
A Review of the Tawny Crazy Ant, Nylanderia fulva, an Emergent Ant Invader in the Southern United States: Is Biological Control a Feasible Management Option?

Abstract
:1. Introduction
2. Biology
3. Taxonomy
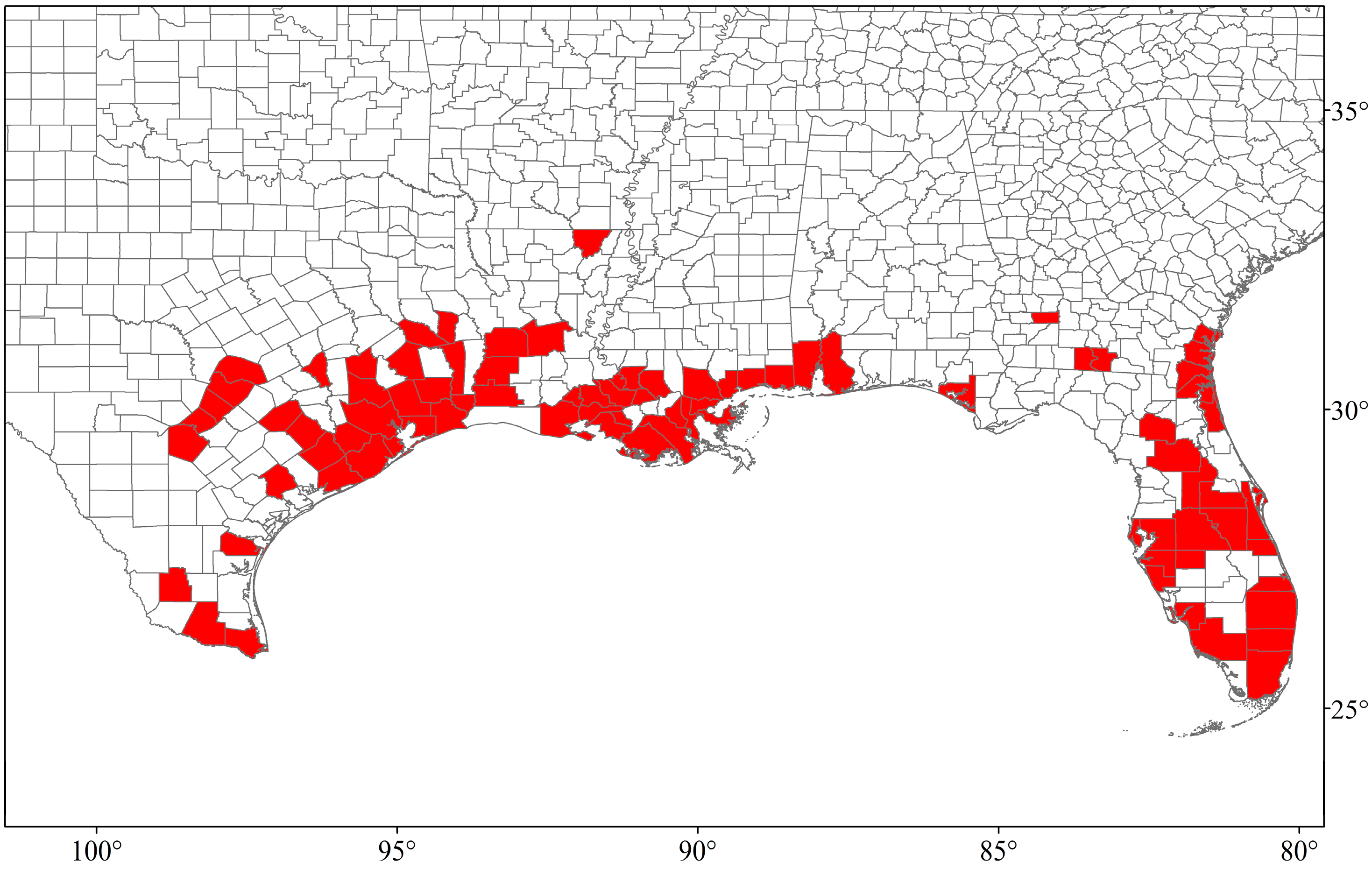
4. Distribution and Spread
5. Ecology
6. Ecological Impact
7. Economic Impacts and Management
8. Biological Control Candidates
9. Conclusions and Future Research
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Meyers, J.M. Identification, Distribution and Control of an Invasive Pest Ant, Paratrechina sp. (Hymenoptera: Formicidae), in Texas. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2008. [Google Scholar]
- LeBrun, E.G.; Abbott, J.; Gilbert, L.E. Imported crazy ant displaces imported fire ant, reduces and homogenizes grassland ant and arthropod assemblages. Biol. Invasions 2013, 15, 2429–2442. [Google Scholar] [CrossRef]
- Lard, C.; Willis, D.B.; Salin, V.; Robison, S. Economic assessments of red imported fire ant on Texas’ urban and agricultural sectors. Southwest Entomol. 2002, 25, 123–137. [Google Scholar]
- McDonald, D.L. Investigation of an Invasive Ant Species: Nylanderia fulva Colony Extraction, Management, Diet Preference, Fecundity, and Mechanical Vector Potential. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2012. [Google Scholar]
- Sharma, S.; Oi, D.H.; Buss, E.A. Honeydew-producing hemipterans in Florida associated with Nylanderia fulva (Hymenoptera: Formicidae), an invasive crazy ant. Fla. Entomol. 2013, 96, 538–547. [Google Scholar] [CrossRef]
- Arcila, A.M.; Gómez, L.A.; Ulloa-Chacón, P. Immature development and colony growth of crazy ant Paratrechina fulva under laboratory conditions (Hymenoptera: Formicidae). Sociobiology 2002, 39, 307–322. [Google Scholar]
- Zenner-Polanía, I. Biological aspects of the “hormiga loca”, Paratrechina (Nylanderia) fulva (Mayr), in Colombia. In Applied Myrmecology: A World Perspective; Meer, R.K.V., Jaffe, K., Cedeno, A., Eds.; Westview Press: Boulder, CO, USA, 1990; pp. 290–297. [Google Scholar]
- McDonald, D.L.; Texas Research Institute for Environemental Studies, Sam Houston State University, Huntsville, TX, USA. Personal communication, 2016.
- Brady, S.G.; Schultz, T.R.; Fisher, B.L.; Ward, P.S. Evaluating alternative hypothese for the early evolution and diversification of ants. Proc. Natl. Acad. Sci. USA 2006, 103, 18172–18177. [Google Scholar] [CrossRef] [PubMed]
- LaPolla, J.; Brady, S.G.; Shattuck, S.O. Monograph of Nylanderia (Hymenoptera: Formicidae) of the world: An introduction to the systematics and biology of the genus. Zootaxa 2011, 3110, 1–9. [Google Scholar]
- LaPolla, J.; Brady, S.G.; Shattuck, S.O. Phylogeny and taxonomy of the Prenolepis genus-group of ants (Hymenoptera: Formicidae). Syst. Entomol. 2010, 35, 118–131. [Google Scholar] [CrossRef]
- Gotzek, D.; Brady, S.G.; Kallal, R.J.; LaPolla, J.S. The importance of using multiple approaches for identifying emerging invasive species: The case of the Rasberry crazy ant in the United States. PLoS ONE 2012, 7, e45314. [Google Scholar] [CrossRef] [PubMed]
- MacGown, J.; Layton, B. The invasive Rasberry crazy ant, Nylanderia sp. Near pubens (Hymenoptera: Formicidae), reported from Mississippi. Midsouth Entomol. 2010, 3, 44–47. [Google Scholar]
- Trager, J.C. A revision of the genus Paratrechina of the continental United States. Sociobiology 1984, 9, 51–162. [Google Scholar]
- Kallal, R.J.; Lapolla, J.S. Monograph of Nylanderia (Hymenoptera: Formicidae) of the world, part II: Nylanderia in the Nearctic. Zootaxa 2012, 3508, e64. [Google Scholar]
- Zhao, L.; Chen, J.; Jones, W.A.; Oi, D.H.; Drees, B.M. Molecular comparisons suggest Caribbean crazy ant from Florida and Rasberry crazy ant from Texas (Hymenoptera: Formicidae: Nylanderia) are the same species. Environ. Entomol. 2012, 41, 1008–1018. [Google Scholar] [CrossRef]
- Entomological Society of America. Common Names of Insects Database. 2016. Available online: http://www.entsoc.org/common-names (accessed on 5 December 2016).
- Kumar, S.; LeBrun, E.G.; Stohlgren, T.J.; Stabach, J.A.; McDonald, D.L.; Oi, D.H.; LaPolla, J.S. Evidence of niche shift and global invasion potential of the tawny crazy ant, Nylanderia fulva. Ecol. Evol. 2015, 5, 4628–4641. [Google Scholar] [CrossRef] [PubMed]
- Wetterer, J.K.; Keularts, J.L. Population explosion of the hairy crazy ant, Paratrechina pubens (Hymenoptera: Formicidae), on St. Croix, US virgin islands. Fla. Entomol. 2008, 91, 423–427. [Google Scholar] [CrossRef]
- Hill, S.K.; Baldwin, R.W.; Pereira, R.M.; Koehler, P.G. Tawny Crazy Ant. Available online: http://edis.ifas.ufl.edu/pdffiles/IN/IN107600.pdf (accessed on 30 November 2016).
- Graham, L.C.; University of Florida, Gainesville, FL, USA; Buss, L.; Auburn University, Auburn, AL, USA; Henderson, G.; Louisiana State University, Baton Rouge, LA, USA; Suiter, D.R.; University of Georgia, Athens, GA, USA; Puckett, R.T.; Texas A&M University, College Station, TX, USA; Layton, B.; Mississippi State University, Starkville, MS, USA. Personal communication, 2016.
- Bentley, M.; Hahn, D.; Oi, F. The thermal breadth of Nylanderia fulva (Hymenoptera: Formicidae) is narrower than that of Solenopsis invicta at three thermal ramping rates: 1.0, 0.12, and 0.06 °C·min−1. Environ. Entomol. 2016. [Google Scholar] [CrossRef] [PubMed]
- USDA. Species Profiles: Red Imported Fire Ant. National Invasive Species Information Center, 2016. Available online: https://www.invasivespeciesinfo.gov/animals/rifa.shtml (accessed on 5 December 2016). [Google Scholar]
- LeBrun, E.G.; Jones, N.T.; Gilbert, L.E. Chemical warfare among invaders: A detoxification interaction facilitates an ant invasion. Science 2014, 343, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Horn, K. Examining Competitive Interaction between Rasberry Crazy Ants (Paratrechina sp. Nr. Pubens) and Red Imported Fire Ants (Solenopsis invicta) Using Laboratory and Field Studies. Master’s Thesis, Rice University, Houston, TX, USA, 2009. [Google Scholar]
- Chen, J.; Rashid, T.; Feng, G.; Zhao, L.; Oi, D. Defensive chemicals of tawny crazy ants, Nylanderia fulva (Hymenoptera: Formicidae) and their toxicity to red imported fire ants, Solenopsis invicta (Hymenoptera: Formicidae). Toxicon 2013, 76, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Horn, K.C.; Eubanks, M.D.; Siemann, E. The effect of diet and opponent size on aggressive interactions involving Caribbean crazy ants (Nylanderia fulva). PLoS ONE 2013, 8, e66912. [Google Scholar] [CrossRef] [PubMed]
- Zenner-Polania, I. Impact of Paratrechina fulva on other ant species. In Exotic Ants: Biology, Impact and Control of Introduced Species; Williams, D.F., Ed.; Westview Press: Boulder, CO, USA, 1994; pp. 121–132. [Google Scholar]
- Lard, C.; Schmidt, J.; Morris, B.; Estes, L.; Ryan, C.; Berquist, D. An Economic Impact of Imported Fire Ants in the United States of America; Texas A&M University: College Station, TX, USA, 2006; Available online: https://articles.extension.org/sites/default/files/Copy%20of%20the%20National%20Study.pdf (accessed on 5 December 2016).
- Oi, F.; Calibeo, D.; Paige, J.; Bentley, M. Integrated Pest Management (IPM) of the Tawny Crazy Ant, Nylanderia fulva (Mayr); ENY-2006 (IN889); University of Florida, Institute of Food and Agricultural Sciences Extension: Gainesville, FL, USA, 2016; Available online: edis.ifas.ufl.edu/pdffiles/in/in88900.pdf (accessed on 5 December 2016).
- Sanchez-Bayo, F.; Goka, K. Pesticide residues and bees–a risk assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.D.; Pasquero, M.A. Illustrated key to Pseudacteon decapitating flies (Diptera: Phoridae) that attack Solenopsis saevissima complex fire ants in South America. Fla. Entomol. 2002, 84, 691–699. [Google Scholar] [CrossRef]
- Pasquero, M.A.; Departamento de Ecologia e Biologia Evolutiva, Universidade Federal de São Carlos, São Carlos, São Paulo, Brazil. Personal communication, 2016.
- Brown, B.V.; Schneider, S.A.; LaPolla, J.S. A new north American species of Pseudacteon (Diptera: Phoridae), parasitic on Nylanderia arenivaga (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 2011, 104, 37–38. [Google Scholar] [CrossRef]
- Porter, S.D. Host specificity and risk assessment of releasing the decapitating fly Pseudacteon curvatus as a classical biocontrol agent for imported fire ants. Biol. Control 2000, 19, 35–47. [Google Scholar] [CrossRef]
- Graham, L.C.; Porter, S.D.; Pereira, R.M.; Dorough, H.D.; Kelley, A.T. Field releases of the decapitating fly Pseudacteon curvatus (Diptera: Phoridae) for control of imported fire ants (Hymenoptera: Formicidae) in Alabama, Florida, and Tennessee. Fla. Entomol. 2003, 86, 334–339. [Google Scholar] [CrossRef]
- Plowes, R.M.; Becnel, J.J.; LeBrun, E.G.; Oi, D.H.; Valles, S.M.; Jones, N.T.; Gilbert, L.E. Myrmecomorba nylanderiae gen. et sp. nov., a microsporidian parasite of the tawny crazy ant Nylanderia fulva. J. Invertebr. Pathol. 2015, 129, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Knell, J.; Allen, G.; Hazard, E. Light and electron microscope study of Thelohania solenopsae n. Sp. (Microsporida: Protozoa) in the red imported fire ant, Solenopsis invicta. J. Invertebr. Pathol. 1977, 29, 192–200. [Google Scholar] [CrossRef]
- Oi, D.H.; Williams, D.F. Impact of Thelohania solenopsae (microsporidia: Thelohaniidae) on polygyne colonies of red imported fire ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2002, 95, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Valles, S.M.; Oi, D.H.; Becnel, J.J.; Wetterer, J.K.; LaPolla, J.S.; Firth, A.E. Isolation and characterization of Nylanderia fulva virus 1, a positive-sense, single-stranded RNA virus infecting the tawny crazy ant, Nylanderia fulva. Virol. J. 2016, 496, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Van Lenteren, J.; Bale, J.; Bigler, F.; Hokkanen, H.; Loomans, A. Assessing risks of releasing exotic biological control agents of arthropod pests. Annu. Rev. Entomol. 2006, 51, 609–634. [Google Scholar] [CrossRef] [PubMed]
- Babendreier, D.; Bigler, F.; Kuhlmann, U. Methods used to assess non-target effects of invertebrate biological control agents of arthropod pests. BioControl 2005, 50, 821–870. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | Native/Invasive | Range | Overlap with N. fulva |
|---|---|---|---|
| N. arenivaga | Native | Eastern and central U.S. | Yes |
| N. austroccidua | Native | Southwest U.S., Mexico to Costa Rica | Yes |
| N. bruesii | Native | Southwest U.S., northwestern Mexico | Yes |
| N. concinna | Native | Eastern U.S. | Yes |
| N. faisonensis | Native | Eastern U.S. | Yes |
| N. hystrix | Native | Southwestern U.S. | Yes |
| N. magnella | Native | Southwestern U.S. | Yes |
| N. parvula | Native | Eastern U.S. | Yes |
| N. phantasma | Native | Southeastern U.S. | Yes |
| N. querna | Native | Central U.S. | Yes |
| N. terricola | Native | U.S. and northern Mexico | Yes |
| N. trageri | Native | Central U.S. | Yes |
| N. vividula | Native | U.S. and northern Mexico | Yes |
| N. wojciki | Native | Southeastern U.S. | Yes |
| N. bourbonica | Introduced | Florida | Yes |
| N. flavipes | Introduced | Northeastern U.S. | No |
| N. guatemalensis | Introduced | Florida | Yes |
| N. pubens | Introduced | Florida | Yes |
| N. steinheili | Introduced | Florida | Yes |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Moshman, L.; Kraus, E.C.; Wilson, B.E.; Acharya, N.; Diaz, R. A Review of the Tawny Crazy Ant, Nylanderia fulva, an Emergent Ant Invader in the Southern United States: Is Biological Control a Feasible Management Option? Insects 2016, 7, 77. https://doi.org/10.3390/insects7040077
Wang Z, Moshman L, Kraus EC, Wilson BE, Acharya N, Diaz R. A Review of the Tawny Crazy Ant, Nylanderia fulva, an Emergent Ant Invader in the Southern United States: Is Biological Control a Feasible Management Option? Insects. 2016; 7(4):77. https://doi.org/10.3390/insects7040077
Chicago/Turabian StyleWang, Zinan, Lori Moshman, Emily C. Kraus, Blake E. Wilson, Namoona Acharya, and Rodrigo Diaz. 2016. "A Review of the Tawny Crazy Ant, Nylanderia fulva, an Emergent Ant Invader in the Southern United States: Is Biological Control a Feasible Management Option?" Insects 7, no. 4: 77. https://doi.org/10.3390/insects7040077
APA StyleWang, Z., Moshman, L., Kraus, E. C., Wilson, B. E., Acharya, N., & Diaz, R. (2016). A Review of the Tawny Crazy Ant, Nylanderia fulva, an Emergent Ant Invader in the Southern United States: Is Biological Control a Feasible Management Option? Insects, 7(4), 77. https://doi.org/10.3390/insects7040077