
Functional Responses of Three Neotropical Mirid Predators to Eggs of Tuta absoluta on Tomato


Abstract
:
1. Introduction
2. Material and Methods
2.1. Insect Rearing
2.2. Determination of Functional Response
2.3. Data Analysis
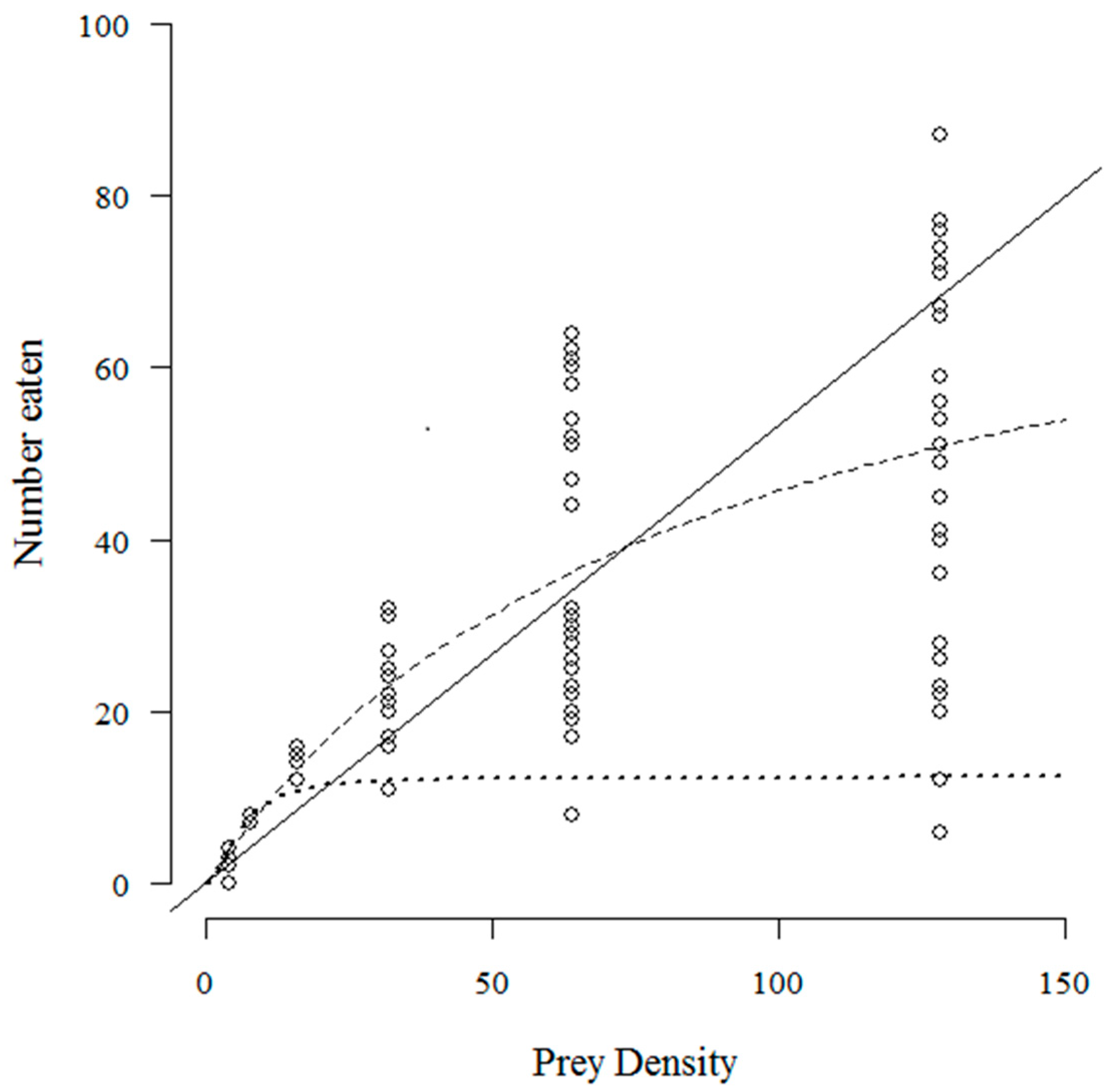
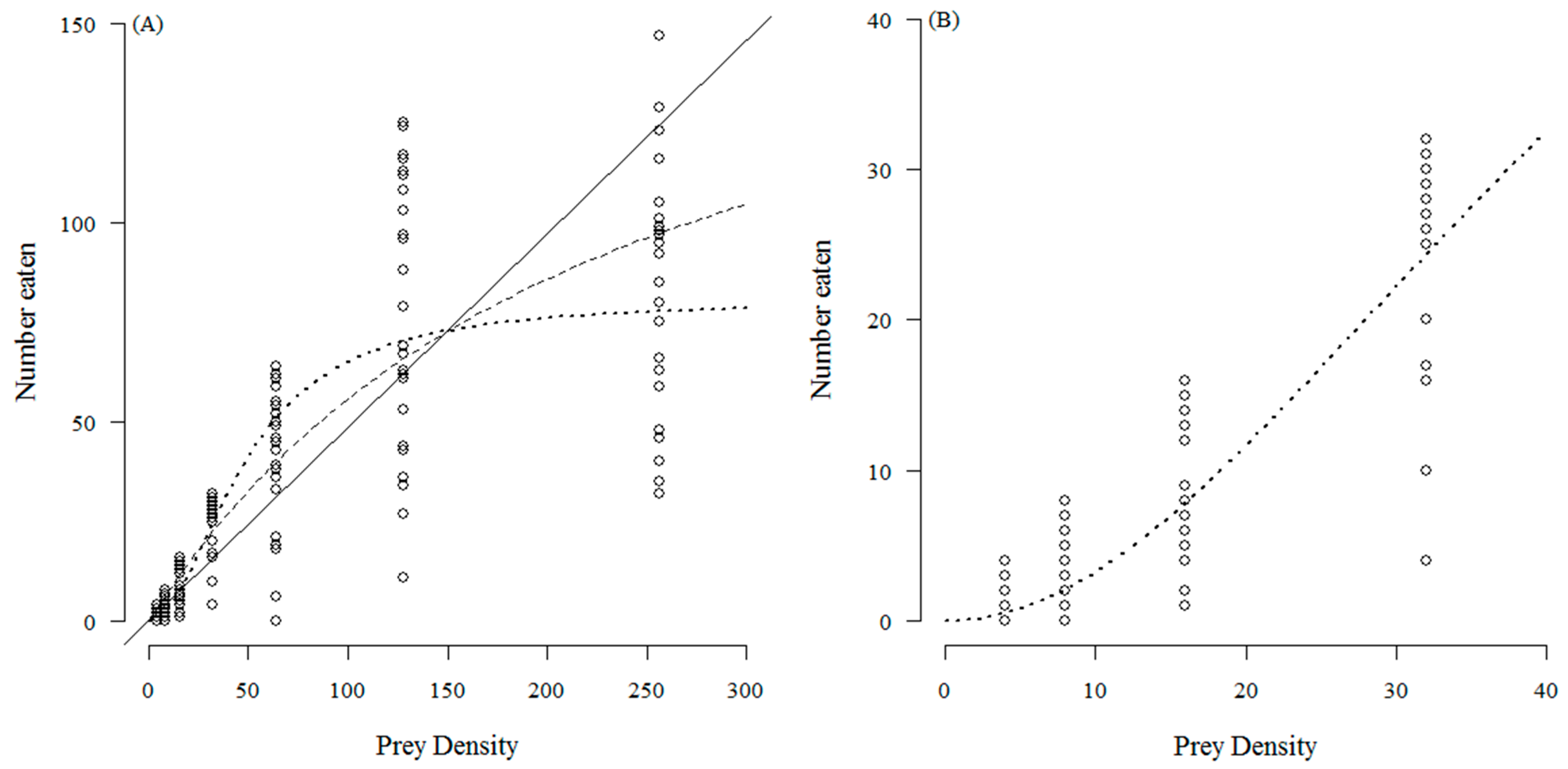
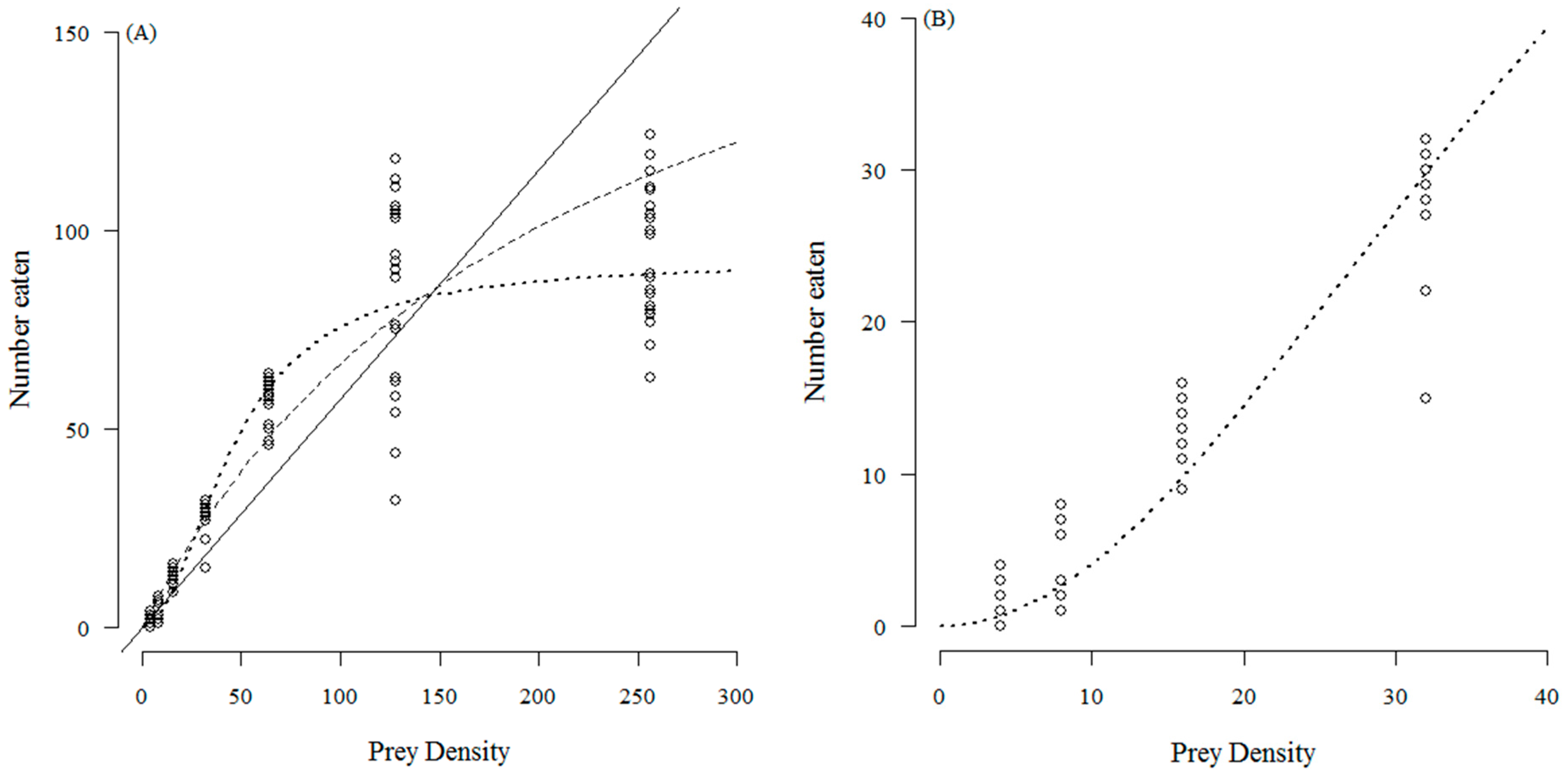
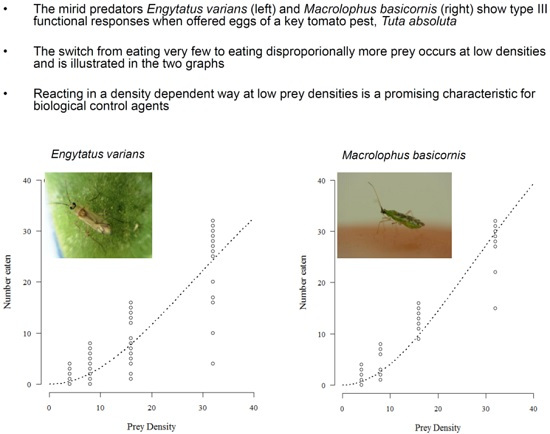
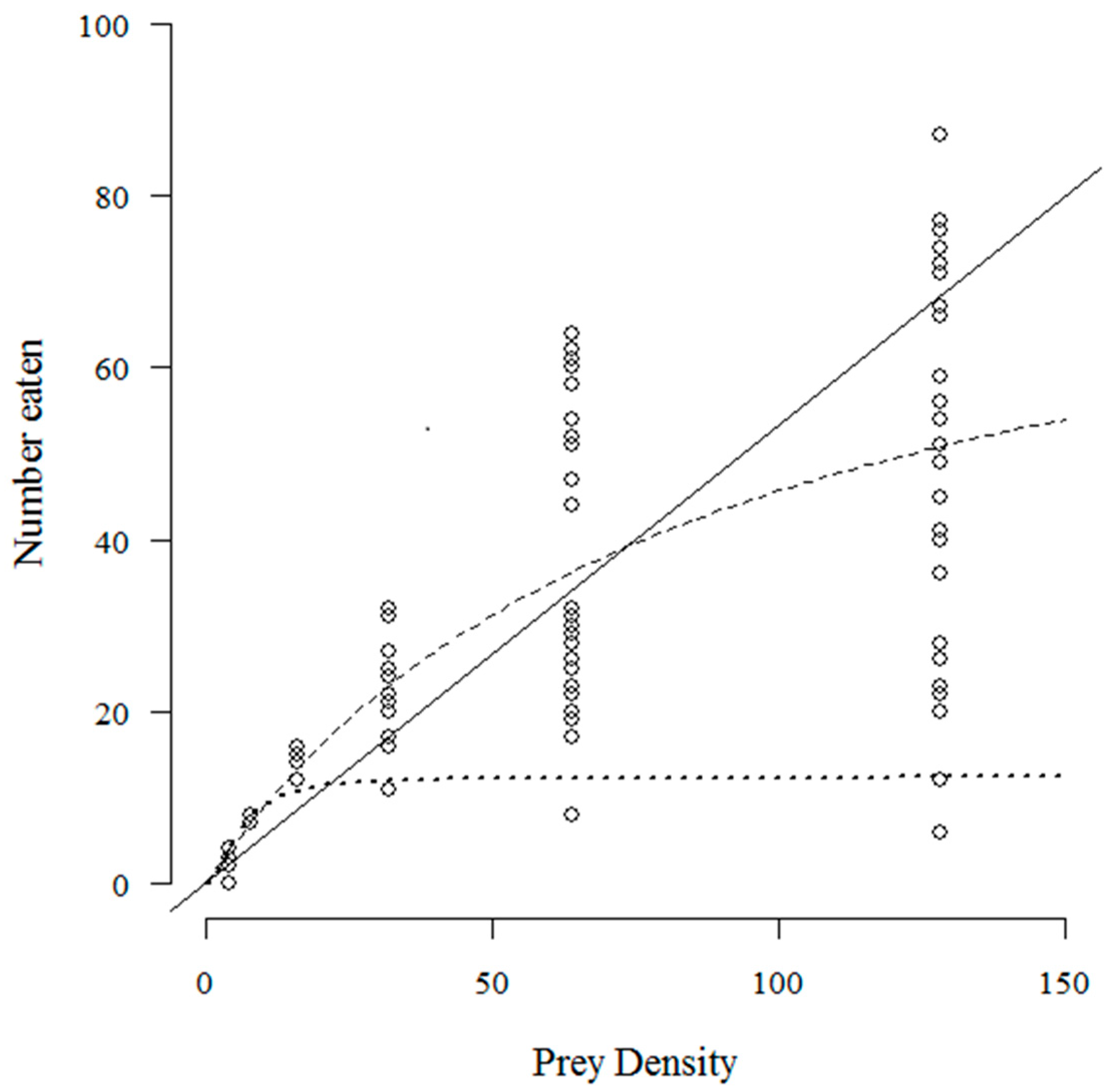
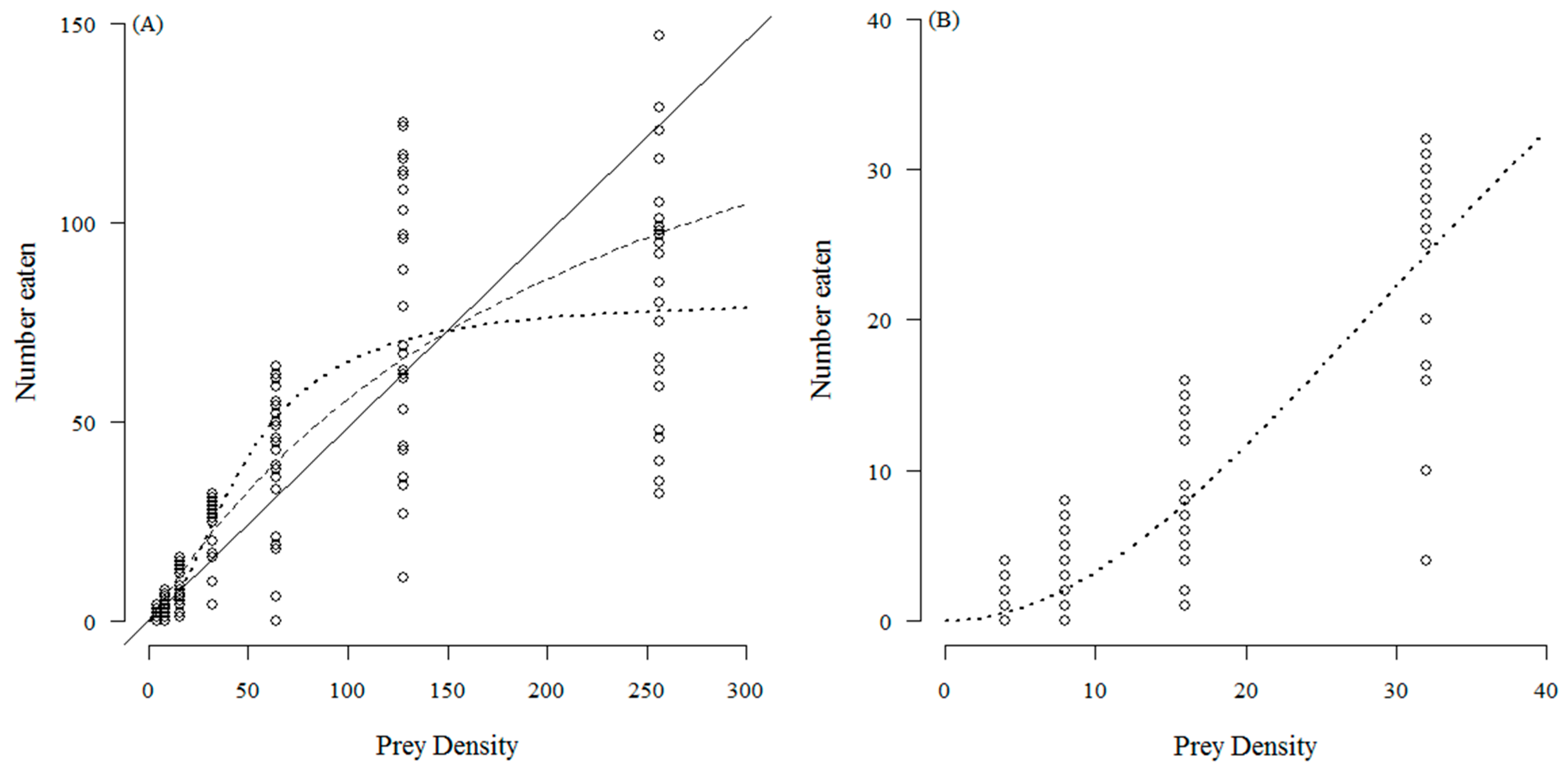
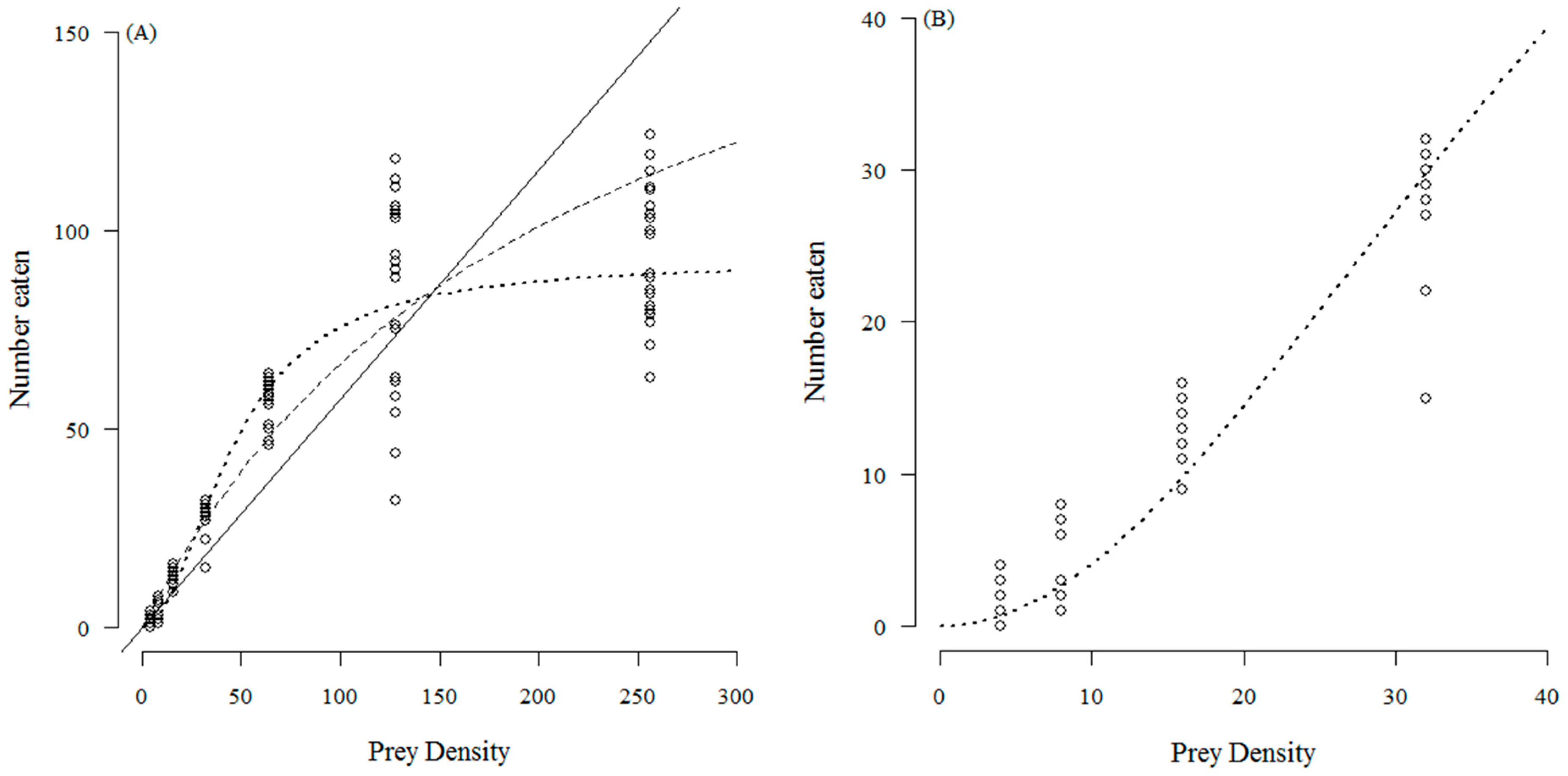
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- DeBach, P.; Rosen, D. Biological Control by Natural Enemies, 2nd ed.; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. Biocontrol 2012, 57, 71–84. [Google Scholar] [CrossRef]
- Solomon, M.E. The natural control of animal populations. J. Anim. Ecol. 1949, 19, 1–35. [Google Scholar] [CrossRef]
- Holling, C.S. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entom. Soc. Can. 1965, 45, 1–60. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Murdoch, W.W. The functional response of predators. J. Appl. Ecol. 1973, 10, 335–342. [Google Scholar]
- Foglar, H.; Malausa, J.; Wajnberg, E. The functional response and preference of Macrolophus caliginosus [Heteroptera: Miridae] for two of its prey: Myzus persicae and Tetranychus urticae. Entomophaga 1990, 35, 465–474. [Google Scholar] [CrossRef]
- Enkegaard, A.; Brødsgaard, H.F.; Hansen, D.L. Macrolophus caliginosus: Functional response to whiteflies and preference and switching capacity between whiteflies and spider mites. Entomol. Exp. Appl. 2001, 101, 81–88. [Google Scholar] [CrossRef]
- PQR-EPPO Data Base on Quarantine Pests. Available online: http://www.eppo.int (accessed on 8 April 2016).
- Zappala, L.; Alma, A.; Al-Jboory, I.J.; Arnó, J.; Bayram, A.; Chailleux, A.; El-Armaouty, A.; Gerling, D.; Guenaoui, Y.; Shaltiel-Harpaz, L.; et al. Natural enemies of the South American moth, Tuta absoluta, in Europe, North Africa and Middle East, and their potential use in pest control strategies. J. Pest. Sci. 2013, 86, 635–647. [Google Scholar] [CrossRef]
- Luna, M.G.; Pereyra, P.C.; Coviella, C.E.; Nieves, E.; Savino, V.; Gervassio, N.G.S.; Luft, E.; Virla, E.; Sanchez, N. Potential of biological control agents against Tuta absoluta (Lepidoptera: Gelechiidae): Current knowledge in Argentina. Fla. Entomol. 2015, 98, 489–494. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Release rate for a pre-plant application of Nesidiocoris tenuis for Bemisia tabaci control in tomato. BioControl 2012, 57, 809–817. [Google Scholar] [CrossRef]
- Urbaneja, A.; González-Cabrera, J.; Arnó, J.; Gabarra, R. Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest. Manag. Sci. 2012, 68, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Nagoya Protocol. Available online: https://www.cbd.int/abs/ (accessed on 8 April 2016).
- Bueno, V.H.P.; Montes, F.C.; Calixto, A.M.; Lins, J.C., Jr.; van Lenteren, J.C. Can recently found Brazilian hemipteran predatory bugs control Tuta absoluta? IOBC-WPRS Bull. 2012, 80, 63–67. [Google Scholar]
- Bueno, V.H.P.; van Lenteren, J.C.; Lins, J.C., Jr.; Calixto, A.M.; Silva, D.B.; Santiago, L.D.; Pérez, L.M. New records of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) predation by Brazilian Hemipteran predatory bugs. J. Appl. Entomol. 2013, 137, 29–34. [Google Scholar] [CrossRef]
- Silva, D.B.; Bueno, V.H.P.; Montes, F.C.; van Lenteren, J.C. Population growth of three mirid predatory bugs feeding on eggs and larvae of Tuta absoluta on tomato. BioControl 2016, in press. [Google Scholar] [CrossRef]
- Silva, D.B.; Bueno, V.H.P.; Lins, J.C., Jr.; van Lenteren, J.C. Life history data and population growth of Tuta absoluta at constant and alternating temperatures on two tomato lines. Bull. Insectol. 2015, 68, 223–232. [Google Scholar]
- Bueno, V.H.P.; Smit, J.; van Lenteren, J.C.; Laboratory Chemical Ecology and Insect Behavior, Department of Entomology and Acarology, Escola Superior de Agricultura Luiz de Queiroz (ESALQ/USP), Piracicaba, Brazil. Unpublished data. 2016.
- Bolker, B.M. Ecological Models and Data in R; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Hassell, M.; Lawton, J.; Beddington, J. Sigmoid functional responses by invertebrate predators and parasitoids. J. Anim. Ecol. 1977, 46, 249–262. [Google Scholar] [CrossRef]
- Streams, F. Effect of prey size on attack components of the functional response by Notonecta undulata. Oecologia 1994, 98, 57–63. [Google Scholar] [CrossRef]
- Hemerik, L.; Yano, E. Scaling up from individual behaviour of Orius sauteri foraging on Thrips palmi to its daily functional response. Popul. Ecol. 2011, 53, 563–572. [Google Scholar] [CrossRef]
- De Clercq, P.; Mohaghegh, J.; Tirry, L. Effect of host plant on the functional response of the predator Podisus nigrispinus (Heteroptera Pentatomidae). Biol. Control 2000, 18, 65–70. [Google Scholar] [CrossRef]
- Mohaghegh, J.; De Clercq, P.; Tirry, L. Functional response of the predators Podisus maculiventris (Say) and Podisus nigrispinus (Dallas) (Het., Pentatomidae) to the beet armyworm, Spodoptera exigua (Hübner) (Lep., Noctuidae): Effect of temperature. J. Appl. Entomol. 2001, 125, 131–134. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bakker, K. Functional responses in invertebrates. Neth. J. Zool. 1976, 26, 567–572. [Google Scholar] [CrossRef]
- Trexler, J.C.; McCulloch, C.E.; Travis, J. How can the functional response best be determined? Oecologia 1988, 76, 206–214. [Google Scholar] [CrossRef]
- Wheeler, A.G. Biology of the Plant. Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists; Cornell University Press: Ithaca, NY, USA, 2001. [Google Scholar]
- Bueno, V.H.P.; van Lenteren, J.C. Predatory Bugs (Heteroptera). In Insect Bioecology and Nutrition for Integrated Pest Management; Panizzi, A.R., Parra, J.R.P., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 539–569. [Google Scholar]
- Silva, D.B.; Bueno, V.H.P.; Calvo, F.J.; van Lenteren, J.C. Do nymphs and adults of three Neotropical zoophytophagous mirids damage leaves and fruits of tomato? Bull. Entomol. Res. 2016. accepted. [Google Scholar]
- Montserrat, M.; Albajes, R.; Castañé, C. Functional response of four heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower thrips (Thysanoptera: Thripidae). Environ. Entomol. 2000, 29, 1075–1082. [Google Scholar] [CrossRef]
- Alvarado, P.; Balta, O.; Alomar, O. Efficiency of four Heteroptera as predators of Aphis gossypii and Macrosiphum euphorbiae (Hom.: Aphididae). Entomophaga 1997, 42, 215–226. [Google Scholar] [CrossRef]
- Fantinou, A.; Perdikis, D.C.; Labropoulos, P.; Maselou, D. Preference and consumption of Macrolophus pygmaeus preying on mixed instar assemblages of Myzus persicae. Biol. Control 2009, 51, 76–80. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bakker, K. Behavioural aspects of the functional response of a parasite (Pseudeucoila bochei Weld) to its host (Drosophila melanogaster). Neth. J. Zool. 1978, 28, 213–233. [Google Scholar] [CrossRef]
- Luck, R.F.; van Lenteren, J.C.; Twine, P.H.; Kuenen, L.; Unruh, T. Prey or host searching behavior that leads to a sigmoid functional response in invertebrate predators and parasitoids. Popul. Ecol. 1979, 20, 257–264. [Google Scholar] [CrossRef]
- Van Roermund, H.J.W.; Hemerik, L.; van Lenteren, J.C. Influence of intra-patch experiences and temperature on the time allocation of the whitefly parasitoid Encarsia formosa (Hymenoptera: Aphelinidae). J. Insect Behav. 1994, 7, 483–501. [Google Scholar] [CrossRef]
- Faria, C.A.; Torres, J.B.; Fernandes, A.M.V.; Farias, A.M.I. Parasitism of Tuta absoluta in tomato plants by Trichogramma pretiosum Riley in response to host density and plant structures. Ciênc. Rural 2008, 38, 1504–1509. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bueno, V.H.P.; Smit, J.; Soares, M.A.; Calixto, A.M.; Lins, J.C.; Laboratory of Entomology, Wageningen University, Wageningen, The Netherlands. Unpublished data. 2016.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mirid Species | AICc | ΔAICc | AICc Weight | a | h |
|---|---|---|---|---|---|
| (a) C. infumatus | |||||
| Holling type I | 2715.1 | 903.2 | ≈0 | ||
| Holling type II | 1811.9 | 0 | ≈1 | 84.87 | 85.83 |
| Holling type III | 5292.2 | 3480.3 | ≈0 | ||
| (b) E. varians | |||||
| Holling type I | 4675.6 | 1083.2 | ≈0 | ||
| Holling type II | 3631.6 | 39.2 | ≈0 | ||
| Holling type III | 3592.4 | 0 | ≈1 | 80.53 | 48.54 |
| (c) M. basicornis | |||||
| Holling type I | 4351.0 | 2288.2 | ≈0 | ||
| Holling type II | 2163.6 | 100.8 | ≈0 | ||
| Holling type III | 2062.8 | 0 | ≈1 | 91.81 | 46.16 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Lenteren, J.C.; Hemerik, L.; Lins, J.C.; Bueno, V.H.P. Functional Responses of Three Neotropical Mirid Predators to Eggs of Tuta absoluta on Tomato. Insects 2016, 7, 34. https://doi.org/10.3390/insects7030034
Van Lenteren JC, Hemerik L, Lins JC, Bueno VHP. Functional Responses of Three Neotropical Mirid Predators to Eggs of Tuta absoluta on Tomato. Insects. 2016; 7(3):34. https://doi.org/10.3390/insects7030034
Chicago/Turabian StyleVan Lenteren, Joop C., Lia Hemerik, Juracy C. Lins, and Vanda H. P. Bueno. 2016. "Functional Responses of Three Neotropical Mirid Predators to Eggs of Tuta absoluta on Tomato" Insects 7, no. 3: 34. https://doi.org/10.3390/insects7030034
APA StyleVan Lenteren, J. C., Hemerik, L., Lins, J. C., & Bueno, V. H. P. (2016). Functional Responses of Three Neotropical Mirid Predators to Eggs of Tuta absoluta on Tomato. Insects, 7(3), 34. https://doi.org/10.3390/insects7030034