1. Introduction
A large proportion of human population is exposed to infectious diseases spread by mosquitoes, including
Culex spp.,
Anopheles spp. and
Aedes spp., which are vectors of parasitic diseases [
1].
Culex quinquefasciatus Say is a vector of lymphatic filariasis. The disease is widely distributed in tropical regions, with around 150 million people infected, ranking filariasis as one of the main causes of global morbidity [
2].
Culex quinquefasciatus is present in tropical areas, a household pest in many urban settings, and widely reported as a vector resistant to registered insecticides [
3].
Tropical areas are more prone to vector-borne parasitic diseases and the risk has increased due to the intensification of globalization [
4]. Changes in climatic conditions due to global warming have promoted favorable conditions (temperature and humidity) for the development of mosquito populations, including hematophagous species. Poor sanitary conditions for water storage intended to household use, as well as unsanitary management of urban solid waste, can increase mosquito populations in urban and rural areas [
1].
On the other hand, the intensive use of synthetic insecticides in mosquito control programs has created resistance in the insect population [
5], undesirable effects on other insects [
6] and negative impacts on the environment [
7]. Although the use of insecticide is the main method in the global effort for mosquito control [
8], vector-borne diseases are persistent due to mosquitoes resistant to traditionally used insecticides [
9] and lack of effective vaccines [
10]. The resistance of mosquitoes to synthetic insecticides has led to an interest in natural products with potential insecticidal activity, especially those derived from plants, which are considered valid alternatives to conventional synthetic pesticides to control a variety of insect pests and vectors. In this context, the essential oils and their constituents have received much attention as potentially useful bioactive compounds against insects [
11]. The complex and variable mixtures of bioactive compounds with different modes of action, offered by plants, may lessen the chance of resistance in mosquito populations [
12].
Essential oils are heterogeneous mixtures of organic chemical compounds from different chemical families [
13]; many have a terpenoid backbone, especially monoterpenes and sesquiterpenes. Low molecular weight aliphatic compounds, acyclic esters or lactones may also be present [
14]. Chemical composition of essential oils is affected by factors such as plant species and subspecies, geographical location, harvest time, part of the plant used and the extraction methods used to obtain the essential oil [
15]. Several studies have reported the larvicidal action of medicinal plants and their essential oils against insect vectors of
Culex and
Anopheles genera, as well as
Aedes aegypti (L.) [
16,
17]. Furthermore, there are many secondary metabolites found in most of those plants that are considered to have insecticidal properties [
18,
19].
The aim of this paper is to determine, by laboratory bioassays, the larvicidal activity of seven essential oils on third instar larva and pupa of C. quinquefasciatus. Essential oils tested include cinnamon (Cinnamomum verum J. Presl), Mexican lime (Citrus aurantifolia Swingle) cumin (Cuminum cyminum Linnaeus), clove (Syzygium aromaticum (L.) Merr. & L.M. Perry), laurel (Laurus nobilis Linnaeus), Mexican oregano (Lippia berlandieri Schauer) and anise (Pimpinella anisum Linnaeus). Larvicidal effect and synergistic behavior of the major components present in the essential oils included in the study are also tested.
4. Discussion
Essential oils can be used as part of insecticides that can affect disease-related insect vectors, but with low impact on the accompanying insect fauna [
29,
30]. There are reports on essential oils with insecticidal and inhibitory oviposition capacity against
A. aegypti [
28]. The potential biological activity of the different essential oils varies according to plant species, its origin and its composition [
31,
32].
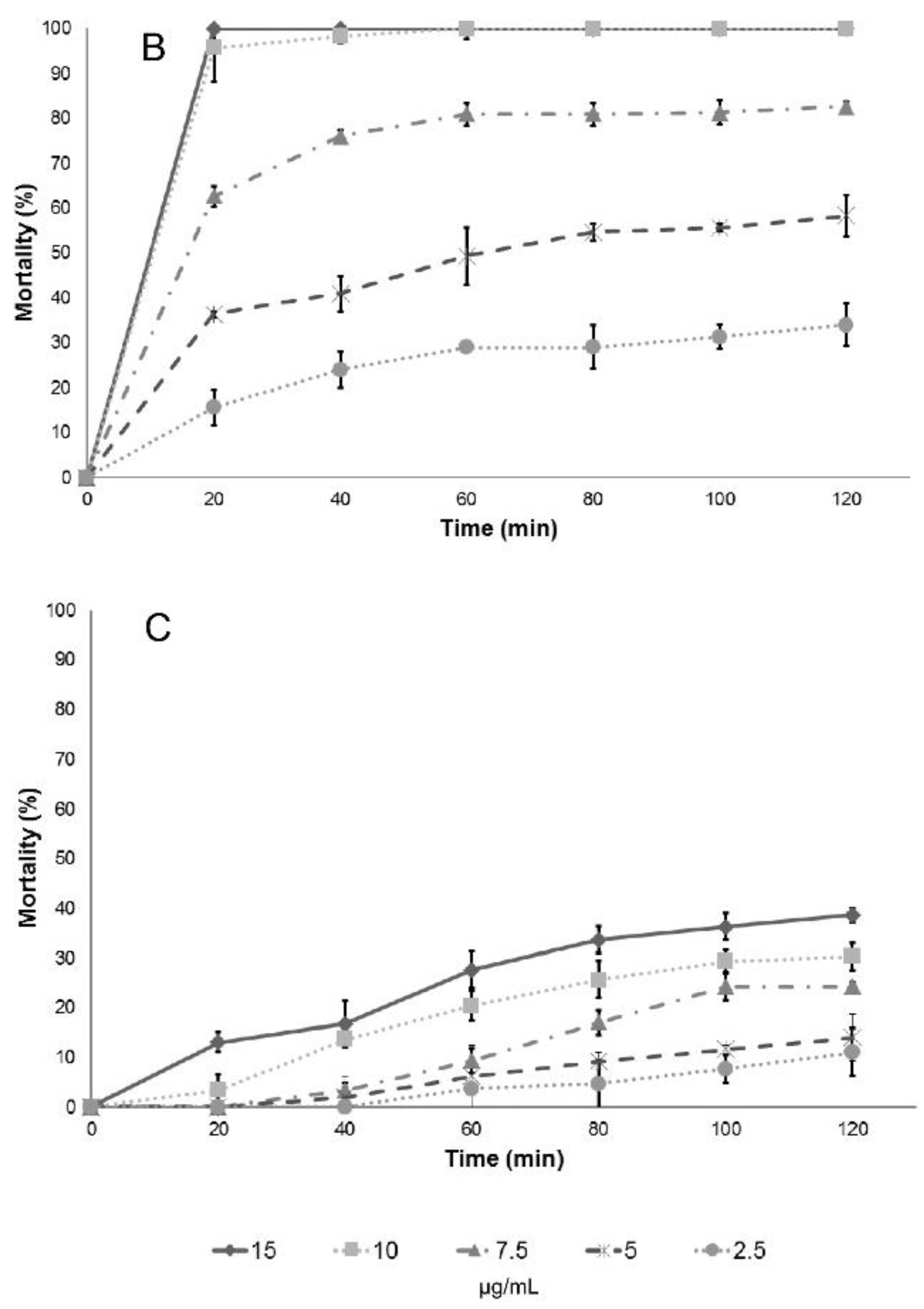
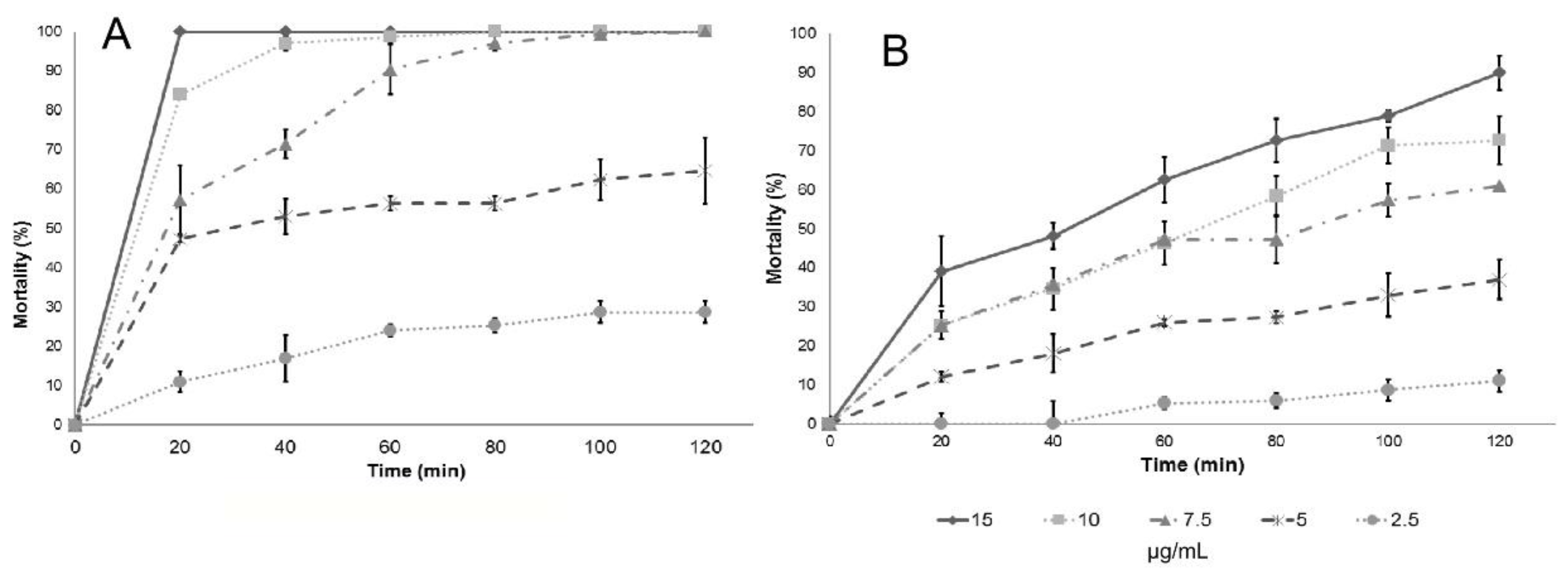
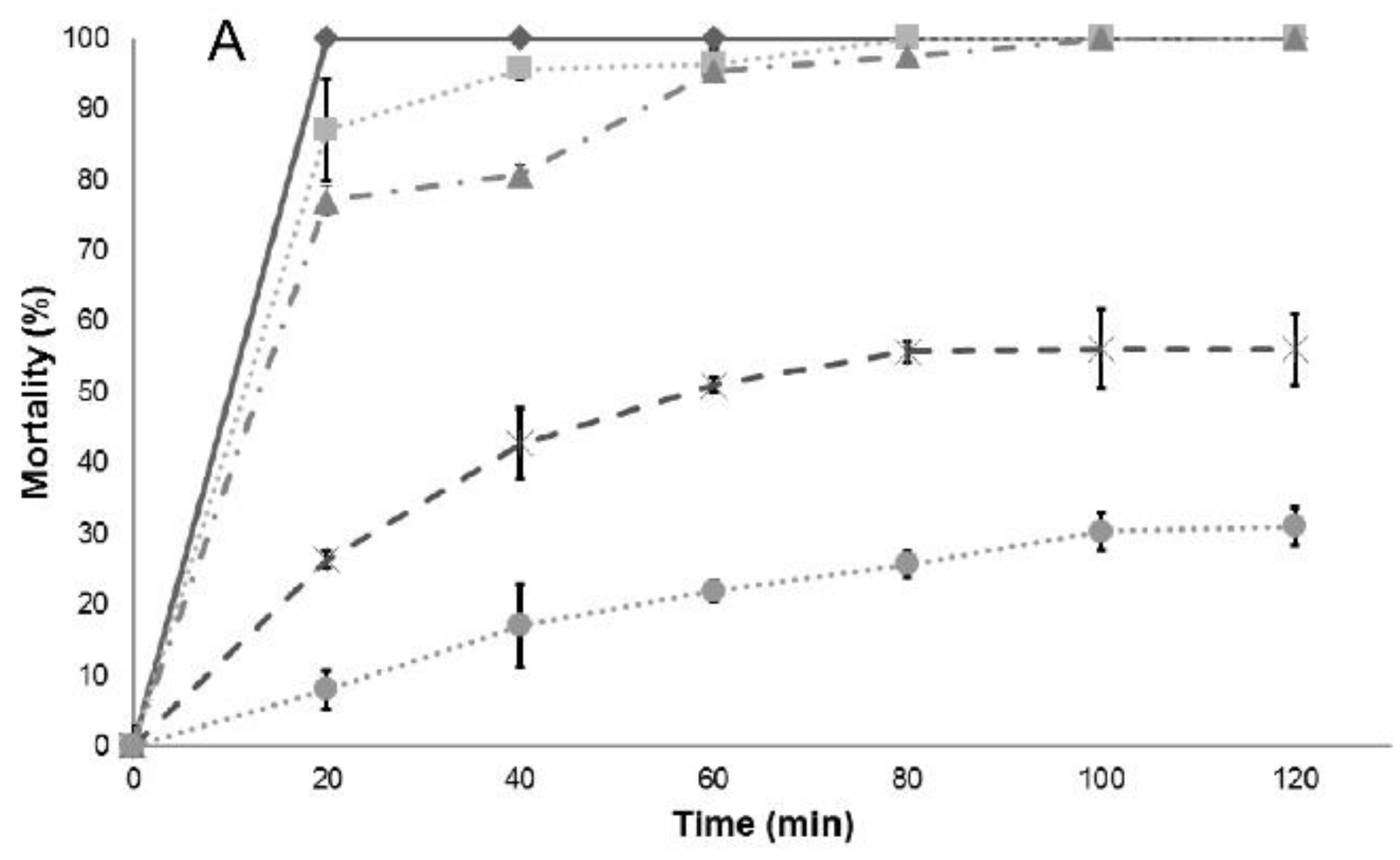
Regarding the larvicidal activities evaluated on instar III, there were no differences among the essential oils studied, except for the cinnamon essential oil (
C. verum J. Presl), which has a variation on LC
50 and LC
90 values. Results included in
Table 2 demonstrate that the essential oils tested can effectively control
C. quinquefasciatus pupae and larvae, and the effect can be the result of the interaction of the many compounds found in each essential oil, many in small or trace quantities, or can be attributed mainly to major components. To answer the latter question, the larvicidal and pupaecidal activity of the chemical compounds reported as major constituents of the essential oils tested were analyzed, and the results are shown in
Table 3.
Lethal concentration tests (LC
50 and LC
90) provide information on the concentration of a given compound to decrease a population by 50% and 90%, respectively. Usually the effect is not linear; therefore, it is important to determine both values. In the determination of those values, at least three replicates are needed for each concentration in the analysis, and this is the reason for the report of confidence intervals (provided in parenthesis in
Table 2 and
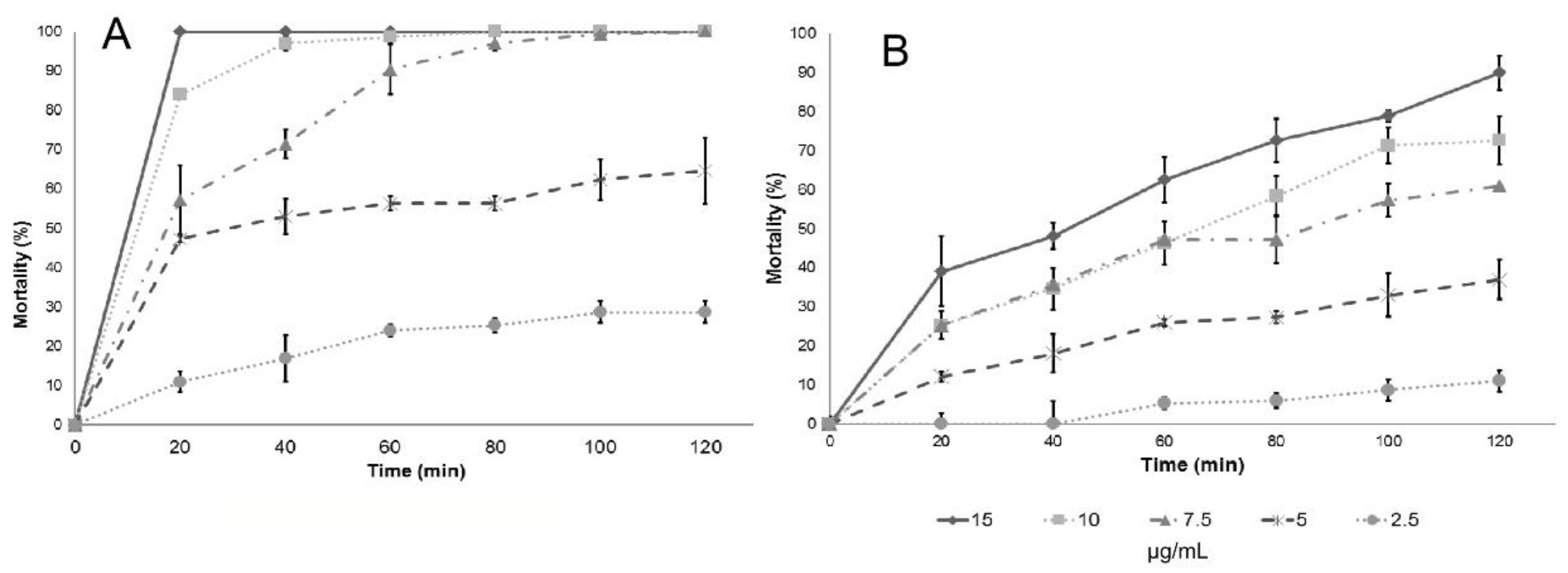
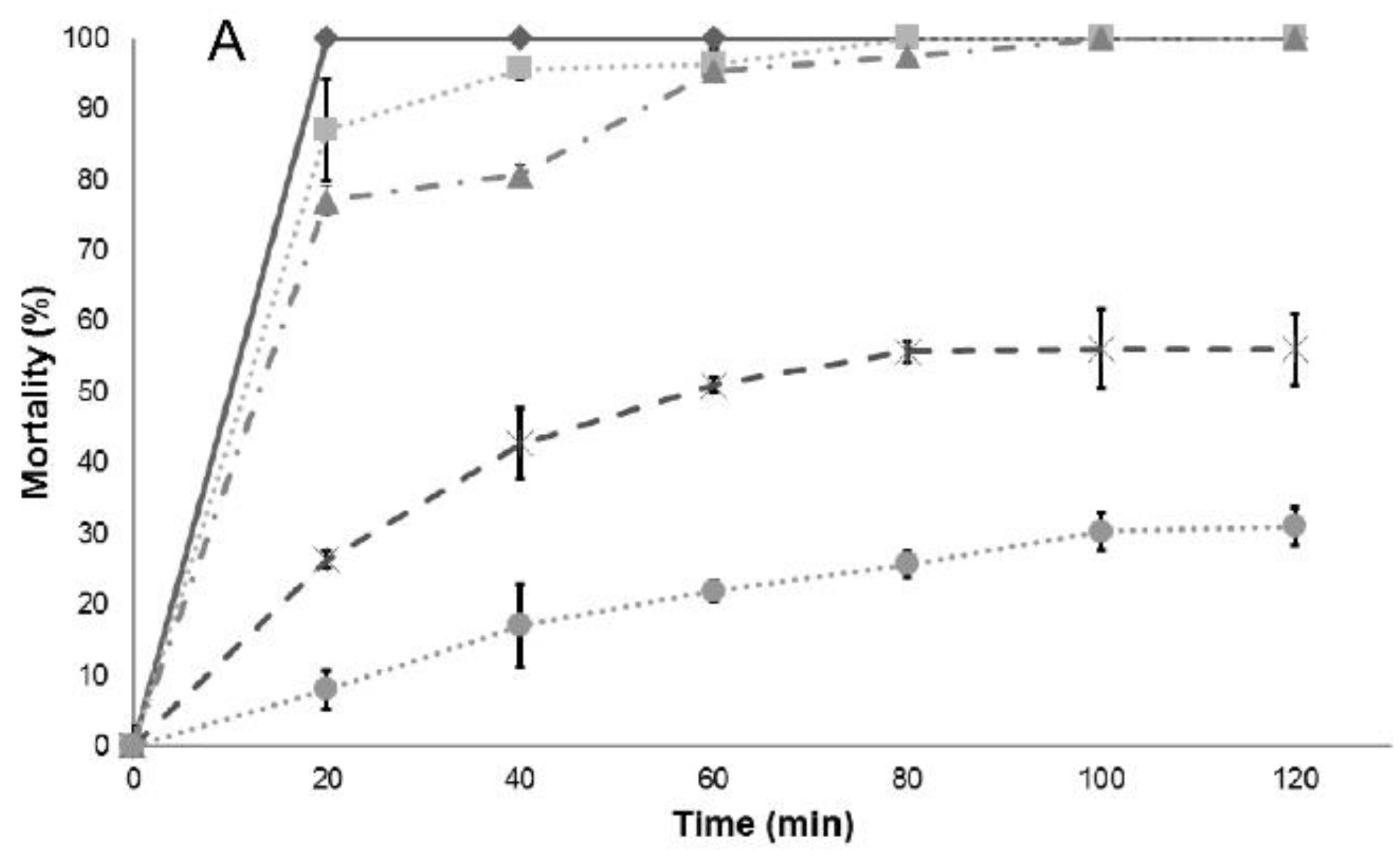
Table 3). However, it is not possible to assess differences among the essential oil or the main components, so the analysis of larval mortality was also included in this work. The essential oils and their main components included in this study also showed a high effectiveness with respect to mortality upon exposure of
Culex quinquefasciatus larvae to water contaminated with lethal doses (
Table 4): in most cases, deaths occurred after short-term exposures.
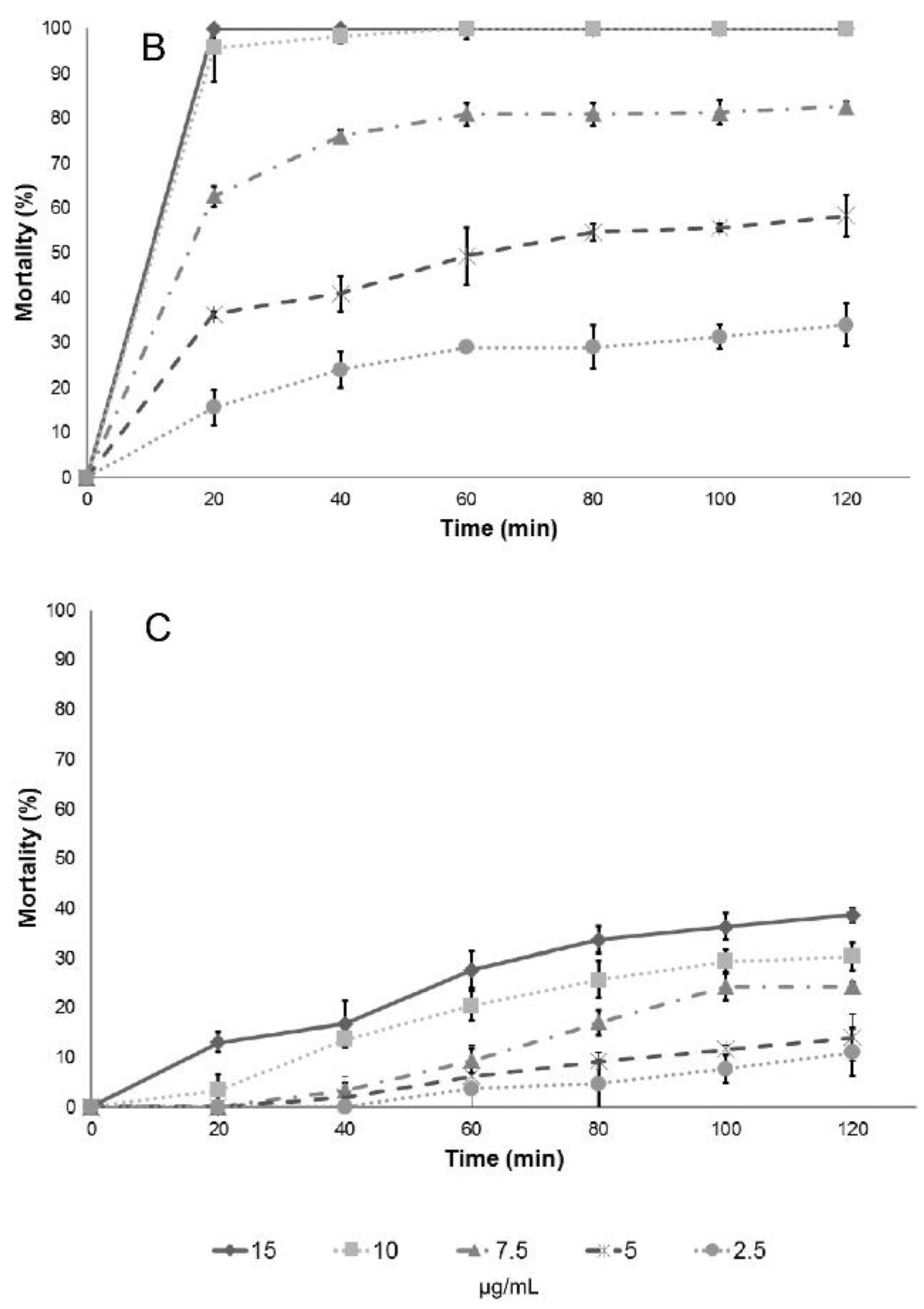
Results of larval mortality demonstrated that the activity of anise essential oil and trans-anethole are statistically similar to the activity of Temephos, while L. nobilis L. essential oil and its main component (eucalyptol) were less effective. Cinnamaldehyde, the major component of cinnamon essential oil, showed higher LC50 compared to the essential oil, suggesting that there are antagonistic effects of some components present in cinnamon essential oil. On the contrary, cuminaldehyde the major constituent of cumin essential oil, is more active than the essential oil.
Eugenol and eucalyptol, by themselves had better activity than the essential oil where they are present. In some cases, such as with eucalyptol that is present in bay laurel essential oil, the larvicidal activity doubles when the molecule is pure. This suggests that the other constituents of the essential oil have antagonistic interactions with eucalyptol. The low larvicidal activity of eucalyptol (1,8-cineol) has been reported previously against larvae of
Aedes aegypti [
33],
Anopheles anthropophagus [
34] and
Culex pipiens [
35], however it has been shown to be effective as a repellent in foods and highly effective as an ovipositional repellent [
33], however the effect did not last for more than 30 min [
36].
Limonene has a larvicidal activity similar to the essential oil of Mexican lime against stage III larvae of
C. quinquefasciatus. Its activity doubled when the molecule was evaluated as a pure compound on pupae. This suggests that other components of the essential oil have an antagonist effect with limonene or that the compound is more effective against larvae in a more mature stage. These results demonstrate the importance of studying the components of the essential oils separately, as different studies could rule out some important molecules because the oil as a whole has seemingly unimportant biological activities. Eleni et al. [
37] reported the larvicidal activity of limonene rich essential oil from
Citrus auranitium subsp. bergamia against
Culex pipiens; the oil displays a LC
50 of 58.73 mg/L. On the other hand,
Mentha longifolia essential oil, containing 20% limonene, has been reported as larvicidal against
Culex pipiens with a LC
50 of 78.28 mg/L after 48 h of exposure [
38].
There are multiple plant species collectively known as Oregano, including plants from the Verbenaceae and the Lamiaceae families, that have in common the presence of thymol and carvacrol in different proportions, but with similar odor and flavor characteristics [
22] and, as such, are comparable in biological activities. In this study, the essential oil of Mexican oregano (
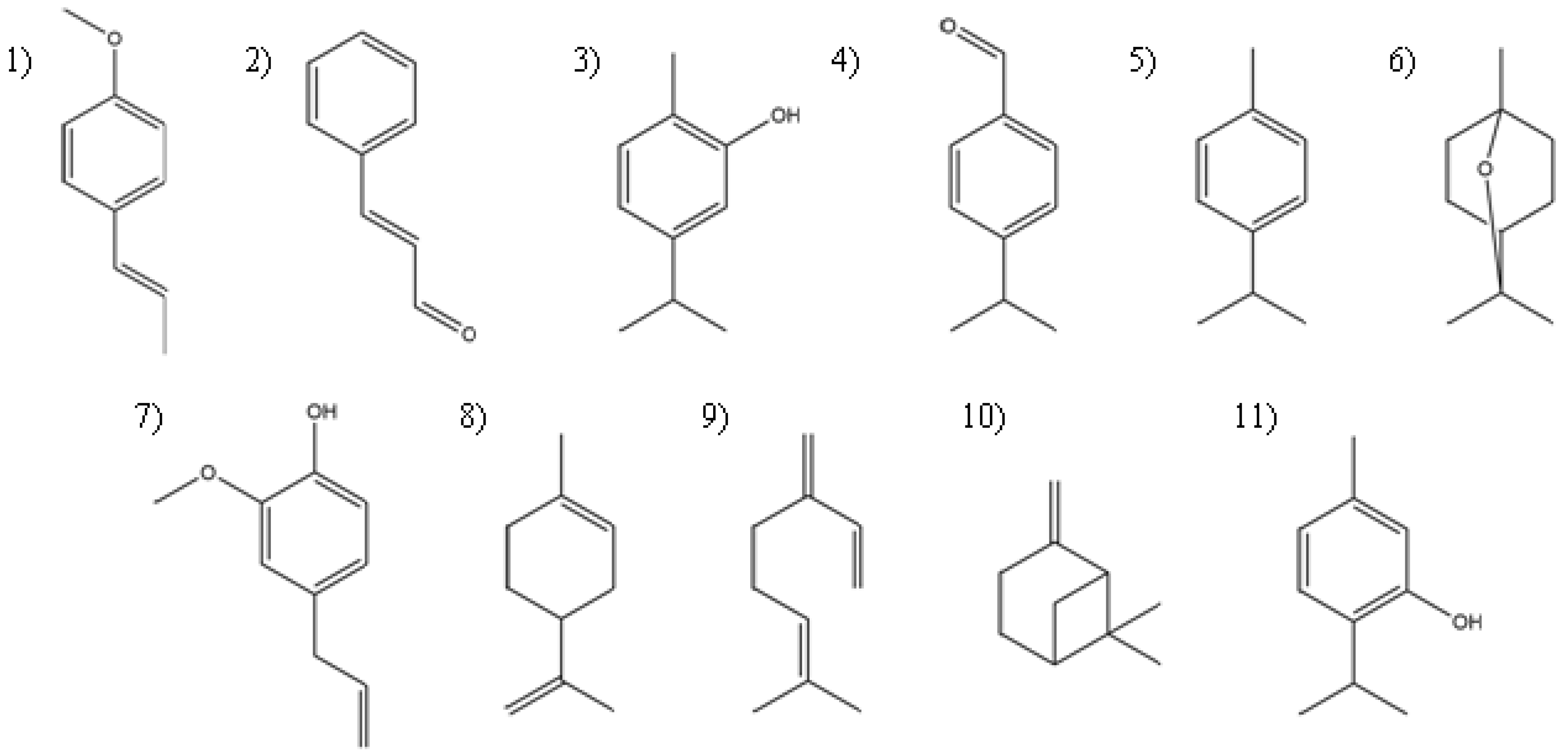
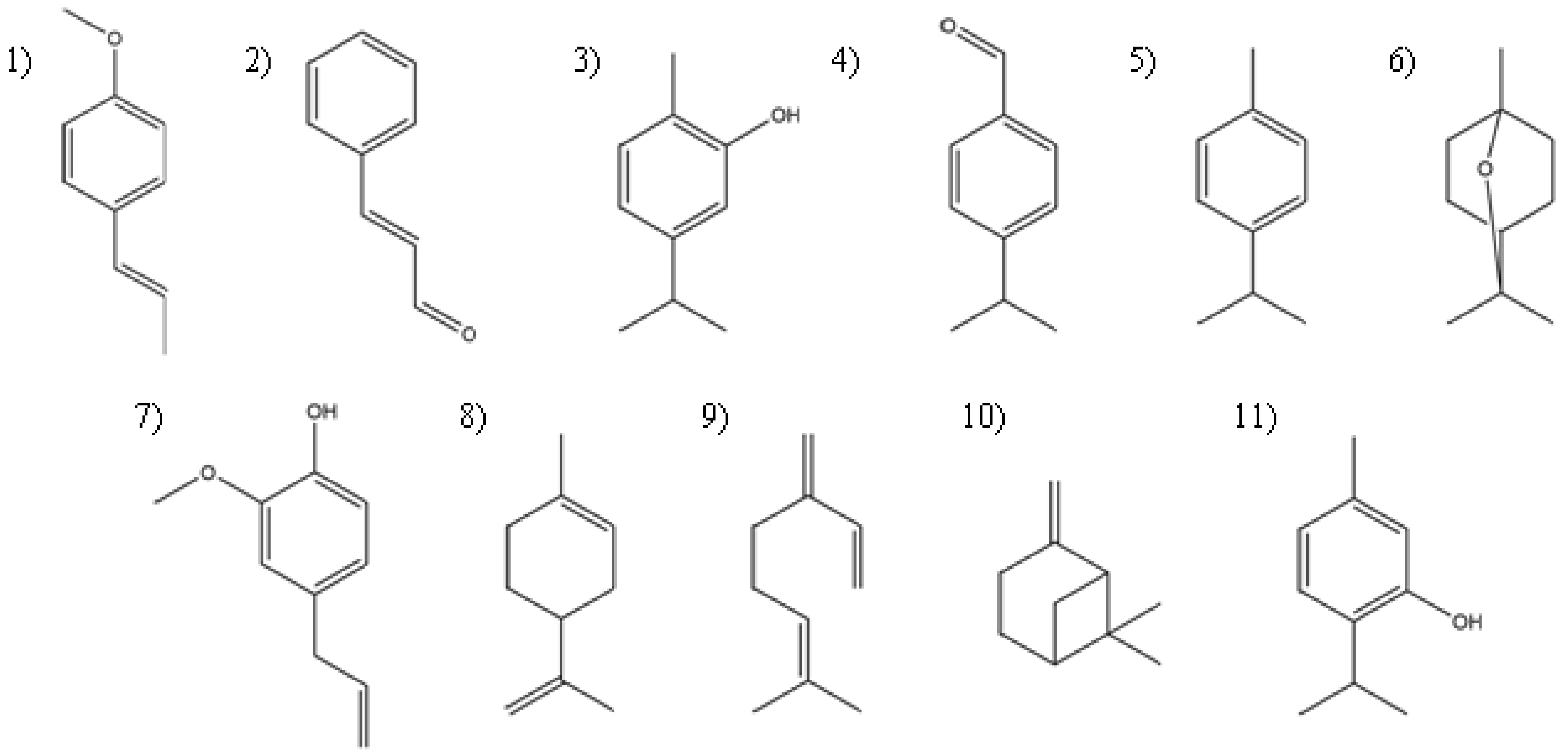
Lippia berlandieri Schauer) has carvacrol as its major constituent (57%), followed by thymol as the second major compound. There is no difference between the larvicidal activity of carvacrol and the essential oil of Mexican oregano, but the larvicidal activity of thymol is significantly lower. The only structural difference between carvacrol and thymol is the position of the hydroxyl group on the benzene ring with respect to the largest aliphatic chain (
Figure 1); this demonstrate the importance of the aliphatic chain and its proximity to other functional groups.
There are numerous reports on the insecticidal activity of the essential oils from
Origanum species; major components such as carvacrol, thymol, γ- terpinene and terpinen-4-ol are reported with fumigant and repellent activity rather than contact toxicity [
39]. Cetin and Yanikoglu determined the insecticidal activity of essential oils from two species of Origanum (
Origanum onites L. and
Origanum minutiflorum) on the third and fourth instar larvae of
Culex pipiens L.: the LC
50 values were 24.8 and 73.8 µg/mL, respectively [
40]. The authors attribute the larvicidal activity to carvacrol, as also suggested in this report. However, it is important to note that other studies have reported that oregano essential oil with high levels of thymol have an effective deterrent oviposition activity against
Culex quinquefasciatus females [
41].
Regarding essential oils derived from plants of the genus
Lippia, this study is the first to report the activity of the essential oil of
Lippia berlandieri Shauer. Vera et al. evaluated the activity of
Lippia alba and
Lippia origanoides against larvae of
Aedes aegypti [
42]. These two essential oils contained carvone and carvacrol as major components, and presented LC
50 values of approximately 50 µg/mL. In addition, the essential oils of
Lippia gracilis and
Lippia sidoides have been evaluated against
Aedes aegypti [
43,
44]. Gleiser and Zygadlo evaluated the activity of the essential oils of
Lippia turbinata and
Lippia polystachya against
Culex quinquefasciatus larvae [
45]. Both oils presented α-thujone as the major component and showed no relevant larvicidal activity.
The larvicidal efficiency of anise essential oil as well
trans-anethole, has been previously reported [
46]. Waliwitiya et al., Pavela and Sousa et al. evaluated the activity of
trans-anethole against larvae of
Aedes aegypti,
Anopheles atroparvus and
Culex quinquefasciatus respectively, finding relevant LC
50 values [
47,
48,
49]. This study confirms the activity of
Pimpinella anisum L. essential oil, but it shows that
trans-anethole does not eliminate the larvae as quickly as it does the essential oil, even though both have similar LC
50 values. This suggests that there are interactions with other components that are present in the essential oil at lower or even trace concentrations. Therefore, it is always important to analyze the activity of the complete essential oil and their major components, since the presence of compounds different than the main components, can present synergistic or antagonistic effects.
The most relevant synergistic effects are observed against larvae with the combination of trans-anetol and limonene and the combination of trans-anethole and β-myrcene against pupae. These effects may be responsible for the rapid larvicidal activity of the essential oil of P. anisum. Synergistic effects are also observed when mixing carvacrol and thymol, the main constituents of Lippia berlandieri essential oil. However, none of these compounds has additive effects on the activity when they are mixed with p-cymene. The only antagonistic effect was observed with the limonene–pinene combination, which may be related to the difference in activity between C.aurantifolia essential oil and limonene against pupae.
The mechanism of action with which the essential oils exert their larvicidal activity is not completely described. Pratti et al. suggest that instant death is due to severe damage of Malpighian tubules, since they are responsible for the excretion, not only of electrolytes and metabolites, but also of the high volume of water naturally present in insect larvae environment [
50]. There are reports suggesting that spice essential oils have neurotoxic poisoning-type effect on insects, similar to the ones produced by organophosphate and carbamate insecticides, by inhibition of the of the acetylcholinesterase enzyme (AChE) [
51]. In a comparative study of the vapor action of essential oils from plants of the Lamiaceae family on
R. dominica adults, it was observed that the essential oils inhibited about 65% of AChE activity, while limonene lowered only 2% of the enzymatic activity [
52]. Furthermore, these authors found that the essential oils significantly increased the levels of cyclic adenosine monophosphate (AMP) (even at very low concentrations), which suggests a possible action on octopamine. Similar results were obtained by Enan in flies and cockroaches exposed to eugenol and α-terpineol [
53]. In silico studies have demonstrated that terpenes can interact with AChE of
A. aegypti, by joining a hydrophobic site of the enzyme, with an interaction with glycine 412, 409, abd 412 and isoleucine 413 amino acids [
54]. Another proposed mechanism of action has been presented by Priestley et al., who suggest that thymol acts on GABA receptors of
Drosophila melanogaster [
55].



 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}