
RNA Interference for Mosquito and Mosquito-Borne Disease Control



Abstract
:1. Introduction
2. RNAi Triggers with Potential Mosquito Control Applications
2.1. RNAi to Induce a Lethal Phenotype
2.2. Olfaction
2.3. Blood Feeding & Digestion
2.4. Reproduction
2.5. Embryogenesis
2.6. Larval & Pupal Development
2.7. Detoxification and Insecticide Metabolism/Resistance
2.8. RNAi-Induced Pathogen Resistance
3. Delivery Systems for RNAi Triggers In Vivo
3.1. Naked RNA and Modified Nucleic Acids
3.2. Insect Plasmid Expression Systems
3.3. Nanoparticle Delivery Systems
3.4. Viral Expression Systems
3.5. Bacterial Expression Systems
3.6. The Pichia pastoris Expression System
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Livadas, G.A.; Georgopoulos, G. Development of resistance to DDT by Anopheles sacharovi in Greece. Bull. World Health Organ. 1953, 8, 497–511. [Google Scholar] [PubMed]
- Hammerstrom, R.J. Insect resistance to insecticides. Public Health Rep. 1958, 73, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Gjullin, C.M.; Peters, R.F. Recent studies of mosquito resistance to insecticides in California. Mosquito News 1952, 12, 1–7. [Google Scholar]
- Brown, A.W. Insecticide resistance in mosquitoes: A pragmatic review. J. Am. Mosq. Control Assoc. 1986, 2, 123–140. [Google Scholar] [PubMed]
- World Health Organization. World Malaria Report; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. Vector Resistance to Pesticides. Fifteenth Report of the WHO Expert Committee on Vector Biology and Control; World Health Organization Technical Report Series; World Health Organization: Geneva, Switzerland, 1992; Volume 818, pp. 1–68. [Google Scholar]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Zaim, M.; Guillet, P. Alternative insecticides: An urgent need. Trends Parasitol. 2002, 18, 161–163. [Google Scholar] [CrossRef]
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gort, T.; Boyle, D.L.; Clem, R.J. Effects of manipulating apoptosis on Sindbis virus infection of Aedes aegypti mosquitoes. J. Virol. 2012, 86, 6546–6554. [Google Scholar] [CrossRef] [PubMed]
- Coy, M.R.; Sanscrainte, N.D.; Chalaire, K.C.; Inberg, A.; Maayan, I.; Glick, E.; Paldi, N.; Becnel, J.J. Gene silencing in adult Aedes aegypti mosquitoes through oral delivery of double-stranded RNA. J. Appl. Entomol. 2012, 136, 741–748. [Google Scholar] [CrossRef]
- Singh, A.D.; Wong, S.; Ryan, C.P.; Whyard, S. Oral delivery of double-stranded RNA in larvae of the yellow fever mosquito, Aedes aegypti: Implications for pest mosquito control. J. Insect Sci. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Whyard, S.; Erdelyan, C.N.; Partridge, A.L.; Singh, A.D.; Beebe, N.W.; Capina, R. Silencing the buzz: A new approach to population suppression of mosquitoes by feeding larvae double-stranded RNAs. Parasites Vectors 2015. [Google Scholar] [CrossRef] [PubMed]
- Thailayil, J.; Magnusson, K.; Godfray, H.C.; Crisanti, A.; Catteruccia, F. Spermless males elicit large-scale female responses to mating in the malaria mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2011, 108, 13677–13681. [Google Scholar] [CrossRef] [PubMed]
- Erdelyan, C.N.; Mahood, T.H.; Bader, T.S.; Whyard, S. Functional validation of the carbon dioxide receptor genes in Aedes aegypti mosquitoes using RNA interference. Insect Mol. Biol. 2012, 21, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Guidolin, A.; Syed, Z.; Cornel, A.J.; Leal, W.S. Knockdown of a mosquito odorant-binding protein involved in the sensitive detection of oviposition attractants. J. Chem. Ecol. 2010, 36, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Chagas, A.C.; Ramirez, J.L.; Jasinskiene, N.; James, A.A.; Ribeiro, J.M.; Marinotti, O.; Calvo, E. Collagen-binding protein, Aegyptin, regulates probing time and blood feeding success in the dengue vector mosquito, Aedes aegypti. Proc. Natl. Acad. Sci. USA 2014, 111, 6946–6951. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Hansen, I.A.; Shiao, S.H.; Raikhel, A.S. Identification of two cationic amino acid transporters required for nutritional signaling during mosquito reproduction. J. Exp. Biol. 2006, 209 Pt 16, 3071–3078. [Google Scholar] [CrossRef] [PubMed]
- Tsao, I.Y.; Lin, U.S.; Christensen, B.M.; Chen, C.C. Armigeres subalbatus prophenoloxidase III: Cloning, characterization and potential role in morphogenesis. Insect Biochem. Mol. Biol. 2009, 39, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, D.D. Creating a ribonuclease-free environment. Methods Enzymol. 1987, 152, 20–24. [Google Scholar] [PubMed]
- Dubelman, S.; Fischer, J.; Zapata, F.; Huizinga, K.; Jiang, C.; Uffman, J.; Levine, S.; Carson, D. Environmental fate of double-stranded RNA in agricultural soils. PLoS ONE 2014, 9, e93155. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.R.; Zapata, F.; Dubelman, S.; Mueller, G.M.; Uffman, J.P.; Jiang, C.; Jensen, P.D.; Levine, S.L. Aquatic fate of a double-stranded RNA in a sediment-water system following an over-water application. Environ. Toxicol. Chem. 2016. [Google Scholar] [CrossRef] [PubMed]
- San Miguel, K.; Scott, J.G. The next generation of insecticides: DsRNA is stable as a foliar-applied insecticide. Pest Manag. Sci. 2016, 72, 801–809. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention, and Control; New Edition Special Programme for Research and Training in Tropical Diseases; World Health Organization: Geneva, Switzerland, 2009; pp. 59–86. [Google Scholar]
- World Health Organization. Indoor Residual Spraying: An Operational Manual for IRS for Malaria Transmission, Control and Elimination, 2nd ed.; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. Malaria Vector Control and Personal Protection: Report of a WHO Study Group; WHO Technical Report Series; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Van Ekert, E.; Powell, C.A.; Shatters, R.G., Jr.; Borovsky, D. Control of larval and egg development in Aedes aegypti with RNA interference against juvenile hormone acid methyl transferase. J. Insect Physiol. 2014, 70, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.C.; Beier, J.C.; Traore, S.F.; Toure, M.B.; Traore, M.M.; Bah, S.; Doumbia, S.; Schlein, Y. Field experiments of Anopheles gambiae attraction to local fruits/seedpods and flowering plants in Mali to optimize strategies for malaria vector control in Africa using attractive toxic sugar bait methods. Malar. J. 2010. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.A. Susceptibility of adult mosquitoes to insecticides in aqueous sucrose baits. J. Vector Ecol. 2011, 36, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Muller, G.C.; Revay, E.E.; Allan, S.A.; Arheart, K.L.; Beier, J.C.; Smith, M.L.; Scott, J.M.; Kravchenko, V.D.; Hausmann, A.; et al. Evaluation of attractive toxic sugar bait (ATSB)-Barrier for control of vector and nuisance mosquitoes and its effect on non-target organisms in sub-tropical environments in Florida. Acta Trop. 2014, 131, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Marshall, J.M.; Qualls, W.A.; Schlein, Y.; McManus, J.W.; Arheart, K.L.; Hlaing, W.M.; Traore, S.F.; Doumbia, S.; Muller, G.C.; et al. Modelling optimum use of attractive toxic sugar bait stations for effective malaria vector control in Africa. Malar. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Qualls, W.A.; Muller, G.C.; Traore, S.F.; Traore, M.M.; Arheart, K.L.; Doumbia, S.; Schlein, Y.; Kravchenko, V.D.; Xue, R.D.; Beier, J.C. Indoor use of attractive toxic sugar bait (ATSB) to effectively control malaria vectors in Mali, West Africa. Malar. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Stewart, Z.P.; Oxborough, R.M.; Tungu, P.K.; Kirby, M.J.; Rowland, M.W.; Irish, S.R. Indoor application of attractive toxic sugar bait (ATSB) in combination with mosquito nets for control of pyrethroid-resistant mosquitoes. PLoS ONE 2013, 8, e84168. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.C.; Beier, J.C.; Traore, S.F.; Toure, M.B.; Traore, M.M.; Bah, S.; Doumbia, S.; Schlein, Y. Successful field trial of attractive toxic sugar bait (ATSB) plant-spraying methods against malaria vectors in the Anopheles gambiae complex in Mali, West Africa. Malar. J. 2010. [Google Scholar] [CrossRef] [PubMed]
- Beier, J.C.; Muller, G.C.; Gu, W.; Arheart, K.L.; Schlein, Y. Attractive toxic sugar bait (ATSB) methods decimate populations of Anopheles malaria vectors in arid environments regardless of the local availability of favoured sugar-source blossoms. Malar. J. 2012. [Google Scholar] [CrossRef] [PubMed]
- Revay, E.E.; Muller, G.C.; Qualls, W.A.; Kline, D.L.; Naranjo, D.P.; Arheart, K.L.; Kravchenko, V.D.; Yefremova, Z.; Hausmann, A.; Beier, J.C.; et al. Control of Aedes albopictus with attractive toxic sugar baits (ATSB) and potential impact on non-target organisms in St. Augustine, Florida. Parasitol. Res. 2014, 113, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Schlein, Y.; Muller, G.C. Experimental control of Phlebotomus papatasi by spraying attractive toxic sugar bait (ATSB) on vegetation. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.M.; Hu, Y.; Yu, B.T.; Mo, X.C.; Mo, J.C. Laboratory evaluation of differential attraction of Culex pipiens pallens to fruit-based sugar baits. Acta Trop. 2016, 163, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Junnila, A.; Revay, E.E.; Muller, G.C.; Kravchenko, V.; Qualls, W.A.; Xue, R.D.; Allen, S.A.; Beier, J.C.; Schlein, Y. Efficacy of attractive toxic sugar baits (ATSB) against Aedes albopictus with garlic oil encapsulated in beta-cyclodextrin as the active ingredient. Acta Trop. 2015, 152, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.C.; Junnila, A.; Qualls, W.; Revay, E.E.; Kline, D.L.; Allan, S.; Schlein, Y.; Xue, R.D. Control of Culex quinquefasciatus in a storm drain system in Florida using attractive toxic sugar baits. Med. Vet. Entomol. 2010, 24, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Muller, G.C.; Junnila, A.; Schlein, Y. Effective control of adult Culex pipiens by spraying an attractive toxic sugar bait solution in the vegetation near larval habitats. J. Med. Entomol. 2010, 47, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Pietri, J.E.; Cheung, K.W.; Luckhart, S. Knockdown of mitogen-activated protein kinase (MAPK) signalling in the midgut of Anopheles stephensi mosquitoes using antisense morpholinos. Insect Mol. Biol. 2014, 23, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Baak-Baak, C.M.; Rodriguez-Ramirez, A.D.; Garcia-Rejon, J.E.; Rios-Delgado, S.; Torres-Estrada, J.L. Development and laboratory evaluation of chemically-based baited ovitrap for the monitoring of Aedes aegypti. J. Vector Ecol. 2013, 38, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Lampman, R.L.; Novak, R.J. Oviposition preferences of Culex pipiens and Culex restuans for infusion-baited traps. J. Am. Mosq. Control Assoc. 1996, 12, 23–32. [Google Scholar] [PubMed]
- Harris, C.; Kihonda, J.; Lwetoijera, D.; Dongus, S.; Devine, G.; Majambere, S. A simple and efficient tool for trapping gravid Anopheles at breeding sites. Parasites Vectors 2011. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Debnath, N.; Cui, Y.; Unrine, J.; Palli, S.R. Chitosan, Carbon Quantum Dot, and Silica Nanoparticle Mediated dsRNA Delivery for Gene Silencing in Aedes aegypti: A Comparative Analysis. ACS Appl. Mater. Interfaces 2015, 7, 19530–19535. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.; Flannery, E.M.; Tomchaney, M.; Severson, D.W.; Duman-Scheel, M. Disruption of Aedes aegypti olfactory system development through chitosan/siRNA nanoparticle targeting of semaphorin-1A. PLoS Negl. Trop. Dis. 2013, 7, e2215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hua, G.; Adang, M.J. Chitosan/DsiRNA nanoparticle targeting identifies AgCad1 cadherin in Anopheles gambiae larvae as an in vivo receptor of Cry11Ba toxin of Bacillus thuringiensis subsjegathesan. Insect Biochem. Mol. Biol. 2015, 60, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Paquette, C.C.; Phanse, Y.; Perry, J.L.; Sanchez-Vargas, I.; Airs, P.M.; Dunphy, B.M.; Xu, J.; Carlson, J.O.; Luft, J.C.; DeSimone, J.M.; et al. Biodistribution and trafficking of hydrogel nanoparticles in adult mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003745. [Google Scholar] [CrossRef] [PubMed]
- Phanse, Y.; Dunphy, B.M.; Perry, J.L.; Airs, P.M.; Paquette, C.C.; Carlson, J.O.; Xu, J.; Luft, J.C.; DeSimone, J.M.; Beaty, B.J.; et al. Biodistribution and Toxicity Studies of PRINT Hydrogel Nanoparticles in Mosquito Larvae and Cells. PLoS Negl. Trop. Dis. 2015, 9, e0003735. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Clem, R.J. Defining the core apoptosis pathway in the mosquito disease vector Aedes aegypti: The roles of iap1, ark, dronc, and effector caspases. Apoptosis 2011, 16, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, J.W.; Zhao, L.; Becnel, J.J.; Strickman, D.A.; Clark, G.G.; Linthicum, K.J. Topically applied AaeIAP1 double-stranded RNA kills female adults of Aedes aegypti. J. Med. Entomol. 2008, 45, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Puglise, J.M.; Estep, A.S.; Becnel, J.J. Expression Profiles and RNAi Silencing of Inhibitor of Apoptosis Transcripts in Aedes, Anopheles, and Culex Mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2016, 53, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, C.B.; Caicedo, P.A.; Jaramillo, G.; Bedoya, R.U.; Baron, O.; Serrato, I.M.; Cooper, D.M.; Lowenberger, C. Differential expression of apoptosis related genes in selected strains of Aedes aegypti with different susceptibilities to dengue virus. PLoS ONE 2013, 8, e61187. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.J.; Brown, M.R.; Strand, M.R. Ovary ecdysteroidogenic hormone requires a receptor tyrosine kinase to activate egg formation in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2015, 112, 5057–5062. [Google Scholar] [CrossRef] [PubMed]
- Clemons, A.; Haugen, M.; Le, C.; Mori, A.; Tomchaney, M.; Severson, D.W.; Duman-Scheel, M. siRNA-mediated gene targeting in Aedes aegypti embryos reveals that frazzled regulates vector mosquito CNS development. PLoS ONE 2011, 6, e16730. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.F.; Guo, Q.; Sun, Y.; Zhou, D.; Hu, M.X.; Hu, H.X.; Liu, B.Q.; Tian, M.M.; Liu, X.M.; Li, X.X.; et al. Identification of protease m1 zinc metalloprotease conferring resistance to deltamethrin by characterization of an AFLP marker in Culex pipiens pallens. Parasites Vectors 2016. [Google Scholar] [CrossRef] [PubMed]
- Clayton, A.M.; Cirimotich, C.M.; Dong, Y.; Dimopoulos, G. Caudal is a negative regulator of the Anopheles IMD pathway that controls resistance to Plasmodium falciparum infection. Dev. Comp. Immunol. 2013, 39, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Papaefthimiou, C.; Theophilidis, G. The cardiotoxic action of the pyrethroid insecticide deltamethrin, the azole fungicide prochloraz, and their synergy on the semi-isolated heart of the bee Apis mellifera macedonica. Pest. Biochem. Physiol. 2001, 69, 77–91. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Antwi, F.B.; Reddy, G.V. Toxicological effects of pyrethroids on non-target aquatic insects. Environ. Toxicol. Pharmacol. 2015, 40, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G. Current issues in organophosphate toxicology. Clin. Chim. Acta 2006, 366, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, M.; Zafra, S.; Bagchi, D. DNA Damage, Gene Expression, and Carcinogenesis by Organophosphates and Carbamates. In Toxicology of Organophosphate & Carbamate Compounds; Gupta, R.C., Ed.; Academic Press: Burlington, VT, USA, 2006; pp. 533–548. [Google Scholar]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.J.; Vucic, D.; Miller, L.K. The Drosophila inhibitor of apoptosis D-IAP1 suppresses cell death induced by the caspase drICE. FEBS Lett. 1998, 440, 243–248. [Google Scholar] [CrossRef]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Luthi, A.U.; Martin, S.J. The CASBAH: A searchable database of caspase substrates. Cell Death Differ. 2007, 14, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, F.; Ghani, Y.; Kafatos, F.C.; Christophides, G.K. Comprehensive genetic dissection of the hemocyte immune response in the malaria mosquito Anopheles gambiae. PLoS Pathog. 2013, 9, e1003145. [Google Scholar] [CrossRef] [PubMed]
- Boutros, M.; Kiger, A.A.; Armknecht, S.; Kerr, K.; Hild, M.; Koch, B.; Haas, S.A.; Paro, R.; Perrimon, N.; Array, C.H.F. Genome-wide RNAi analysis of growth and viability in Drosophila cells. Science 2004, 303, 832–835. [Google Scholar] [CrossRef] [PubMed]
- Zdobnov, E.M.; von Mering, C.; Letunic, I.; Torrents, D.; Suyama, M.; Copley, R.R.; Christophides, G.K.; Thomasova, D.; Holt, R.A.; Subramanian, G.M.; et al. Comparative genome and proteome analysis of Anopheles gambiae and Drosophila melanogaster. Science 2002, 298, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Huang, J.H.; Liu, Y.; Belles, X.; Lee, H.J. Oral delivery of dsRNA lipoplexes to German cockroach protects dsRNA from degradation and induces RNAi response. Pest Manag. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Whitten, M.M.; PFacey, D.; del Sol, R.; Fernandez-Martinez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. Biol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Cancino-Rodezno, A.; Lozano, L.; Oppert, C.; Castro, J.I.; Lanz-Mendoza, H.; Encarnacion, S.; Evans, A.E.; Gill, S.S.; Soberon, M.; Jurat-Fuentes, J.L.; et al. Comparative proteomic analysis of Aedes aegypti larval midgut after intoxication with Cry11Aa toxin from Bacillus thuringiensis. PLoS ONE 2012, 7, e37034. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Perzov, N.; Cohen, A.; Hagai, K.; Padler, V.; Nelson, H. The cellular biology of proton-motive force generation by V-ATPases. J. Exp. Biol. 2000, 203, 89–95. [Google Scholar] [PubMed]
- Allan, A.K.; Du, J.; Davies, S.A.; Dow, J.A. Genome-wide survey of V-ATPase genes in Drosophila reveals a conserved renal phenotype for lethal alleles. Physiol. Genom. 2005, 22, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Wuriyanghan, H.; Rosa, C.; Falk, B.W. Oral delivery of double-stranded RNAs and siRNAs induces RNAi effects in the potato/tomato psyllid, Bactericerca cockerelli. PLoS ONE 2011, 6, e27736. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zhang, P.; Liu, C.; Zeng, F. Co-silence of the coatomer beta and V-ATPase A genes by siRNA feeding reduces larval survival rate and weight gain of cotton bollworm, Helicoverpa armigera. Pestic. Biochem. Physiol. 2015, 118, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Shields, A.R.; Jupatanakul, N.; Dimopoulos, G. Suppressing dengue-2 infection by chemical inhibition of Aedes aegypti host factors. PLoS Negl. Trop. Dis. 2014, 8, e3084. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Liu, M.; Deng, Y.; Peng, H.; Chen, X. Development of an efficient recombinant mosquito densovirus-mediated RNA interference system and its preliminary application in mosquito control. PLoS ONE 2011, 6, e21329. [Google Scholar] [CrossRef] [PubMed]
- Isoe, J.; Collins, J.; Badgandi, H.; Day, W.A.; Miesfeld, R.L. Defects in coatomer protein I (COPI) transport cause blood feeding-induced mortality in Yellow Fever mosquitoes. Proc. Natl. Acad. Sci. USA 2011, 108, E211–E217. [Google Scholar] [CrossRef] [PubMed]
- Isoe, J.; Stover, W.; Miesfeld, R.B.; Miesfeld, R.L. COPI-mediated blood meal digestion in vector mosquitoes is independent of midgut ARF-GEF and ARF-GAP regulatory activities. Insect Biochem. Mol. Biol. 2013, 43, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Isoe, J.; Day, W.A.; Miesfeld, R.L. Alpha-COPI coatomer protein is required for rough endoplasmic reticulum whorl formation in mosquito midgut epithelial cells. PLoS ONE 2011, 6, e18150. [Google Scholar] [CrossRef] [PubMed]
- Tsao, I.Y.; Christensen, B.M.; Chen, C.C. Armigeres subalbatus (Diptera: Culicidae) prophenoloxidase III is required for mosquito cuticle formation: Ultrastructural study on dsRNA-knockdown mosquitoes. J. Med. Entomol. 2010, 47, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- DeGennaro, M.; McBride, C.S.; Seeholzer, L.; Nakagawa, T.; Dennis, E.J.; Goldman, C.; Jasinskiene, N.; James, A.A.; Vosshall, L.B. Orco mutant mosquitoes lose strong preference for humans and are not repelled by volatile DEET. Nature 2013, 498, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Choo, Y.M.; de la Rosa, A.; Leal, W.S. Mosquito odorant receptor for DEET and methyl jasmonate. Proc. Natl. Acad. Sci. USA 2014, 111, 16592–16597. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, J.; Leal, W.S. Characterization of olfactory genes in the antennae of the Southern house mosquito, Culex quinquefasciatus. J. Insect Physiol. 2011, 57, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.X.; Zwiebel, L.J.; Smith, D. Identification of a distinct family of genes encoding atypical odorant-binding proteins in the malaria vector mosquito, Anopheles gambiae. Insect Mol. Biol. 2003, 12, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; He, X.L.; Pickett, J.A.; Field, L.M. Identification of odorant-binding proteins of the yellow fever mosquito Aedes aegypti: Genome annotation and comparative analyses. Insect Mol. Biol. 2008, 17, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Rinker, D.C.; Pitts, R.J.; Zhou, X.; Suh, E.; Rokas, A.; Zwiebel, L.J. Blood meal-induced changes to antennal transcriptome profiles reveal shifts in odor sensitivities in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2013, 110, 8260–8265. [Google Scholar] [CrossRef] [PubMed]
- Rinker, D.C.; Zhou, X.; Pitts, R.J.; Consortium, A.G.C.; Rokas, A.; Zwiebel, L.J. Antennal transcriptome profiles of anopheline mosquitoes reveal human host olfactory specialization in Anopheles gambiae. BMC Genom. 2013. [Google Scholar] [CrossRef] [PubMed]
- Won Jung, J.; Baeck, S.-J.; Perumalsamy, H.; Hansson, B.S.; Ahn, Y.-J.; Kwon, H.W. A novel olfactory pathway is essential for fast and efficient blood-feeding in mosquitoes. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, T.; Xie, L.; Wang, X.; Deng, Y.; Chen, C.H.; James, A.A.; Chen, X.G. Functional analysis of Orco and odorant receptors in odor recognition in Aedes albopictus. Parasites Vectors 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Xu, P.; Barbosa, R.M.; Choo, Y.M.; Leal, W.S. RNAi-based demonstration of direct link between specific odorant receptors and mosquito oviposition behavior. Insect Biochem. Mol. Biol. 2013, 43, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dimopoulos, G. Molecular analysis of photic inhibition of blood-feeding in Anopheles gambiae. BMC Physiol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Marinotti, O.; Nguyen, Q.K.; Calvo, E.; James, A.A.; Ribeiro, J.M. Microarray analysis of genes showing variable expression following a blood meal in Anopheles gambiae. Insect Mol. Biol. 2005, 14, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Marinotti, O.; Calvo, E.; Nguyen, Q.K.; Dissanayake, S.; Ribeiro, J.M.; James, A.A. Genome-wide analysis of gene expression in adult Anopheles gambiae. Insect Mol. Biol. 2006, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Isoe, J.; Rascon, A.A., Jr.; Kunz, S.; Miesfeld, R.L. Molecular genetic analysis of midgut serine proteases in Aedes aegypti mosquitoes. Insect Biochem. Mol. Biol. 2009, 39, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Isoe, J.; Scaraffia, P.Y. Urea synthesis and excretion in Aedes aegypti mosquitoes are regulated by a unique cross-talk mechanism. PLoS ONE 2013, 8, e65393. [Google Scholar] [CrossRef] [PubMed]
- Overend, G.; Cabrero, P.; Halberg, K.A.; Ranford-Cartwright, L.C.; Woods, D.J.; Davies, S.A.; Dow, J.A. A comprehensive transcriptomic view of renal function in the malaria vector, Anopheles gambiae. Insect Biochem. Mol. Biol. 2015, 67, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Drake, L.L.; Boudko, D.Y.; Marinotti, O.; Carpenter, V.K.; Dawe, A.L.; Hansen, I.A. The Aquaporin gene family of the yellow fever mosquito, Aedes aegypti. PLoS ONE 2010, 5, e15578. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Tsujimoto, H.; Cha, S.J.; Agre, P.; Rasgon, J.L. Aquaporin water channel AgAQP1 in the malaria vector mosquito Anopheles gambiae during blood feeding and humidity adaptation. Proc. Natl. Acad. Sci. USA 2011, 108, 6062–6066. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.A.; Attardo, G.M.; Roy, S.G.; Raikhel, A.S. Target of rapamycin-dependent activation of S6 kinase is a central step in the transduction of nutritional signals during egg development in a mosquito. J. Biol. Chem. 2005, 280, 20565–20572. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.A.; Attardo, G.M.; Park, J.H.; Peng, Q.; Raikhel, A.S. Target of rapamycin-mediated amino acid signaling in mosquito anautogeny. Proc. Natl. Acad. Sci. USA 2004, 101, 10626–10631. [Google Scholar] [CrossRef] [PubMed]
- Raphemot, R.; Estevez-Lao, T.Y.; Rouhier, M.F.; Piermarini, P.M.; Denton, J.S.; Hillyer, J.F. Molecular and functional characterization of Anopheles gambiae inward rectifier potassium (Kir1) channels: A novel role in egg production. Insect Biochem. Mol. Biol. 2014, 51, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Telang, A.; Rechel, J.A.; Brandt, J.R.; Donnell, D.M. Analysis of ovary-specific genes in relation to egg maturation and female nutritional condition in the mosquitoes Georgecraigius atropalpus and Aedes aegypti (Diptera: Culicidae). J. Insect Physiol. 2013, 59, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D.M.; Ahuja, L.G.; Nunes, R.D.; Cudischevitch, C.O.; Daumas-Filho, C.R.; Medeiros-Castro, P.; Ventura-Martins, G.; Jablonka, W.; Gazos-Lopes, F.; Senna, R.; et al. Molecular analysis of Aedes aegypti classical protein tyrosine phosphatases uncovers an ortholog of mammalian PTP-1B implicated in the control of egg production in mosquitoes. PLoS ONE 2014, 9, e104878. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.A.; Boudko, D.Y.; Shiao, S.H.; Voronov, D.A.; Meleshkevitch, E.A.; Drake, L.L.; Aguirre, S.E.; Fox, J.M.; Attardo, G.M.; Raikhel, A.S. AaCAT1 of the yellow fever mosquito, Aedes aegypti: A novel histidine-specific amino acid transporter from the SLC7 family. J. Biol. Chem. 2011, 286, 10803–10813. [Google Scholar] [CrossRef] [PubMed]
- Sim, C.; Denlinger, D.L. A shut-down in expression of an insulin-like peptide, ILP-1, halts ovarian maturation during the overwintering diapause of the mosquito Culex pipiens. Insect Mol. Biol. 2009, 18, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.W.; Baldini, F.; Battaglia, F.; Panico, M.; Dell, A.; Morris, H.R.; Catteruccia, F. Transglutaminase-mediated semen coagulation controls sperm storage in the malaria mosquito. PLoS Biol. 2009, 7, e1000272. [Google Scholar] [CrossRef] [PubMed]
- Hoang, K.P.; Teo, T.M.; Ho, T.X.; Le, V.S. Mechanisms of sex determination and transmission ratio distortion in Aedes aegypti. Parasites Vectors 2016. [Google Scholar] [CrossRef] [PubMed]
- Akbari, O.S.; Antoshechkin, I.; Amrhein, H.; Williams, B.; Diloreto, R.; Sandler, J.; Hay, B.A. The developmental transcriptome of the mosquito Aedes aegypti, an invasive species and major arbovirus vector. G3 2013, 3, 1493–1509. [Google Scholar] [CrossRef] [PubMed]
- Koutsos, A.C.; Blass, C.; Meister, S.; Schmidt, S.; MacCallum, R.M.; Soares, M.B.; Collins, F.H.; Benes, V.; Zdobnov, E.; Kafatos, F.C.; et al. Life cycle transcriptome of the malaria mosquito Anopheles gambiae and comparison with the fruitfly Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2007, 104, 11304–11309. [Google Scholar] [CrossRef] [PubMed]
- Sarro, J.; Andrews, E.; Sun, L.; Behura, S.K.; Tan, J.C.; Zeng, E.; Severson, D.W.; Duman-Scheel, M. Requirement for commissureless2 function during dipteran insect nerve cord development. Dev. Dyn. 2013, 242, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Haugen, M.; Flannery, E.; Tomchaney, M.; Mori, A.; Behura, S.K.; Severson, D.W.; Duman-Scheel, M. Semaphorin-1A is required for Aedes aegypti embryonic nerve cord development. PLoS ONE 2011, 6, e21694. [Google Scholar] [CrossRef] [PubMed]
- Meleshkevitch, E.A.; Voronov, D.A.; Miller, M.M.; Penneda, M.; Fox, J.M.; Metzler, R.; Boudko, D.Y. A novel eukaryotic Na+ methionine selective symporter is essential for mosquito development. Insect Biochem. Mol. Biol. 2013, 43, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Fu, Q.; Hong, H.; Schwaegler, T.; Lan, Q. THAP and ATF-2 regulated sterol carrier protein-2 promoter activities in the larval midgut of the yellow fever mosquito, Aedes aegypti. PLoS ONE 2012, 7, e46948. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Maklokova, V.I.; Chandrashekhar, J.H.; Lan, Q. In vivo functional genomic studies of sterol carrier protein-2 gene in the yellow fever mosquito. PLoS ONE 2011, 6, e18030. [Google Scholar] [CrossRef] [PubMed]
- Bucher, G.; Scholten, J.; Klingler, M. Parental RNAi in Tribolium (Coleoptera). Curr. Biol. 2002, 12, R85–R86. [Google Scholar] [CrossRef]
- Cancino-Rodezno, A.; Alexander, C.; Villasenor, R.; Pacheco, S.; Porta, H.; Pauchet, Y.; Soberon, M.; Gill, S.S.; Bravo, A. The mitogen-activated protein kinase p38 is involved in insect defense against Cry toxins from Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2010, 40, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Bedoya-Perez, L.P.; Cancino-Rodezno, A.; Flores-Escobar, B.; Soberon, M.; Bravo, A. Role of UPR pathway in defense response of Aedes aegypti against Cry11Aa toxin from Bacillus thuringiensis. Int. J. Mol. Sci. 2013, 14, 8467–8478. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Martinez, G.; Meuti, M.; Denlinger, D.L. Rehydration driven RNAi: A novel approach for effectively delivering dsRNA to mosquito larvae. J. Med. Entomol. 2012, 49, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Pitts, R.J.; Bohbot, J.D.; Jones, P.L.; Wang, G.; Zwiebel, L.J. Distinct olfactory signaling mechanisms in the malaria vector mosquito Anopheles gambiae. PLoS Biol. 2010, 8, e1000467. [Google Scholar] [CrossRef] [PubMed]
- Saengwiman, S.; Aroonkesorn, A.; Dedvisitsakul, P.; Sakdee, S.; Leetachewa, S.; Angsuthanasombat, C.; Pootanakit, K. In vivo identification of Bacillus thuringiensis Cry4Ba toxin receptors by RNA interference knockdown of glycosylphosphatidylinositol-linked aminopeptidase N transcripts in Aedes aegypti larvae. Biochem. Biophys. Res. Commun. 2011, 407, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Figueira-Mansur, J.; Ferreira-Pereira, A.; Mansur, J.F.; Franco, T.A.; Alvarenga, E.S.; Sorgine, M.H.; Neves, B.C.; Melo, A.C.; Leal, W.S.; Masuda, H.; et al. Silencing of P-glycoprotein increases mortality in temephos-treated Aedes aegypti larvae. Insect Mol. Biol. 2013, 22, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.; Andrews, E.; Li, P.; Duman-Scheel, M. Chitosan/siRNA nanoparticle targeting demonstrates a requirement for single-minded during larval and pupal olfactory system development of the vector mosquito Aedes aegypti. BMC Dev. Biol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mysore, K.; Flannery, E.; Michel, K.; Severson, D.W.; Zhu, K.Y.; Duman-Scheel, M. Chitosan/interfering RNA nanoparticle mediated gene silencing in disease vector mosquito larvae. J. Vis. Exp. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Almazan, C.; Reyes, E.Z.; Zuniga-Navarrete, F.; Munoz-Garay, C.; Gomez, I.; Evans, A.M.; Likitvivatanavong, S.; Bravo, A.; Gill, S.S.; Soberon, M. Cadherin binding is not a limiting step for Bacillus thuringiensis subsisraelensis Cry4Ba toxicity to Aedes aegypti larvae. Biochem. J. 2012, 443, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.I.; Reyes, E.Z.; Cancino-Rodezno, A.; Bedoya-Perez, L.P.; Caballero-Flores, G.G.; Muriel-Millan, L.F.; Likitvivatanavong, S.; Gill, S.S.; Bravo, A.; Soberon, M. Aedes aegypti alkaline phosphatase ALP1 is a functional receptor of Bacillus thuringiensis Cry4Ba and Cry11Aa toxins. Insect Biochem. Mol. Biol. 2012, 42, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.J.; Oh, M.; Yeo, W.S.; Galvao, K.N.; Jeong, K.C. Underlying mechanism of antimicrobial activity of chitosan microparticles and implications for the treatment of infectious diseases. PLoS ONE 2014, 9, e92723. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Adams, P. Preserving pyrethroids. Bull. World Health Organ. 2014, 92, 158–159. [Google Scholar] [PubMed]
- Powers, A.M.; Olson, K.E.; Higgs, S.; Carlson, J.O.; Beaty, B.J. Intracellular immunization of mosquito cells to LaCrosse virus using a recombinant Sindbis virus vector. Virus Res. 1994, 32, 57–67. [Google Scholar] [CrossRef]
- Gaines, P.J.; Olson, K.E.; Higgs, S.; Powers, A.M.; Beaty, B.J.; Blair, C.D. Pathogen-derived resistance to dengue type 2 virus in mosquito cells by expression of the premembrane coding region of the viral genome. J. Virol. 1996, 70, 2132–2137. [Google Scholar] [PubMed]
- Olson, K.E.; Higgs, S.; Gaines, P.J.; Powers, A.M.; Davis, B.S.; Kamrud, K.I.; Carlson, J.O.; Blair, C.D.; Beaty, B.J. Genetically engineered resistance to dengue-2 virus transmission in mosquitoes. Science 1996, 272, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Higgs, S.; Rayner, J.O.; Olson, K.E.; Davis, B.S.; Beaty, B.J.; Blair, C.D. Engineered resistance in Aedes aegypti to a West African and a South American strain of yellow fever virus. Am. J. Trop. Med. Hyg. 1998, 58, 663–670. [Google Scholar] [PubMed]
- Franz, A.W.; Sanchez-Vargas, I.; Raban, R.R.; Black, W.C.t.; James, A.A.; Olson, K.E. Fitness impact and stability of a transgene conferring resistance to dengue-2 virus following introgression into a genetically diverse Aedes aegypti strain. PLoS Negl. Trop. Dis. 2014, 8, e2833. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.W.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-derived DNA drives mosquito vector tolerance to arboviral infection. Nat. Commun. 2016. [Google Scholar] [CrossRef] [PubMed]
- Myles, K.M.; Wiley, M.R.; Morazzani, E.M.; Adelman, Z.N. Alphavirus-derived small RNAs modulate pathogenesis in disease vector mosquitoes. Proc. Natl. Acad. Sci. USA 2008, 105, 19938–19943. [Google Scholar] [CrossRef] [PubMed]
- Cirimotich, C.M.; Scott, J.C.; Phillips, A.T.; Geiss, B.J.; Olson, K.E. Suppression of RNA interference increases alphavirus replication and virus-associated mortality in Aedes aegypti mosquitoes. BMC Microbiol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Blandin, S.A.; Marois, E.; Levashina, E.A. Antimalarial responses in Anopheles gambiae: From a complement-like protein to a complement-like pathway. Cell Host Microbe 2008, 3, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Garver, L.S.; Bahia, A.C.; Das, S.; Souza-Neto, J.A.; Shiao, J.; Dong, Y.; Dimopoulos, G. Anopheles Imd pathway factors and effectors in infection intensity-dependent anti-Plasmodium action. PLoS Pathog. 2012, 8, e1002737. [Google Scholar] [CrossRef] [PubMed]
- Garver, L.S.; Dong, Y.; Dimopoulos, G. Caspar controls resistance to Plasmodium falciparum in diverse anopheline species. PLoS Pathog. 2009, 5, e1000335. [Google Scholar] [CrossRef] [PubMed]
- Geldhof, P.; Visser, A.; Clark, D.; Saunders, G.; Britton, C.; Gilleard, J.; Berriman, M.; Knox, D. RNA interference in parasitic helminths: Current situation, potential pitfalls and future prospects. Parasitology 2007, 134, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Gallup, J.M.; Day, T.A.; Bartholomay, L.C.; Kimber, M.J. Development of an in vivo RNAi protocol to investigate gene function in the filarial nematode, Brugia malayi. PLoS Pathog. 2010, 6, e1001239. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Higgs, S.; Klingler, K.A.; Vanlandingham, D.L.; Raikhel, A.S. RNA interference-mediated knockdown of a GATA factor reveals a link to anautogeny in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2003, 100, 13374–13379. [Google Scholar] [CrossRef] [PubMed]
- Wynant, N.; Santos, D.; Verdonck, R.; Spit, J.; van Wielendaele, P.; Broeck, J.V. Identification, functional characterization and phylogenetic analysis of double stranded RNA degrading enzymes present in the gut of the desert locust, Schistocerca gregaria. Insect Biochem. Mol. Biol. 2014, 46, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Chen, Y.; Hu, Z.; Hu, M. Testing insecticidal activity of novel chemically synthesized siRNA against Plutella xylostella under laboratory and field conditions. PLoS ONE 2013, 8, e62990. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Yang, X.; Zhang, B.; Zhong, G.; Hu, M. Silencing of Rieske iron-sulfur protein using chemically synthesised siRNA as a potential biopesticide against Plutella xylostella. Pest Manag. Sci. 2011, 67, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Papathanos, P.A.; Windbichler, N.; Menichelli, M.; Burt, A.; Crisanti, A. The vasa regulatory region mediates germline expression and maternal transmission of proteins in the malaria mosquito Anopheles gambiae: A versatile tool for genetic control strategies. BMC Mol. Biol. 2009. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Crisanti, A.; Catteruccia, F. Comparative analysis of DNA vectors at mediating RNAi in Anopheles mosquito cells and larvae. J. Exp. Biol. 2003, 206, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Bugeon, L.; Crisanti, A.; Catteruccia, F. Stable and heritable gene silencing in the malaria vector Anopheles stephensi. Nucleic Acids Res. 2003, 31, e85. [Google Scholar] [CrossRef] [PubMed]
- Adelman, Z.N.; Anderson, M.A.; Morazzani, E.M.; Myles, K.M. A transgenic sensor strain for monitoring the RNAi pathway in the yellow fever mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Liu, B.R.; Dai, Y.H.; Lee, C.Y.; Chan, M.H.; Chen, H.H.; Chiang, H.J.; Lee, H.J. A gene delivery system for insect cells mediated by arginine-rich cell-penetrating peptides. Gene 2012, 493, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Ramesh Kumar, D.; Kumar, P.S.; Gandhi, M.R.; Al-Dhabi, N.A.; Paulraj, M.G.; Ignacimuthu, S. Delivery of chitosan/dsRNA nanoparticles for silencing of wing development vestigial (vg) gene in Aedes aegypti mosquitoes. Int. J. Biol. Macromol. 2016, 86, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Meade, B.R.; Dowdy, S.F. Enhancing the cellular uptake of siRNA duplexes following noncovalent packaging with protein transduction domain peptides. Adv. Drug Deliv. Rev. 2008, 60, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Meade, B.R.; Dowdy, S.F. Exogenous siRNA delivery using peptide transduction domains/cell penetrating peptides. Adv. Drug Deliv. Rev. 2007, 59, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Airs, P.M.; Bartholomay, L.C.; Department of Pathobiological Sciences, University of Wisconsin-Madison, Madison, WI, USA. Unpublished work. 2016.
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Olson, K.; Higgs, S.; Beaty, B. Molecular genetic manipulation of mosquito vectors. Annu. Rev. Entomol. 1995, 40, 359–388. [Google Scholar] [CrossRef] [PubMed]
- Levis, R.; Huang, H.; Schlesinger, S. Engineered defective interfering RNAs of Sindbis virus express bacterial chloramphenicol acetyltransferase in avian cells. Proc. Natl. Acad. Sci. USA 1987, 84, 4811–4815. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Hansen, I.A.; Raikhel, A.S. Nutritional regulation of vitellogenesis in mosquitoes: Implications for anautogeny. Insect Biochem. Mol. Biol. 2005, 35, 661–675. [Google Scholar] [CrossRef] [PubMed]
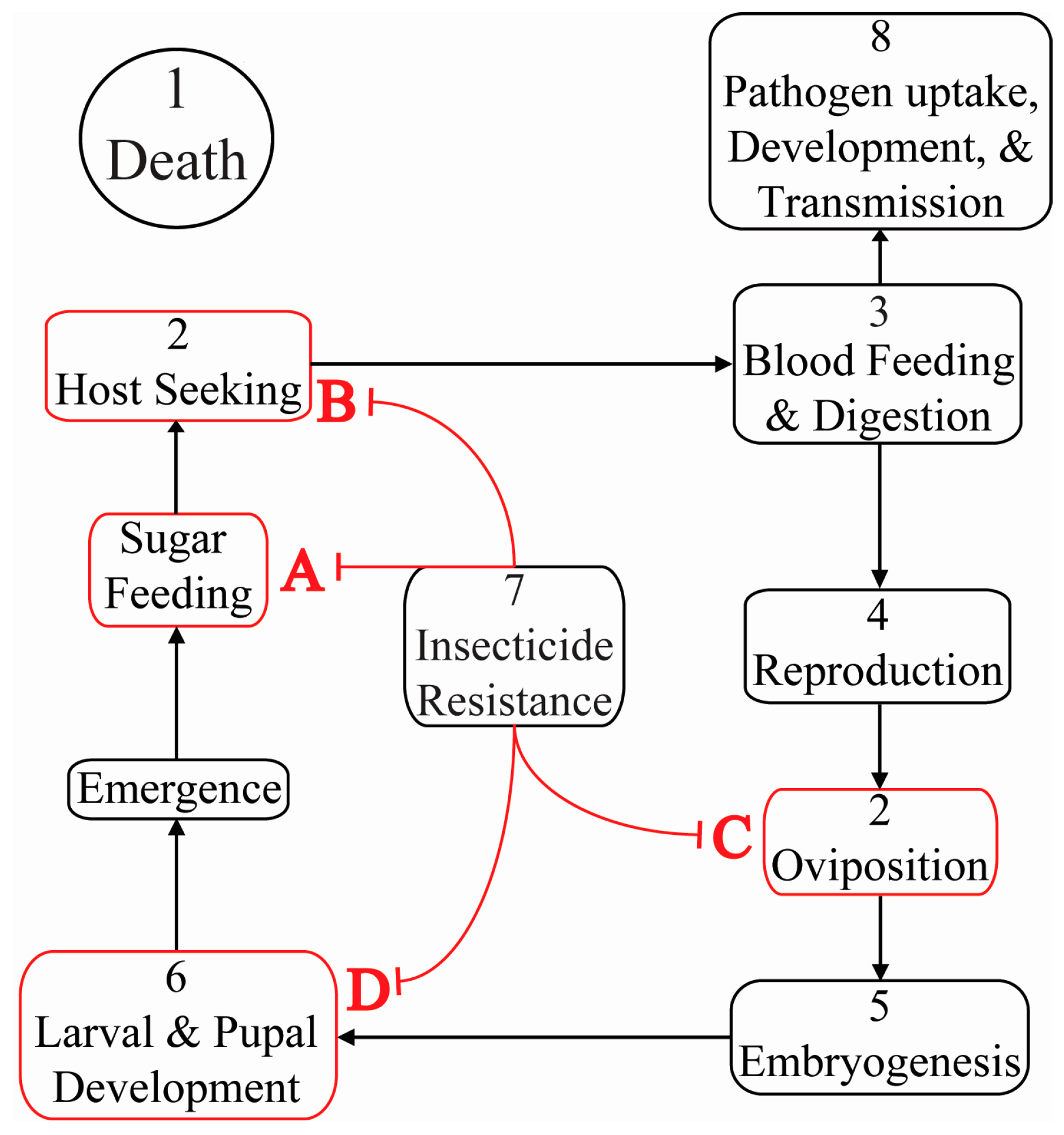
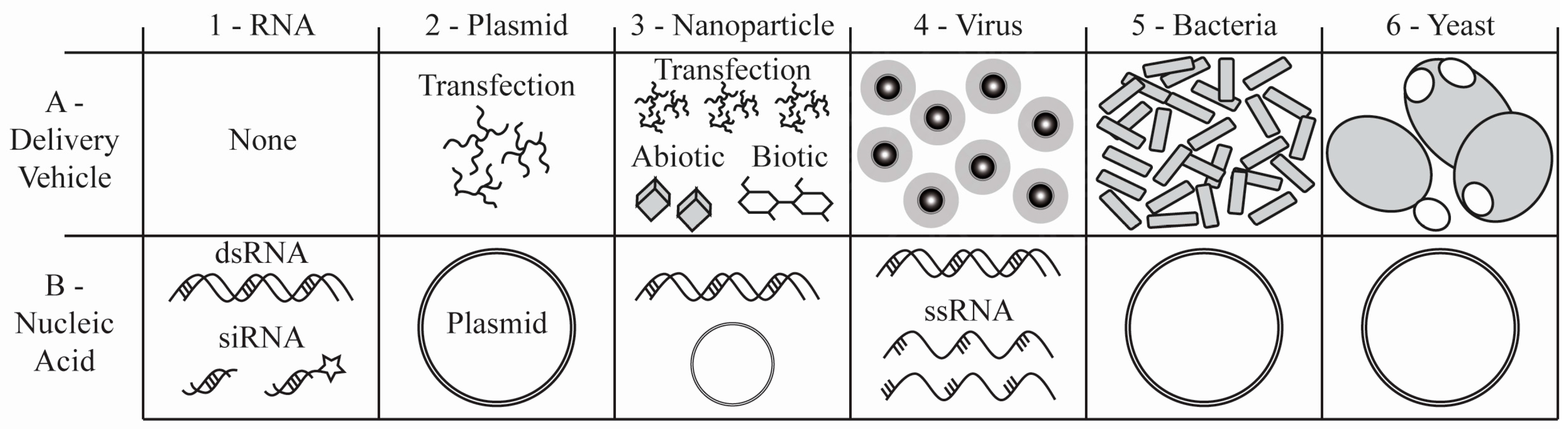

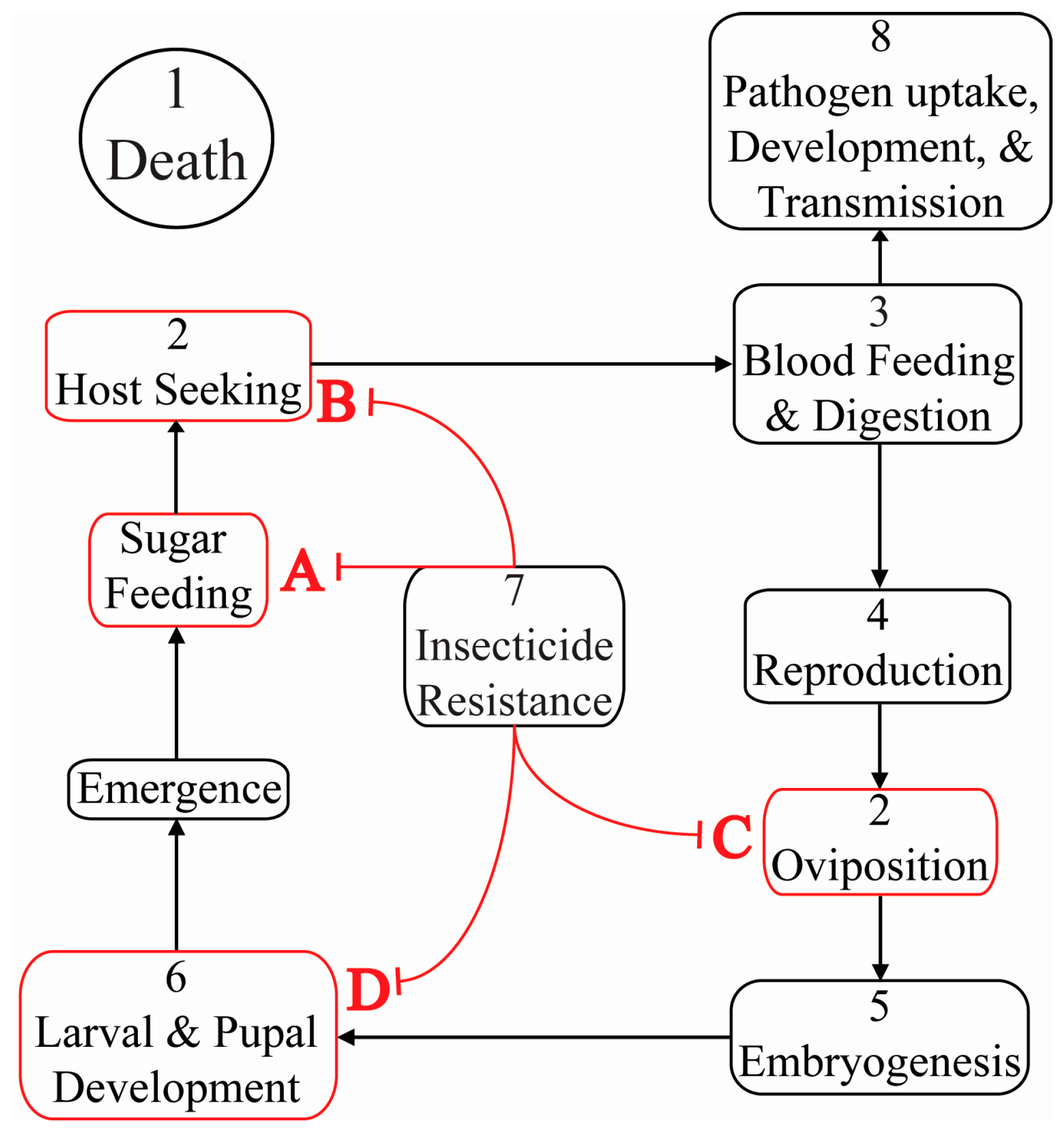{kind=link}
{kind=link}
| Legend * | Function | Gene of Interest | Accession | Outcome | Species | Reference |
|---|---|---|---|---|---|---|
| 1 | Death | Inhibitor of Apoptosis 1 | AAEL009074 | Death | Ae. aegypti | [10,52,53,54,55] |
| 2 | Olfaction & Sensation | Gustatory receptors 1 & 3 | AAEL002380, AAEL010058 | Inability to detect CO2 | Ae. aegypti | [15] |
| Odorant binding protein 1 | CPIJ007604 | Reduced oviposition attractant sensing | Cx. quinquefasciatus | [16] | ||
| 3 | Blood Feeding | Aegyptin | AGAP009974 | Diminished blood feeding success | Ae. aegypti | [17] |
| 4 | Reproduction | Ovary ecdysteroidogenic hormone receptor | AAEL001915 | Diminished egg development | Ae. aegypti | [56] |
| zero population growth | AGAP006241 | Spermless males | An. gambiae | [14] | ||
| 5 | Embryogenesis | Frazzled | AAEL014592 | Malformed ventral nerve cord | Ae. aegypti | [57] |
| 6 | Larval & Pupal Development | Chitin synthase 1 | AAEL002718 | Disrupted peritrophic matrix | An. gambiae | [49] |
| Morphogenesis | Prophenoloxidase III | AY487171.1 | Malformed pharate adult cuticle | Am. subalbatus | [19] | |
| 7 | Pesticide Resistance | Protease m1 zinc metalloprotease | CPIJ012471 | Death by deltamethrin susceptibility | Cx. pipiens | [58] |
| 8 | Pathogen Uptake, Development & Transmission | Caspar | AGAP006473 | Suppresses malaria parasite numbers in the midgut | An. gambiae | [59] |
| Legend * | (A) Delivery Vehicle | (B) Nucleic Acid | RNAi Target | Delivery Route | Reference |
|---|---|---|---|---|---|
| 1 | None | dsRNA | vATPase A | Adult per os | [11] |
| None | dsRNA | P-glycoprotein | Larval per os | [127] | |
| None | siRNA | Semaphorin A | Embryo Injection | [117] | |
| 2 | None | pMOS-dsRED plasmid | Aegyptin | Embryo Injection | [17] |
| 3 | Liposome (Effectene®) | dsRNA | Caspase 1 | Larval per os | [123] |
| Chitosan | dsRNA | Chitin synthase 1 | Larval per os | [49] | |
| 4 | Sindbis virus | Long hairpin RNA | GATA factor | Adult Injection | [149] |
| Densovirus | Short hairpin RNA | vATPase A | Transfection (C6/36 cells) | [81] | |
| 5 | E. coli | Long hairpin RNA | AAEL001684 | Larval per os of E. coli | [13] |
| 6 | P. pastoris | Long hairpin RNA | JH acid methyl transferase | Larval per os of P. pastoris | [27] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Airs, P.M.; Bartholomay, L.C. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects 2017, 8, 4. https://doi.org/10.3390/insects8010004
Airs PM, Bartholomay LC. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects. 2017; 8(1):4. https://doi.org/10.3390/insects8010004
Chicago/Turabian StyleAirs, Paul M., and Lyric C. Bartholomay. 2017. "RNA Interference for Mosquito and Mosquito-Borne Disease Control" Insects 8, no. 1: 4. https://doi.org/10.3390/insects8010004
APA StyleAirs, P. M., & Bartholomay, L. C. (2017). RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects, 8(1), 4. https://doi.org/10.3390/insects8010004