1. Introduction
Worldwide there has been an increased interest in alternative pest management strategies to reduce pesticide use and create more environmentally sound production systems. One such alternative is an ecological management approach called a “push-pull strategy” [
1,
2]. One form of this approach employs the use of a trap crop (pull) and a deterrent intercrop (push) to effectively prevent a pest from settling on the main crop [
1]. This strategy has been used in some systems to target Lepidoptera pests and reduce larval feeding damage on the main crop. An example of this cropping approach is the effective management of the stemborer,
Chilo partellus Swinhoe (Lepidoptera: Crambidae), on maize (
Zea mays L.) by the use of Napier grass (
Pennisetum purpureum Schumach) as a trap crop and molasses grass (
Melinis minutiflora P. Beauv.) as an intercrop [
1,
3]. In this system the adult females prefer to lay their eggs on Napier grass and are repelled by molasses grass leading to lower oviposition on maize. A similar push-pull approach was evaluated for
Eldana saccharina Walker (Lepidoptera: Pyralidae) management in sugarcane where the determination was that adult female moths were not strongly attracted or repelled by the plants tested; these plants were not recommended for use in a push-pull management approach [
4]. Thus, in the case of lepidopteran pests, the key to the success of this strategy lies in the oviposition preferences of the adult females. The trap crop should be preferred for oviposition over the main crop and the intercrop should be an oviposition deterrent which, upon contact, will stimulate movement elsewhere for oviposition [
5]. However, larval mobility has important implications for pest management strategies that involve polyculture, such as trap cropping or a push-pull system. Ideally, larvae remain and feed on the trap crop and the main crop receives little or no damage. If the larvae, as well as the adults, prefer the trap crop to the main crop, then this could enhance the efficacy of the push-pull system by increasing tenure times and decreasing larval movement within the trap crop [
6]. However, larvae are capable and often stimulated to move between plants [
7,
8]. So, even if adult females lay their eggs on the trap crop, subsequent movement of larvae into the adjacent main crop could minimize the effectiveness of a push-pull strategy. Although oviposition preference is crucial for the success of this pest management strategy, the behavior of larvae also has to be considered, especially if this is the stage when damage is inflicted, as is the case with many lepidopterans [
9].
Pickleworms, the larvae of
Diaphania nitidalis Cramer (Lepidoptera: Crambidae), are a pest of cucurbits found in Hawaii, North America (primarily in the Southeastern United States), Central and South America, and the West Indies [
10,
11,
12,
13,
14]. Pickleworms feed on the fruits, buds, flowers, and stems of cucurbits [
10,
11,
13]. Newly-hatched larvae feed on vegetative tissues, buds, and flowers, while older larvae tend to feed on fruits or tunnel the stems if the former are unavailable [
12,
13]. Pickleworms are primarily managed through weekly applications of insecticides and such practice is generally considered unsustainable. A potentially sustainable and environmentally sound management strategy for this pest could include push-pull cropping. We propose a potential push-pull system for pickleworm management on cantaloupe using squash (
Cucurbita pepo L.) as a trap crop and watermelon (
Citrullus lanatus [Thunb.] Matsum. and Nakai) as an intercrop. Squash was selected as a trap crop because it has been suggested that the use of squash as a trap crop (“pull”) might lower the risk of damage by pickleworm to other cucurbit crops [
10,
11,
15,
16]. Adult females have been shown to preferentially lay their eggs on squash plants [
17,
18] but field trials using squash as a trap crop have had variable success in reducing pickleworm damage to other cucurbit main crops [
10,
11,
12,
16]. It may be that even though there is a preference for oviposition on squash over cantaloupe that larval movement leads to reduced success of this strategy. In our study, watermelon was selected as an intercrop (“push”) primarily because it has typically had very low pickleworm damage despite being a cucurbit [
18,
19]. This could be because the adult moths lay a relatively low number of eggs on this plant. In addition, other factors, such as larval movement off the plant, could also contribute to the low level of damage seen.
In this study, we investigated the behavior of neonate pickleworm larvae, in relation to the potential push-pull cropping approach. Understanding the behavior of neonates is important to evaluate the potential of the proposed push-pull system. There may be various stimuli for neonate larval movement including crowding, reduced food resources or quality, presence of predators or parasitoids, microclimate preferences and feeding preferences [
7]. If neonate larvae present on a squash plant (pull/trap crop) are stimulated to move and in their travel encounter either a cantaloupe or a watermelon plant. Their acceptance of either of these as a food plant will influence whether they remain on that plant or whether they continue to search for another host plant. The same would be true for a larva moving from a watermelon or a cantaloupe. Thus, understanding such larval preference could influence the effectiveness of a push-pull approach for management where the aim is to create a more complex habitat to influence pest behavior [
4,
9].
Furthermore, there may be an influence of early feeding on subsequent host plant choices should the larvae migrate away from its natal host plant. Lepidopteran larvae may acquire an induced preference after feeding on a particular host plant [
20]. Experience has been shown to influence subsequent feeding activity. For example, after having eaten cabbage through its fourth instar,
Pieris brassicae L. (Lepidoptera: Pieridae) larvae choose to starve to death rather than to feed on another host plant, nasturtium (
Tropaeolum majus L.) [
20]. The same phenomenon was found true for
Pieris rapae L. (Lepidoptera: Pieridae) [
21]. Thus, if an adult female pickleworm lays eggs on her preferred host plant and the larvae feed on that plant, it could become preferred over other plants and influence subsequent choices if the larvae left their natal host plant. Such a preference based on experience would be beneficial if the eggs were originally laid on the trap crop and the neonates then would choose squash over cantaloupe. However, this experience-induced preference could be damaging if it keeps any larvae within the main crop instead of traveling to the trap crop if they are stimulated to move.
There are other factors that will influence the success of neonate interplant movement and, hence, their interaction with the habitat. First, in spite of their small size, neonate larvae must be capable of moving relatively great distances. The mobility of neonate lepidopteran larvae has been previously described. For example, young larvae of the diamondback moth (
Plutella xylostella L. (Lepidoptera: Plutellidae)) are capable of movement off their natal plants to adjacent ones [
8]. Such behavior has also been described in the neonate larvae of the light brown apple moth (
Epiphyas postvittana Walker (Lepidoptera: Tortricidae)) [
22]. Foster and Howard concluded that these neonates are very mobile and their preferences have more influence than the adult female oviposition preference on the selection of the plants on which the larvae subsequently feed [
22]. Although neonate pickleworms are small, around 1.5 mm in length, we observed in the lab that they are highly mobile; moving across lab counters and up walls in search of food after hatching from a cage in which no food was available to them. We observed that newly hatched neonate larvae could easily transverse the distance of 30 cm in under an hour, while walking on a flat piece of poster board. Another factor that will influence success of neonate mobility is their longevity without food. If larvae are without a food source they will eventually starve if they do not find a suitable food source and this is likely to be of critical importance for neonates where there are limited energy reserves. In the case of pickleworm, the longevity of starved neonates is unknown.
The first objective of this study was to gain a better understanding of the biology of neonate larvae; particularly the longevity of newly-hatched neonates without a food source to gain a better sense for how long a neonate could live while searching for a host plant or switching host plants instead of feeding. We conducted an experiment to examine the longevity of neonates when completely starved and after having limited access to food (24 h).
The second objective was to investigate if naive neonate larvae have a preference between host plants and if this preference can be modified through experience. To address the second objective a series of experiments were conducted to (1) see if naive neonate larvae had an orientation or settlement preference between squash, watermelon, cantaloupe, bean (non-host), and an artificial diet; and (2) to determine if experience feeding on one of the above food sources would influence the neonates’ subsequent orientation and settlement preference between the five food sources.
2. Materials and Methods
2.1. Insect Colony
Experimental larvae were obtained from a laboratory colony of pickleworm maintained at the University of Hawaii at Manoa, HI, USA. This colony was initiated in spring 2011 and sustained for two years. Larvae collected on the island of Oahu HI, USA from kabocha, zucchini, cucumber, and cantaloupe plants, were used to start and maintain the colony. The colony was keep at room temperature (22 °C) and exposed to natural day length.
Pickleworm larvae in the colony were reared on a pinto bean-based artificial diet (Bioserv Black Cutworm Diet, Newark, DE, USA). Once the larvae had finished feeding and were ready to pupate, indicated by a color change from greenish to pink, they were removed from the diet and placed into a separate container with paper towels, which they used to create a cocoon. After a few days, the pupae were collected and placed directly into the adult cage for emergence. The adult cage comprised of a fabric mesh flight cage (70 cm × 35.5 cm × 35.5 cm; BioQuip Products, Inc., Compton, CA, USA) in which the adults were provided with a 10% honey water solution. To provide an adequate level of humidity for oviposition, the cage was periodically sprayed with water and covered with white plastic trash bags. At any given time, there were up to several hundred adults in the cage of varying age and different generations, and they were allowed to mate freely. Cotton pads were hung from the top of the cage for the females to lay their eggs on, the cotton pads with eggs were then removed from the colony and placed in a glass jar. The newly-hatched neonates were obtained and used in the following experiments. The experiments were conducted over a period of two years, thus using neonates from different cohorts.
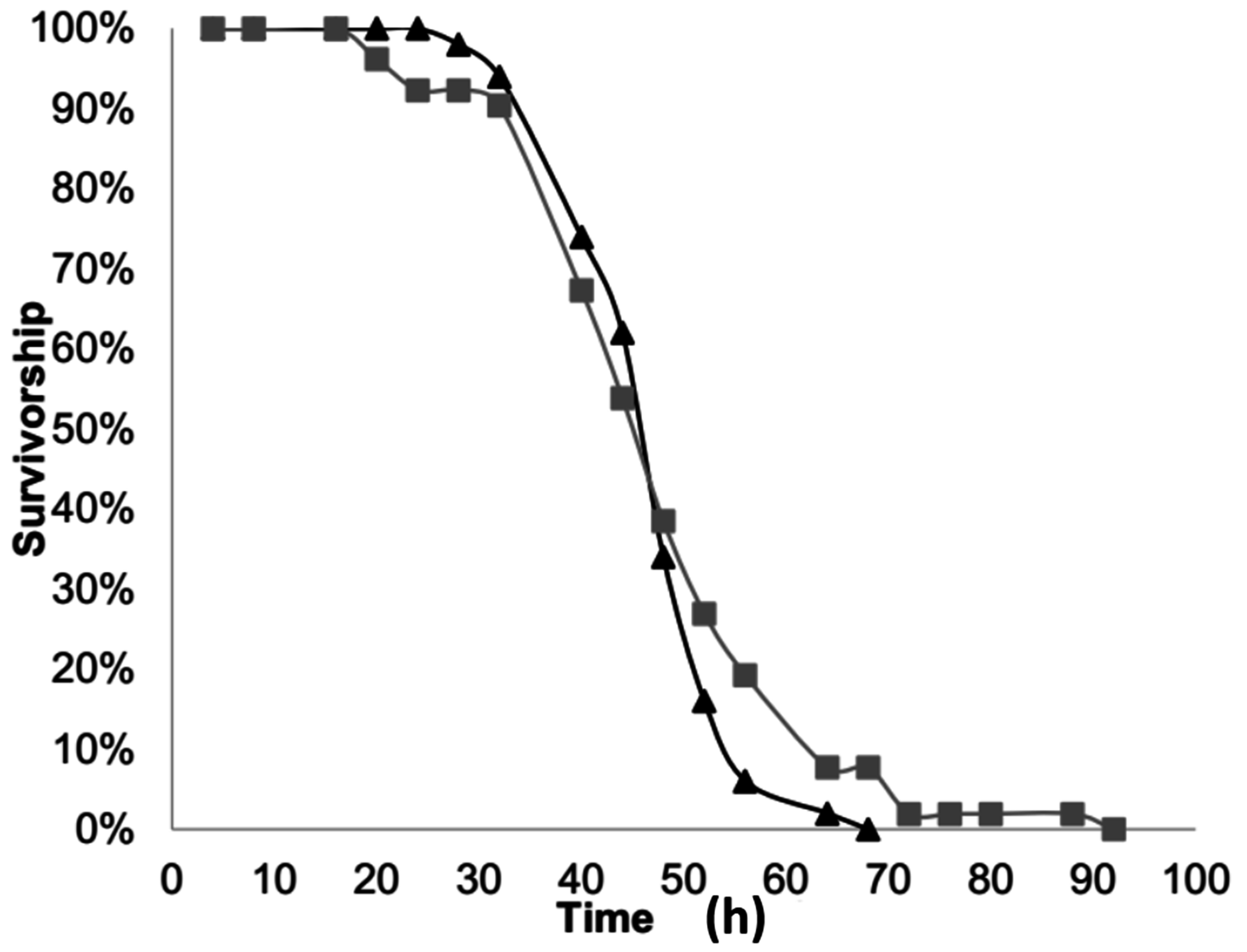
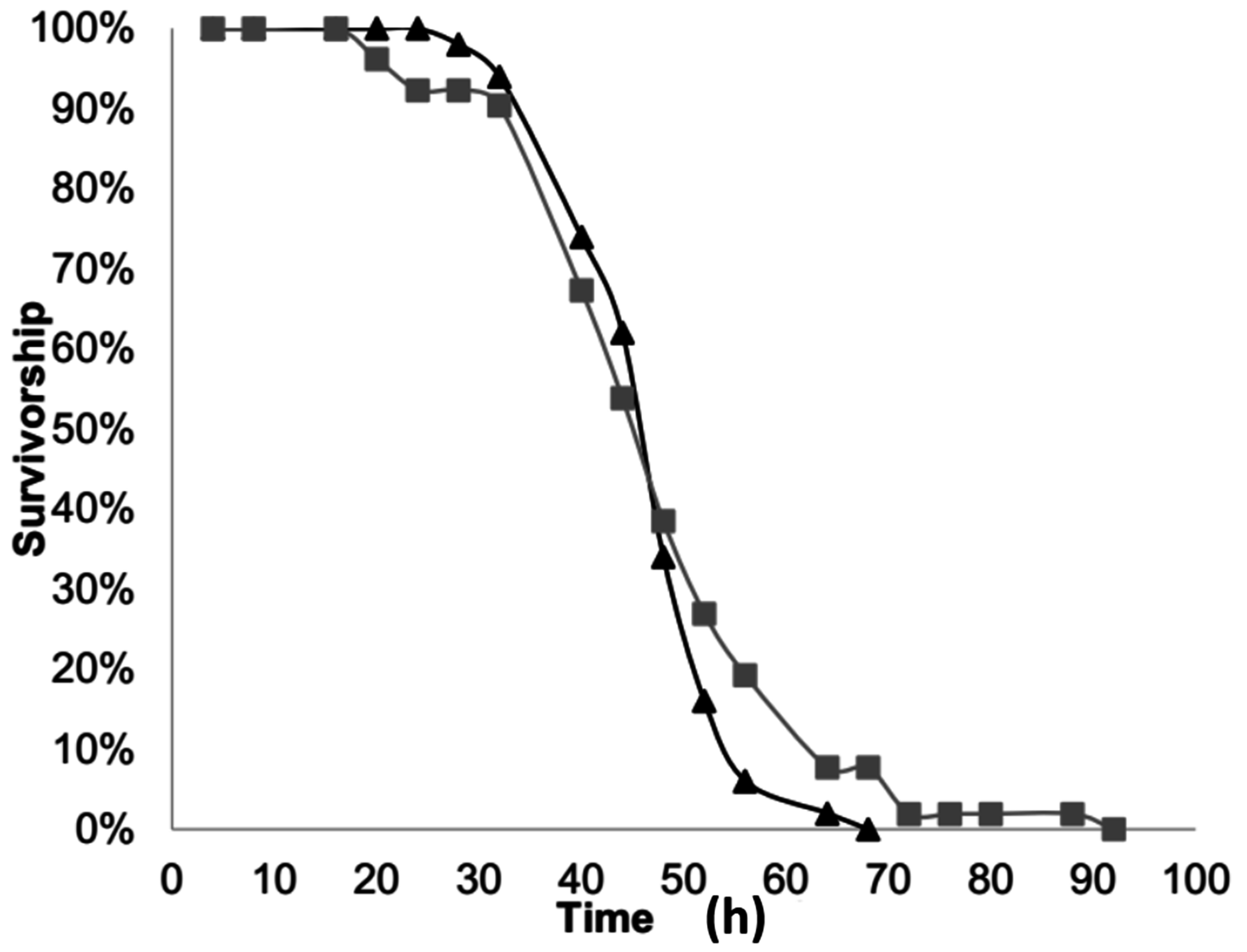
2.2. Neonate Longevity
To gather larvae for this experiment, unhatched eggs on cotton pads were in placed in plastic vials (3 cm diameter, 5.5 cm tall) with a plastic cap. After being placed in a plastic vial, the eggs within were given two hours to hatch, after which the newly hatched larvae were removed from the vial and used for the experiment. To examine neonate longevity, newly-hatched larvae were placed in one of two groups: “completely-starved” or “partially-starved”.
The neonates in the “completely-starved” group were placed in a plastic vial with no food. Larval survivorship was checked every 4 to 8 h until all larvae were dead. This was replicated fifty times. For each individual larva the total longevity was calculated as the time it was placed in the plastic vial to the last time-interval it was noted as being alive. In addition, the proportion of larvae that survived to each time interval was calculated.
The second group of “partially-starved” neonates was allowed to feed for 24 h and then placed in a plastic vial with no food, where their survivorship was checked every 4 to 8 h. To provide larvae with the opportunity to feed for 24 h each newly hatched larva was placed individually in a Petri dish (10 cm diameter × 1 cm deep) containing an l cm leaf disc of squash (Cucurbita pepo L. var. Cocozelle) atop moistened filter paper. This was replicated fifty-two times. For each larva the total longevity was calculated from the time it was placed in the Petri dish to feed to the last-time interval in the plastic vial where it was noted as being alive. Thus, for this group, the total longevity included the 24 h of feeding. The longevity post-feeding was calculated for the “partially-starved” group as the total longevity minus 24 h. In addition, the proportion of larvae that survived to each post-feeding time interval was calculated.
A two-sample t-test was used to calculate if there was a significant difference between the total longevity of the “completely-starved” and the “partially-starved” neonates. A second two-sample t-test was conducted to calculate if there was a different between the total longevity of the “completely-starved” group and the longevity post-feeding of the “partially-starved” group; thus comparing the longevity of both groups without access to food.
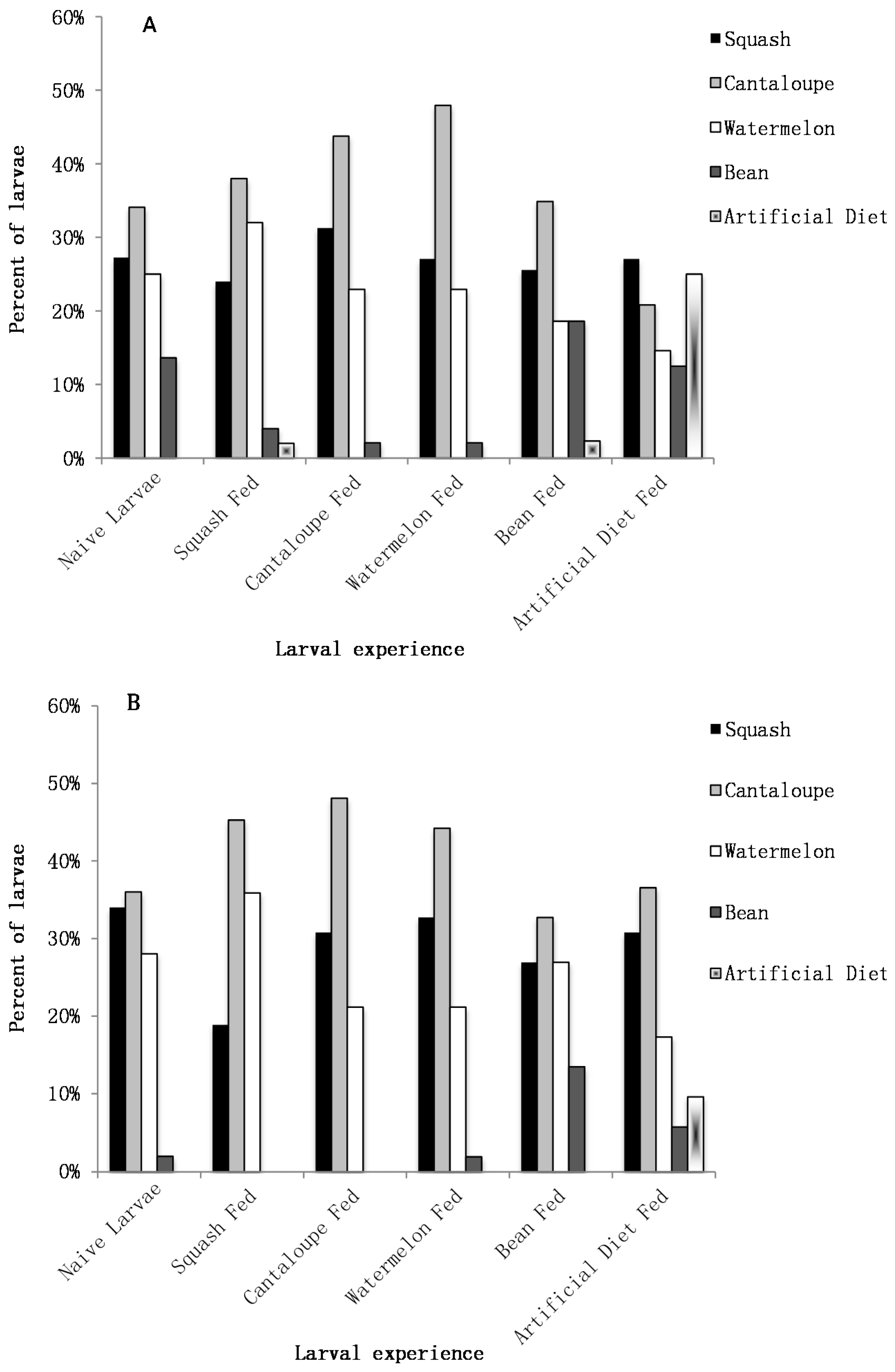
2.3. Naive Neonate Preference
Newly-hatched larvae were given a choice between different foods to investigate if they have naive orientation and settlement preferences. These preferences were determined by what food source the larva was on after 1 h (orientation preference) and after 24 h (settlement preference) of being presented with a choice of food sources. The larvae were obtained as described above. They were given a choice between potential foods: leaf discs of three host plant species in the plant family Cucurbitacae, a leaf disc from one non-host plant, and 1 cm disc of a pinto bean-based artificial diet (Bioserv Black Cutworm Diet, Newark, DE, USA). The two common host plants in Cucurbitacae were cantaloupe (
Cucumis melo L.), and squash (
Cucurbita pepo L.), and the third cucurbit, which is considered a rare host plant, was watermelon (
Citrullus lanatus [Thunb.] Matsum. and Nakai) [
11,
18]. The non-host plant used was pole bean (
Phaseolus vulgaris L.), in the plant family Fabaceae, because it has been reported that neonate larvae can feed on this plant species for 48 h without dying [
18]. The plant varieties used were Atlantia Cantaloupe, Yellow Summer Crookneck Squash, Crimson Sweet Watermelon, and Emerite Pole Bean. These were selected because they are varieties grown in Hawai’i [
23]. The artificial diet was incorporated because it had been used to maintain the pickleworm colony and had demonstrated itself as a sufficient diet for pickleworm development.
Each plant was greenhouse grown in an 8 cm square pot (9.5 cm depth) with Sunshine Mix 1 Professional Growing Mix (Pacific Agricultural Sales and Services, Inc., Kapolei, HI, USA) fertilized with 1.25 mL Osmocote Indoor and Outdoor Plant Food (Scotts-Sierra Horticultural Products Company, Marysville, OH, USA) and watered when needed, generally on a daily basis. The plants were between 27 to 31 days old when they were used in the experiments.
Leaf discs, 1 cm in diameter, were cut from leaves of each plant species. In addition, the artificial diet was prepared as a thin layer and cut in 1 cm discs. The leaf and diet discs were placed equal-distance apart from each other around the edge of a 10 cm diameter × 1 cm deep Petri dish in a random order to create a choice arena. To prevent the food discs from drying out, a moist piece of filter paper was on the bottom of each Petri dish.
To assess neonate preference one larva was placed in the center of each choice arena. The orientation preference of each larva was determined by recording the position of the larva after 1 h in the arena. The settlement preference of each larva was determined by recording the position of the larva after 24 h in the arena [
9]. If a larva was not on a food source after 1 h it was recorded as a “no-choice” and was not used in determining the orientation preference. The orientation and settlement preferences for 50 naive neonate larvae were recorded.
2.4. Experienced Neonate Preference
A second experiment was conducted to investigate if the orientation and settlement preference of the larvae is influenced by 24 h of experience feeding on a particular food source. This was performed as described above, except that once a larva hatched it was placed on one of the five food sources to feed for 24 h before it was placed in the choice arena and given a choice of all five food sources. For 24 h following hatching, each larva was placed on a 2.5 cm disc of one of the five food sources in a 5.5 cm × 3.5 cm × 3 cm plastic box and allowed to feed. The larva was then removed and placed in a choice arena with each of the five foods as described above. This was replicated at least 50 times for each of the five food sources.
For both the naive and experienced neonate preference experiments, a chi-squared test of goodness-of-fit test was used to find any unequal distributions between the larval orientation and settlement on the different food sources. If the chi-squared test indicated that there was not an equal distribution between the five food sources, then a subdivided chi-squares goodness-of-fit was done to determine if there was an equal distribution on the four plant species (the three cucurbits and bean). Finally, if there was an unequal distribution between the four plant species, then just the three cucurbit plant species were tested for an equal distribution.
4. Discussion
The longevity experiment demonstrated that neonate larvae are capable of living, on average, 42.48 h, and up to 64 h, without food. When larvae had access to food for 24 h they lived up to 112 h total, with an average of 67.1 h total. However, feeding for 24 h before starvation did not provide larvae any greater benefit once they no longer had access to food. Overall, the longevity of completely-starved or partially-starved neonate larvae indicates that they have enough energy to live for a brief period while in search of a favorable food source. In a cropping environment, it is recommended that squash rows be planted 3–4 feet apart and plants within rows be planted 24–30 inches apart [
23]. Given their 1 to 2-day longevity without feeding, neonate larvae could possess sufficient energy reserves that might allow them to move between rows of plants, if they had some sort of stimulation to do so. The question then follows: if they do move, would they be discriminating about their destination and would experience influence that decision?
Neonate pickleworm larvae are able to discriminate and make a feeding choice. They prefer plant foods to a suitable artificial diet as well as differentiate and choose host plants, cucurbits, over non-hosts, as evidenced in the movement of neonates over the course of the feeding preference experiment. Even within the first 24 h after hatching, larvae will consider their available options and make clear feeding choices based on the food sources presented to them.
Nevertheless, there does not appear to be an innate larval preference or a significant discrimination between the cucurbits even after having a prior feeding experience with a particular cucurbit species. As with the naive larvae, experienced larvae that had fed on a cucurbit for 24 h, did not orient or settle more frequently on one cucurbit over another. Overall these results indicate that there is no neonate larval preference between squash, cantaloupe, and watermelon, regardless of experience. Unless there is some trigger for movement, neonate larvae are not likely to move from their natal plant. In terms of this potential push-pull cropping system, this suggests that neonate larvae are not likely to move from the trap crop, if that is where they hatched, to the main crop due to a feeding preference alone.
A somewhat surprising result was that the influence of prior experience with a non-host food led to a change in larval behavior for both the bean-fed and artificial diet-fed neonates. Larvae that had the opportunity to feed upon bean for 24 h chose bean as frequently as the cucurbit species during both the orientation and settlement period. This pattern of orientation was the same as naive larvae; but the settlement choice was different suggesting that experience modified the behavior. It appears that experience with a non-host plant can actually influence the neonates’ preference and surprisingly make them more likely to accept a non-host plant (on which they may not be able to complete their development) in the presence of host plants.
In addition, larvae that had fed on a pinto bean-based artificial diet behaved differently than their naive counterparts, as well as those larvae that had fed upon a plant food source. The naive larvae did not orient nor settle on the pinto bean-based artificial diet. However, after having feeding experience on the artificial diet the larvae oriented to all five food sources equally. Furthermore, this was the only time larvae actually chose to settle on the artificial diet. These results indicate that the experience of a non-host food source will increase the larval acceptance of that food source even in the presence of host plants.
While interesting, this behavior of accepting of non-hosts is unlikely to influence the effectiveness of a push-pull cropping approach. Although, there has been speculation that in other polyculture systems the acceptance of non-host plants might reduce the efficiency of a polyculture approach [
8]. It was found that the diamondback moth “may lay a large percentage of their eggs on non-host plants in polycultures” and this, coupled with larval movement into the main crop, could lower the effectiveness of companion planting, intercropping, and under-sowing on that pest [
8]. However, the occurrence of this problem has more to do with the oviposition behavior of the females rather than the ability of the larvae to move from a non-host to a nearby host plant. Another study, which investigated the oviposition preference of pickleworm, suggested that female pickleworms did not prefer to lay their eggs on the non-host plant bean [
17]. In fact, the adult females seemed to avoid oviposition on bean leaves, laying more eggs on their cage rather than on the bean leaves [
17]. Whereas, when offered plants of cantaloupe, squash, and watermelon, adult female pickleworms laid eggs on all three but appear to favor squash [
17]. Therefore, even though neonate pickleworm larvae did choose to feed on bean in some instances, it is unlikely that their mothers would choose to place them on this plant to begin with.



{kind=link}
{kind=link}