
Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to Imidacloprid, Thiamethoxam, Dinotefuran and Flupyradifurone in South Florida

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Field Colonies
2.2. Bioassay Technique
2.3. Bioassay Sampling
2.4. Statistical Analysis
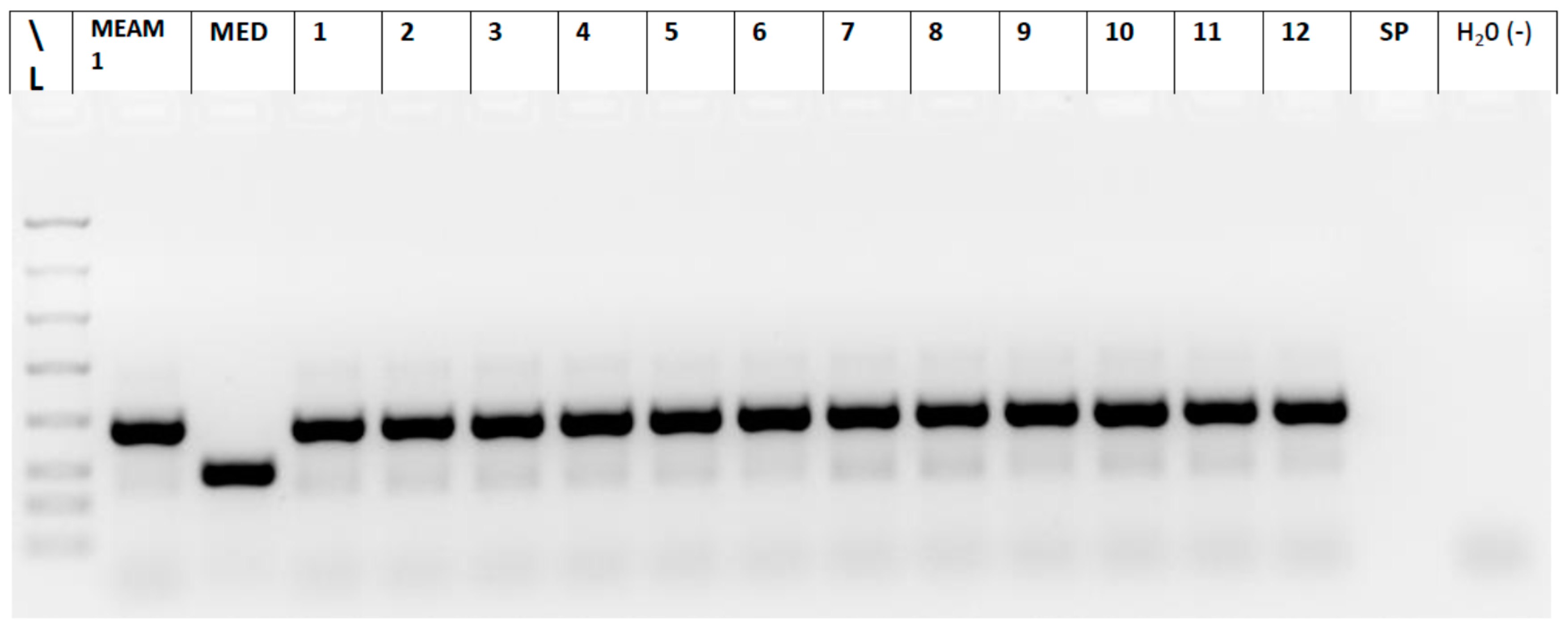
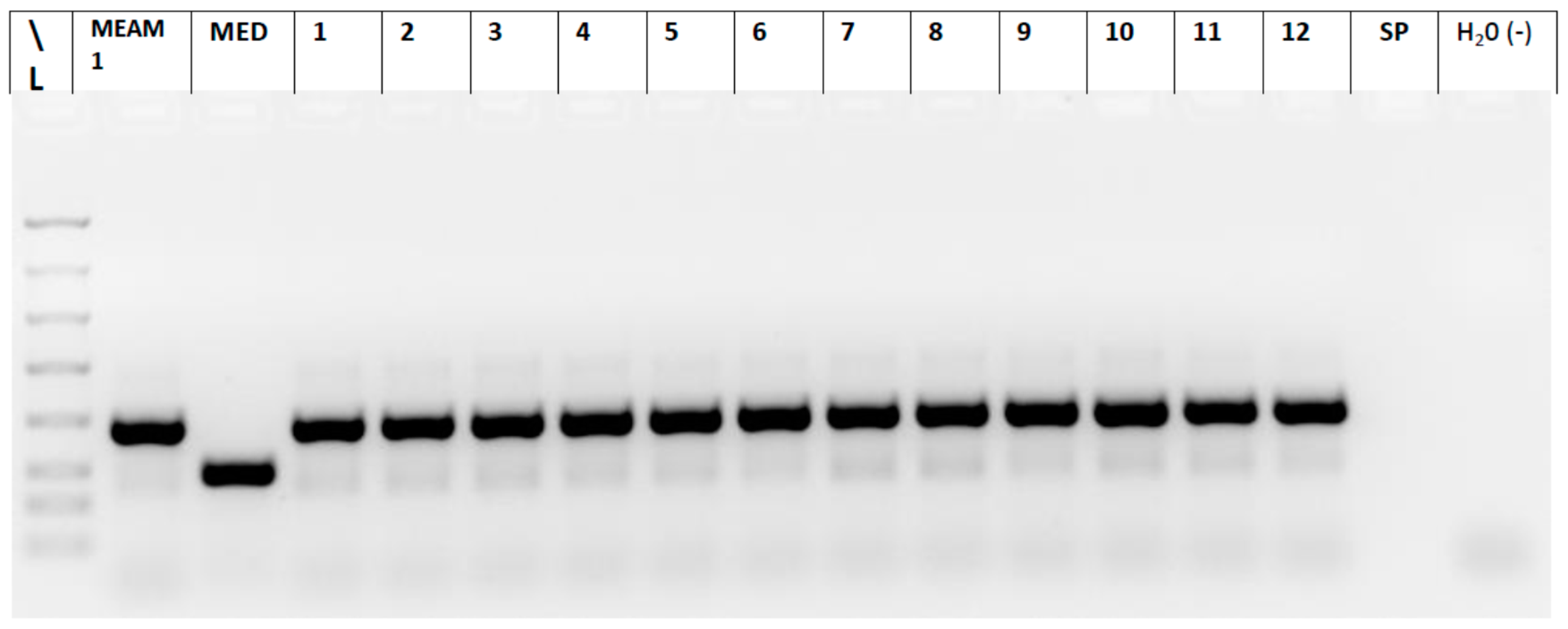
2.5. Whitefly Biotype Determination
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DNA | Deoxyribonucleic acid |
| GCREC | Gulf Coast Research and Education Center |
| LC | Lethal concentration |
| MEAM1 | Middle East Asia Minor group of Bemisia tabaci (commonly known as biotype B) |
| MED | Mediterranean group of Bemisia tabaci (commonly known as biotype Q) |
| PCR | Polymerase chain reaction |
| TYLCV | Tomato yellow leaf curl virus |
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | linear dichroism |
References
- Thompson, W.M.O. Introduction: Whiteflies, geminiviruses and recent events. In The Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) Interaction with Geminivirus-Infected Host Plants; Springer: Berlin, Germany, 2011; pp. 1–13. [Google Scholar]
- Polston, J.E.; de Barro, P.; Boykin, L.M. Transmission specificities of plant viruses with the newly identified species of the Bemisia tabaci species complex. Pest Manage. Sci. 2014, 70, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.J.; Stansly, P.A.; Polston, J.E. Expressions of plant damage by Bemisia. In Bemisia: 1995, Taxonomy, Biology, Damage, Control and Management; Andover: Hants, UK, 1996. [Google Scholar]
- Mosler, M.; Aerts, M.J.; Nesheim, O.N. Florida crop/pest management profiles: Tomatoes. Available online: http://edis.ifas.ufl.edu/pi039 (accessed on 14 July 2016).
- Levy, D.; Lapidot, M. Effect of plant age at inoculation on expression of genetic resistance to tomato yellow leaf curl virus. Arch. Virol. 2008, 153, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Vegetables annual summary. Available online: http://usda.mannlib.cornell.edu/MannUsda/viewDocumentInfo.do?documentID=1183files/97/viewDocumentInfo.html (accessed on 14 July 2016).
- Polston, J.E.; McGovern, R.J.; Brown, L.G. Introduction of tomato yellow leaf curl virus in Florida and implications for the spread of this and other geminiviruses of tomato. Plant Dis. 1999, 83, 984–988. [Google Scholar] [CrossRef]
- Adkins, S.; Webster, C.G.; Kousik, C.S.; Webb, S.E.; Roberts, P.D.; Stansly, P.A.; Turechek, W.W. Ecology and management of whitefly-transmitted viruses of vegetable crops in Florida. Virus Res. 2011, 159, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Stansly, P.A.; Smith, H.A.; Seal, D.R.; McAvoy, E.; Polston, J.E.; Gilreath, P.R.; Schuster, D.J. Management of whiteflies, whitefly-vectored plant virus, and insecticide resistance for vegetable production in southern Florida. Available online: http://edis.ifas.ufl.edu/in695files/102/in695.html (accessed on 19 May 2016).
- Smith, H.A.; Nagle, C.A. Combining novel modes of action for early-season management of Bemisia tabaci (Hemiptera: Aleyrodidae) and Tomato Yellow Leaf Curl Virus in tomato. Fla. Entomol. 2014, 97, 1750–1765. [Google Scholar] [CrossRef]
- Dittrich, V.; Uk, S.; Ernst, G.H. Chemical control and insecticide resistance of whiteflies. Whiteflies 1990, 263–286. [Google Scholar]
- Denholm, I.; Cahill, M.; Byrne, F.J.; Devonshire, A.L. Progress with documenting and combating insecticide resistance in Bemisia. In Bemisia: 1995, Taxonomy, Biology, Damage, Control and Management; Andover: Hants, UK, 1996. [Google Scholar]
- Castle, S.J.; Palumbo, J.C.; Prabhaker, N.; Horowitz, A.R.; Denholm, I.; Stansly, P.A.; Naranjo, S.E. Ecological Determinants of Bemisia tabaci Resistance to Insecticides. In Bemisia: Bionomics and Management of a Global Pest; Springer: Dordrecht, The Netherlands, 2009; pp. 423–465. [Google Scholar]
- Cahill, M.; Gorman, K.; Day, S.; Denholm, I.; Elbert, A.; Nauen, R. Baseline determination and detection of resistance to imidacloprid in Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 1996, 86, 343–349. [Google Scholar] [CrossRef]
- Prabhaker, N.; Castle, S.; Henneberry, T.J.; Toscano, N.C. Assessment of cross-resistance potential to neonicotinoid insecticides in Bemisia tabaci (Hemiptera: Aleyrodidae). Bull. Entomol. Res. 2005, 95, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Castle, S.J.; Prabhaker, N. Monitoring changes in Bemisia tabaci (Hemiptera: Aleyrodidae) susceptibility to neonicotinoid insecticides in Arizona and California. J. Econ. Entomol. 2013, 106, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.J.; Mann, R.S.; Toapanta, M.; Cordero, R.; Thompson, S.; Cyman, S.; Shurtleff, A.; Morris, R.F. Monitoring neonicotinoid resistance in biotype b of Bemisia tabaci in Florida. Pest Manag. Sci. 2010, 66, 186–195. [Google Scholar]
- Caballero, R.; Cyman, S.; Schuster, D.J. Monitoring insecticide resistance in biotype B of Bemisia tabaci (Hemiptera: Aleyrodidae) in Florida. Fla. Entomol. 2014, 96, 1243–1256. [Google Scholar] [CrossRef]
- McKenzie, C.L.; Hodges, G.; Osborne, L.S.; Byrne, F.J.; Shatters, R.G. Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) biotypes in Florida—Investigating the Q invasion. J. Econ. Entomol. 2009, 102, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.R.; Gorman, K.; Ross, G.; Denholm, I. Inheritance of pyriproxyfen resistance in the whitefly, Bemisia tabaci (Q biotype). Arch. Insect Biochem. Physiol. 2003, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.R.; Kontsedalov, S.; Ishaaya, I. Dynamics of resistance to the neonicotinoids acetamiprid and thiamethoxam in Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 2004, 97, 2051–2056. [Google Scholar] [CrossRef] [PubMed]
- Schierow, L.J.; Johnson, R.; Corn, M.L. Bee health: The role of pesticides. Available online: https://www.fas.org/sgp/crs/misc/R42855.pdf (accessed on 15 May 2016).
- Nauen, R.; Jeschke, P.; Velten, R.; Beck, M.E.; Ebbinghaus-Kintscher, U.; Thielert, W.; Wolfel, K.; Haas, M.; Kunz, K.; Raupach, G. Flupyradifurone: A brief profile of a new butenolide insecticide. Pest Manag. Sci. 2014, 71, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.A.; Giurcanu, M.C. Residual effects of new insecticides on egg and nymph densities of Bemisia tabaci (Hemiptera: Aleyrodidae). Fla. Entomol. 2013, 504–511. [Google Scholar] [CrossRef]
- Smith, H.A.; Giurcanu, M.C. New insecticides for management of Tomato Yellow Leaf Curl, a virus vectored by the silverleaf whitefly, Bemisia tabaci. J. Insect Sci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Insecticide Resistance Action Committee. IRAC susceptibility test methods series. Available online: http://www.irac-online.org/content/uploads/Method_016_v3_june09.pdf (accessed 1 September 2016).
- SPSS Statistics for Windows, 22.0; IBM Corp.: Armonk, NY, USA, 2013.
- Shatters, R.G.; Powell, C.A.; Boykin, L.M.; Liansheng, H.; McKenzie, C.L. Improved DNA barcoding method for Bemisia tabaci and related Aleyrodidae: Development of universal and Bemisia tabaci biotype-specific mitochondrial cytochrome c oxidase I polymerase chain reaction primers. J. Econ. Entomol. 2009, 102, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.J.; Stansly, P.A.; Gilreath, P.R.; Polston, J.E. Management of Bemisia, TYLCV, and insecticide resistance in Florida vegetables. J. Insect Sci. 2008, 8, 43–44. [Google Scholar]
- Nauen, R.; Stumpf, N.; Elbert, A. Toxicological and mechanistic studies on neonicotinoid cross resistance in Q-type Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2002, 58, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhu, C.; Feng, Y.; Li, W.; Shao, X.; Xu, Z.; Cheng, J.; Li, Z. Computational insights into the different resistance mechanism of imidacloprid versus dinotefuran in Bemisia tabaci. J. Agric. Food Chem. 2016, 64, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Qiong, R.; Yonghua, X.; Chen, L.; Hongyu, Z.; Jones, C.M.; Devine, G.J.; Gorman, K.; Denhom, I. Characterisation of neonicotinoid and pymetrozine resistance in strains of Bemisia tabaci from China. J. Integr. Agric. 2012, 11, 321–326. [Google Scholar]
- Feng, Y.; Wu, Q.; Wang, S.; Chang, X.; Xie, W.; Xu, B.; Zhang, Y. Cross-resistance study and biochemical mechanisms of thiamethoxam resistance in B-biotype Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manag. Sci. 2010, 66, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Gorman, K.; Slater, R.; Blande, J.D.; Clarke, A.; Wren, J.; McCaffery, A.; Denholm, I. Cross-resistance relationships between neonicotinoids and pymetrozine in Bemisia tabaci (Hemiptera: Aleyrodidae). Pest Manage. Sci. 2010, 66, 1186–1190. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Population Code 1 | Mortality | Resistance Ratio 4 | ||
|---|---|---|---|---|
| No. Tested 2 | Slope ± SE | LC50 (95% CL) 3 | ||
| Laboratory | 492 | 0.564 ± 0.054 | 0.131 (0.018–0.457) | - |
| Hendry3 | 465 | 0.828 ± 0.058 | 0.901 (0.411–1.689) | 6.88 |
| Manatee1 | 412 | 0.481 ± 0.040 | 0.995 (0.263–2.727) | 7.60 |
| Manatee4 | 234 | 0.457 ± 0.047 | 1.349 (0.245–5.067) | 10.30 |
| Hillsborough3 | 449 | 0.522 ± 0.041 | 1.584 (0.502–3.881) | 12.09 |
| IndianRiver-Emilia | 292 | 0.570 ± 0.045 | 1.601 (0.450–4.651) | 12.22 |
| Hendry2 | 371 | 0.705 ± 0.063 | 1.721 (0.670–3.516) | 13.14 |
| Manatee3 | 570 | 0.620 ± 0.033 | 2.055 (0.947–4.168) | 15.69 |
| Hillsborough-Emilia | 542 | 0.641 ± 0.032 | 2.408 (1.249–4.431) | 18.38 |
| Manatee2 | 491 | 0.502 ± 0.031 | 2.629 (1.087–5.913) | 20.07 |
| IndianRiver-Squash | 795 | 0.566 ± 0.026 | 4.112 (2.133–7.690) | 31.39 |
| Hillsborough4 | 235 | 0.460 ± 0.045 | 4.677 (1.118–16.650) | 35.70 |
| Hillsborough1 | 322 | 1.216 ± 0.072 | 5.326 (3.143–8.768) | 40.66 |
| Miami-Dade1 | 526 | 0.812 ± 0.047 | 9.098 (5.543–14.905) | 69.45 |
| IndianRiver1 | 373 | 0.707 ± 0.057 | 9.406 (4.700–18.968) | 71.80 |
| Hillsborough2 | 485 | 0.821 ± 0.052 | 10.590 (5.779–19.783) | 80.84 |
| Miami-Dade3 | 213 | 0.505 ± 0.045 | 11.994 (3.166–46.568) | 91.56 |
| Hendry1 | 581 | 0.749 ± 0.043 | 12.919 (8.089–20.894) | 98.62 |
| Hillsborough5 | 228 | 0.477 ± 0.044 | 21.524 (7.263–70.000) | 164.31 |
| Miami-Dade2 | 363 | 0.530 ± 0.052 | 24.952 (11.519–60.435) | 190.47 |
| Population Code 1 | Mortality | Resistance Ratio 4 | ||
|---|---|---|---|---|
| No. Tested 2 | Slope ± SE | LC50 (95% CL) 3 | ||
| Laboratory | 506 | 0.564 ± 0.054 | 0.139 (0.021–0.460) | - |
| Hendry3 | 473 | 0.828 ± 0.058 | 0.965 (0.443–1.795) | 6.94 |
| Hendry2 | 384 | 0.705 ± 0.063 | 1.078 (0.388–2.288) | 7.76 |
| Manatee2 | 552 | 0.502 ± 0.031 | 1.228 (0.487–2.767) | 8.84 |
| Hillsborough3 | 450 | 0.522 ± 0.041 | 1.372 (0.420–3.418) | 9.87 |
| Manatee4 | 272 | 0.457 ± 0.047 | 1.492 (0.308–5.174) | 10.73 |
| Manatee3 | 581 | 0.620 ± 0.033 | 1.978 (0.916–4.011) | 14.23 |
| Hillsborough4 | 235 | 0.460 ± 0.045 | 2.027 (0.437–7.402) | 14.58 |
| IndianRiver-Emilia | 278 | 0.570 ± 0.045 | 2.760 (0.797–8.254) | 19.86 |
| Manatee1 | 454 | 0.481 ± 0.040 | 3.057 (1.063–7.227) | 21.99 |
| IndianRiver-Squash | 861 | 0.566 ± 0.026 | 3.621 (1.903–6.675) | 26.05 |
| IndianRiver1 | 382 | 0.707 ± 0.057 | 4.239 (1.947–8.458) | 30.50 |
| Hillsborough1 | 373 | 1.216 ± 0.072 | 4.477 (2.687–7.225) | 32.21 |
| Hillsborough5 | 227 | 0.477 ± 0.044 | 5.089 (1.592–15.549) | 36.61 |
| Hillsborough-Emilia | 538 | 0.641 ± 0.032 | 5.636 (3.026–10.324) | 40.55 |
| Miami-Dade3 | 240 | 0.505 ± 0.045 | 10.076 (2.753–37.392) | 72.49 |
| Hillsborough2 | 470 | 0.821 ± 0.052 | 10.879 (5.734-21.010) | 78.27 |
| Miami-Dade1 | 577 | 0.812 ± 0.047 | 12.676 (7.853–20.769) | 91.19 |
| Hendry1 | 560 | 0.749 ± 0.043 | 16.484 (10.156–27.410) | 118.59 |
| Miami-Dade2 | 400 | 0.530 ± 0.052 | 24.430 (11.541–59.064) | 175.76 |
| Population Code 1 | Mortality | Resistance Ratio 4 | ||
|---|---|---|---|---|
| No. Tested 2 | Slope ± SE | LC50 (95% CL) 3 | ||
| Laboratory | 499 | 0.564 ± 0.054 | 0.026 (0.002–0.122) | - |
| Manatee4 | 257 | 0.457 ± 0.047 | 0.043 (0.003–0.240) | 1.65 |
| IndianRiver-Emilia | 298 | 0.570 ± 0.045 | 0.068 (0.009–0.289) | 2.62 |
| Hendry2 | 375 | 0.705 ± 0.063 | 0.125 (0.024–0.377) | 4.81 |
| Manatee2 | 515 | 0.502 ± 0.031 | 0.187 (0.056–0.497) | 7.19 |
| Manatee3 | 542 | 0.620 ± 0.033 | 0.189 (0.065–0.462) | 7.27 |
| Hendry3 | 467 | 0.828 ± 0.058 | 0.217 (0.075–0.494) | 8.35 |
| Hillsborough3 | 451 | 0.522 ± 0.041 | 0.320 (0.069–0.971) | 12.31 |
| IndianRiver1 | 363 | 0.707 ± 0.057 | 0.356 (0.099–0.891) | 13.69 |
| Hillsborough-Emilia | 555 | 0.641 ± 0.032 | 0.396 (0.178–0.800) | 15.23 |
| IndianRiver-Squash | 774 | 0.566 ± 0.026 | 0.494 (0.215–1.019) | 19.00 |
| Miami-Dade3 | 234 | 0.505 ± 0.045 | 0.556 (0.095–2.131) | 21.39 |
| Hillsborough2 | 431 | 0.821 ± 0.052 | 0.575 (0.200–1.298) | 22.12 |
| Hillsborough4 | 213 | 0.460 ± 0.045 | 0.587 (0.090–2.418) | 22.58 |
| Manatee1 | 462 | 0.481 ± 0.040 | 0.589 (0.149–1.621) | 22.65 |
| Miami-Dade2 | 361 | 0.530 ± 0.052 | 0.930 (0.284–2.228) | 35.77 |
| Hillsborough5 | 213 | 0.477 ± 0.044 | 0.953 (0.237–3.025) | 36.65 |
| Hillsborough1 | 330 | 1.216 ± 0.072 | 1.233 (0.624–2.196) | 47.42 |
| Miami-Dade1 | 539 | 0.812 ± 0.047 | 1.244 (0.642–2.175) | 47.85 |
| Hendry1 | 562 | 0.749 ± 0.043 | 3.350 (1.960–5.451) | 128.85 |
| Population Code 1 | Mortality | Resistance Ratio 4 | ||
|---|---|---|---|---|
| No. Tested 2 | Slope ± SE | LC50 (95% CL) 3 | ||
| Laboratory | 491 | 0.564 ± 0.054 | 0.010 (0.001–0.060) | - |
| Manatee4 | 259 | 0.457 ± 0.047 | 0.011 (<0.001–0.085) | 1.1 |
| Hendry3 | 477 | 0.828 ± 0.058 | 0.117 (0.036–0.290) | 11.7 |
| Manatee3 | 574 | 0.620 ± 0.033 | 0.127 (0.041–0.323) | 12.7 |
| Hendry2 | 380 | 0.705 ± 0.063 | 0.140 (0.026–0.420) | 14.0 |
| IndianRiver-Emilia | 287 | 0.570 ± 0.045 | 0.151 (0.026–0.566) | 15.1 |
| Manatee1 | 476 | 0.481 ± 0.040 | 0.177 (0.033–0.575) | 17.7 |
| IndianRiver-Squash | 814 | 0.566 ± 0.026 | 0.189 (0.074–0.419) | 18.9 |
| Miami-Dade3 | 252 | 0.505 ± 0.045 | 0.193 (0.025–0.825) | 19.3 |
| Hillsborough3 | 439 | 0.522 ± 0.041 | 0.194 (0.037–0.633) | 19.4 |
| Manatee2 | 538 | 0.502 ± 0.031 | 0.269 (0.087–0.675) | 26.9 |
| Hillsborough4 | 251 | 0.460 ± 0.045 | 0.274 (0.041–1.094) | 27.4 |
| IndianRiver1 | 378 | 0.707 ± 0.057 | 0.325 (0.086–0.835) | 32.5 |
| Hillsborough2 | 541 | 0.821 ± 0.052 | 0.364 (0.121–0.823) | 36.4 |
| Hillsborough-Emilia | 579 | 0.641 ± 0.032 | 0.379 (0.171–0.760) | 37.9 |
| Hillsborough1 | 337 | 1.216 ± 0.072 | 0.548 (0.250–1.041) | 54.8 |
| Miami-Dade1 | 558 | 0.812 ± 0.047 | 0.613 (0.289–1.136) | 61.3 |
| Hillsborough5 | 236 | 0.477 ± 0.044 | 0.646 (0.152–2.054) | 64.6 |
| Miami-Dade2 | 360 | 0.530 ± 0.052 | 0.790 (0.232–1.937) | 79.0 |
| Hendry1 | 585 | 0.749 ± 0.043 | 1.471 (0.802–2.477) | 147.1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, H.A.; Nagle, C.A.; MacVean, C.A.; McKenzie, C.L. Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to Imidacloprid, Thiamethoxam, Dinotefuran and Flupyradifurone in South Florida. Insects 2016, 7, 57. https://doi.org/10.3390/insects7040057
Smith HA, Nagle CA, MacVean CA, McKenzie CL. Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to Imidacloprid, Thiamethoxam, Dinotefuran and Flupyradifurone in South Florida. Insects. 2016; 7(4):57. https://doi.org/10.3390/insects7040057
Chicago/Turabian StyleSmith, Hugh A., Curtis A. Nagle, Charles A. MacVean, and Cindy L. McKenzie. 2016. "Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to Imidacloprid, Thiamethoxam, Dinotefuran and Flupyradifurone in South Florida" Insects 7, no. 4: 57. https://doi.org/10.3390/insects7040057
APA StyleSmith, H. A., Nagle, C. A., MacVean, C. A., & McKenzie, C. L. (2016). Susceptibility of Bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) to Imidacloprid, Thiamethoxam, Dinotefuran and Flupyradifurone in South Florida. Insects, 7(4), 57. https://doi.org/10.3390/insects7040057