda): Implications for their taxonomy. BioControl 2015, 60, 547–554. [Google Scholar] [CrossRef]
- Padmanaban, K.; Karthik, R.R.; Razia, M.; Chellapandi, P.; Sivaramakrishnan, S. Genetic diversity of entomopathogenic nematodes in undisrupted ecosystem revealed with PCR-RAPD markers. Int. J. Nematol. 2014, 24, 2. [Google Scholar]
- Mandadi, N.; Hendrickson, C.; Handanahal, S.; Rajappa, T.; Pai, N.; Javeed, S.; Verghese, A.; Rai, A.; Pappu, A.; Nagaraj, G.; et al. Genome sequences of Photorhabdus luminescens strains isolated from entomopathogenic nematodes from southern India. Genomics Data 2015, 6, 46–47. [Google Scholar] [CrossRef] [PubMed]
- Salgado-morales, R.; Rivera-gómez, N.; Beltrán, L.F.L.; Hernández-mendoza, A.; Dantán-Gonzáles, E. Draft genome sequence of a Pseudomonas aeruginosa NA04 bacterium isolated from an entomopathogenic nematode. Genome Announc. 2017, 5, e00746-17. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Afrin, T.; Yoshida, M. Complete mitochondrial genomes of four entomopathogenic nematode species of the genus Steinernema. Parasit. Vectors 2016, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D.; Wood, P.L.; Burk, T.J.; Crawford, B.; Wright, S.M.; Adams, B.J. Evolution of virulence in Photorhabdus spp., entomopathogenic nematode symbionts. Syst. Appl. Microbiol. 2016, 39, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Santhi, V.S.; Ment, D.; Salame, L.; Soroker, V.; Glazer, I. Genetic improvement of host-seeking ability in the entomopathogenic nematodes Steinernema carpocapsae and Heterorhabditis bacteriophora toward the Red Palm Weevil Rhynchophorus ferrugineus. Biol. Control 2016, 100, 29–36. [Google Scholar] [CrossRef]
- Lu, D.; Baiocchi, T.; Dillman, A.R. Genomics of entomopathogenic nematodes and implications for pest control. Trends Parasitol. 2016, 32, 588–598. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).