
Further Screening of Entomopathogenic Fungi and Nematodes as Control Agents for Drosophila suzukii


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Insects and Control Agents
- Fungi: Metarhizium anisopliae (1% solution, supplied by Fargo Ltd, Toddington, UK); Isaria fumosorosea (1% solution) and a non-commercial coded product (Coded B, 50 mL/L) (both supplied by BASF, Littlehampton, UK).
- Nematodes: Steinernema feltiae, S. carpocapsae, S. kraussei and Heterorhabditis bacteriophora (all applied at 10,000 IJ’s/mL supplied by BASF, Littlehampton, UK).
2.2. Entomopathogenic Fungi Trials
2.2.1. Treating Infested Berries
2.2.2. Treating Uninfested Berries
2.3. Entomopathogenic Nematode Trials
Potential for Soil Drenching
2.4. Data Analysis
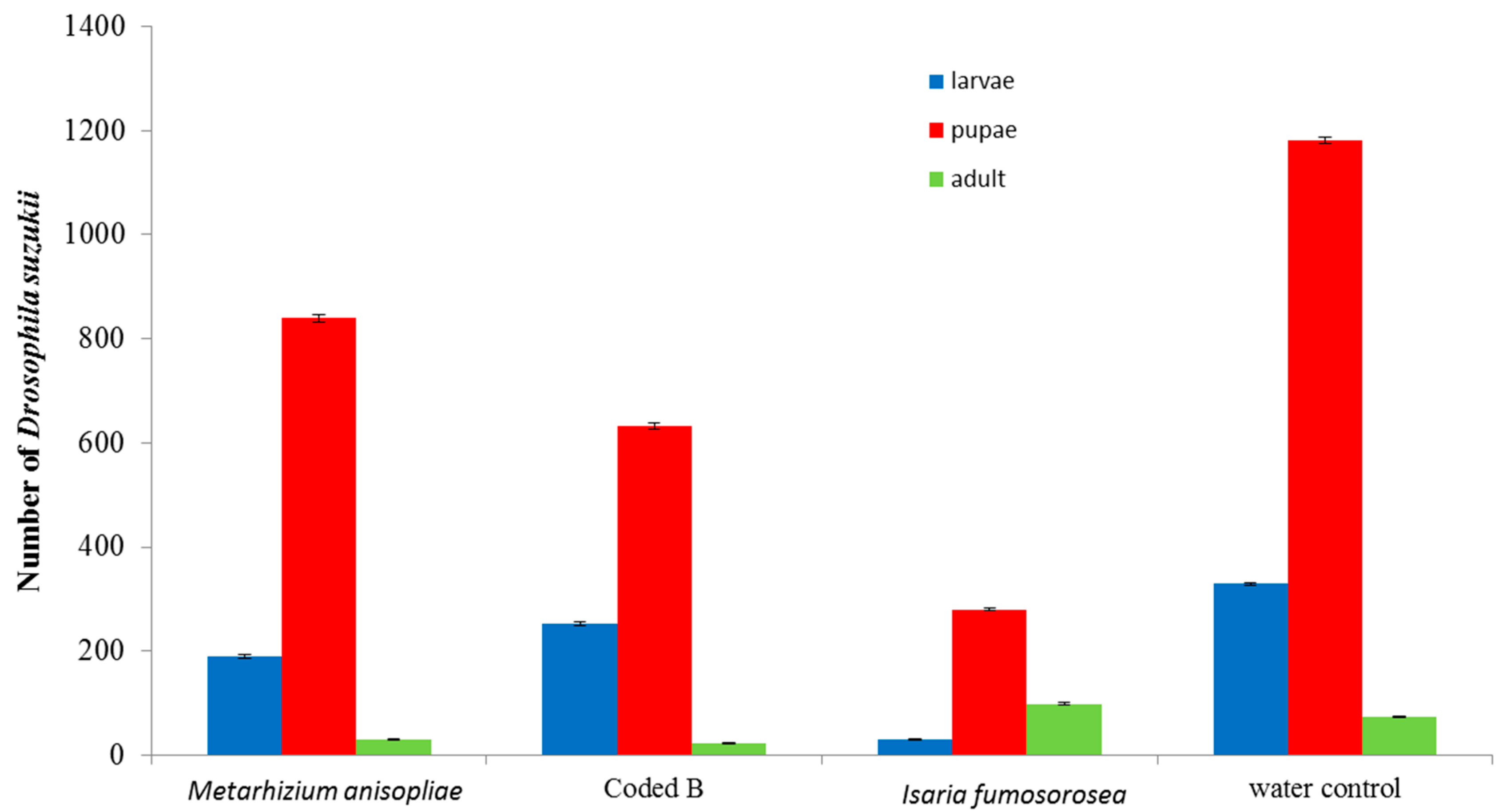
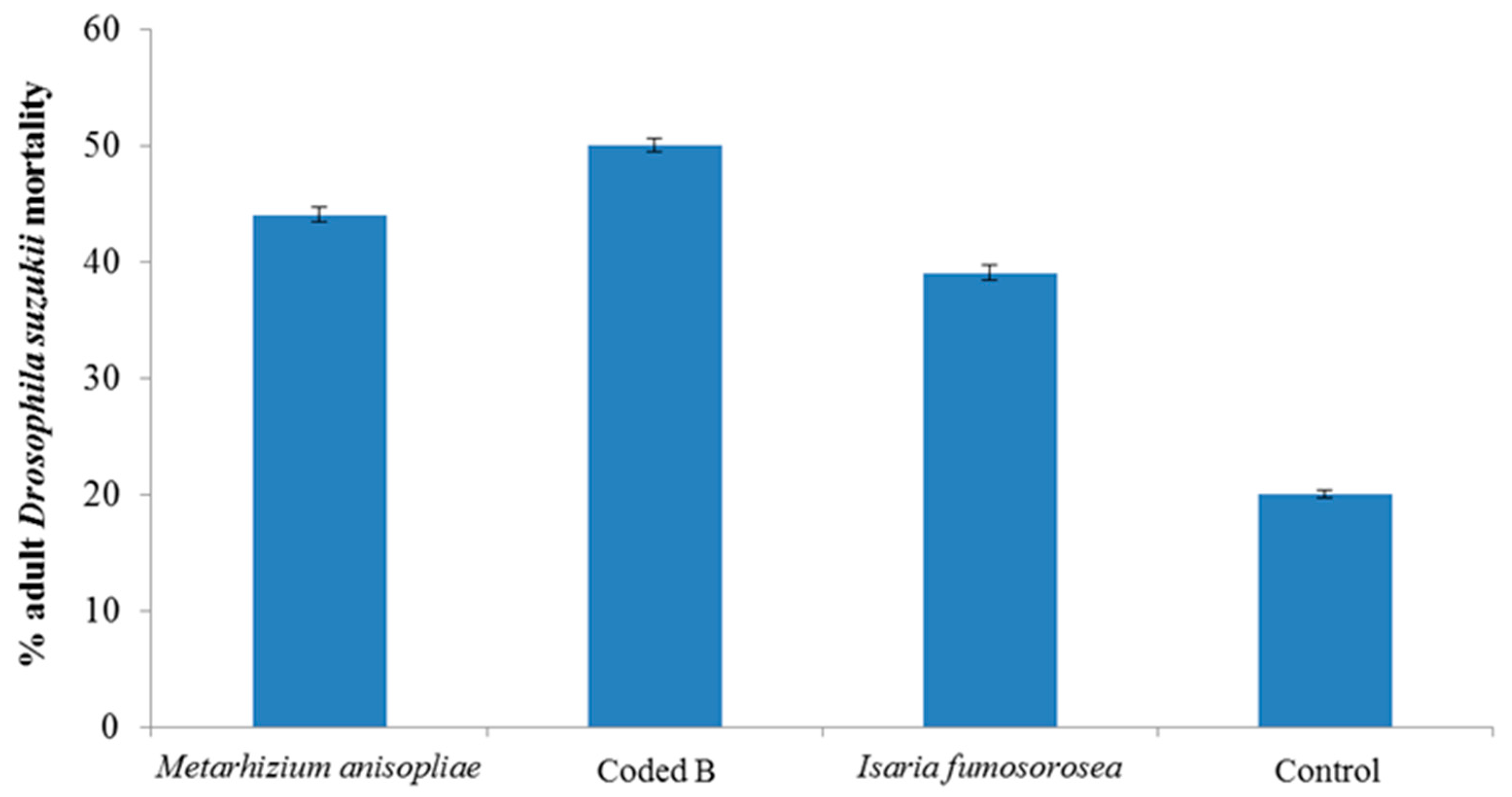
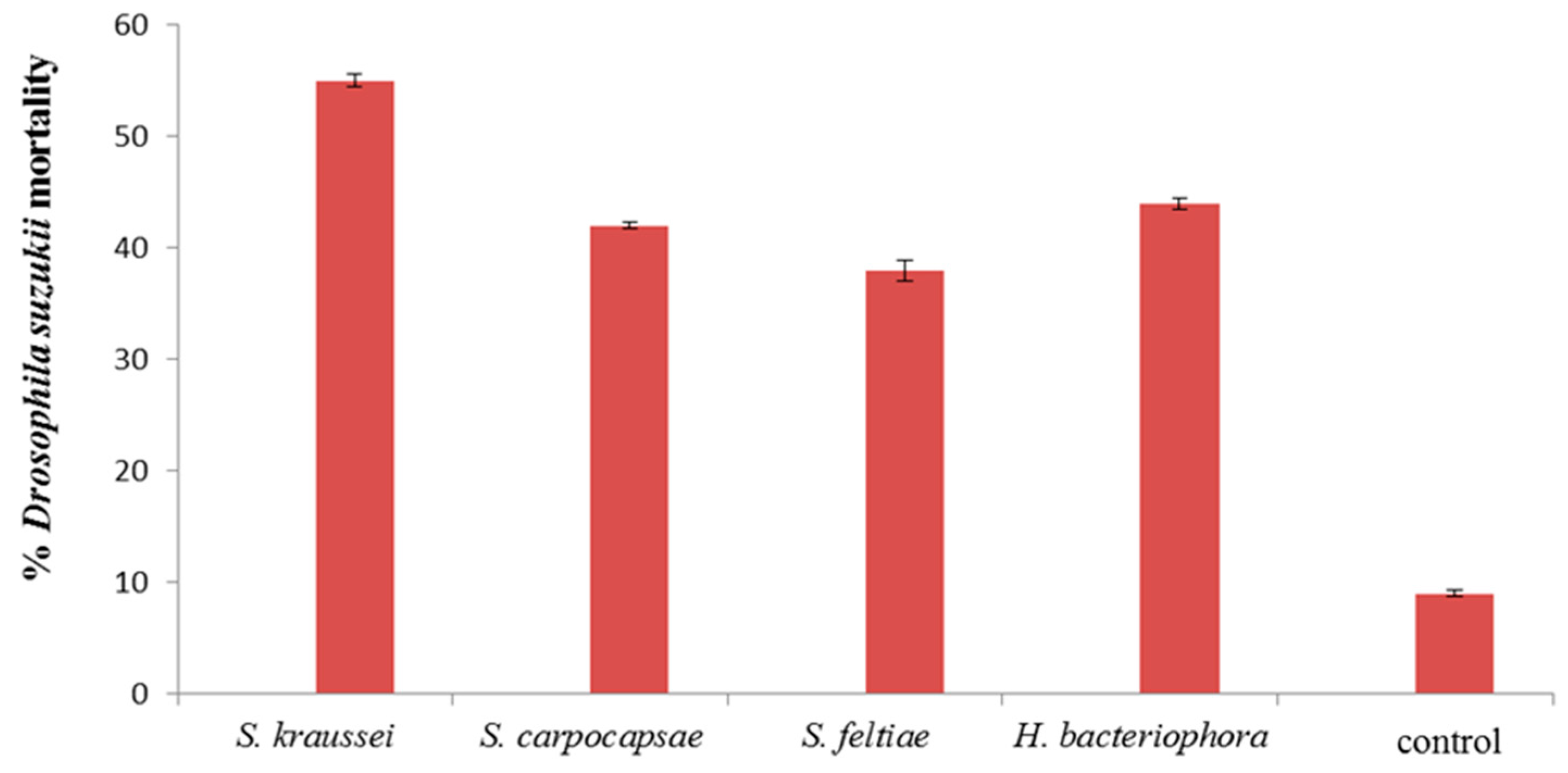
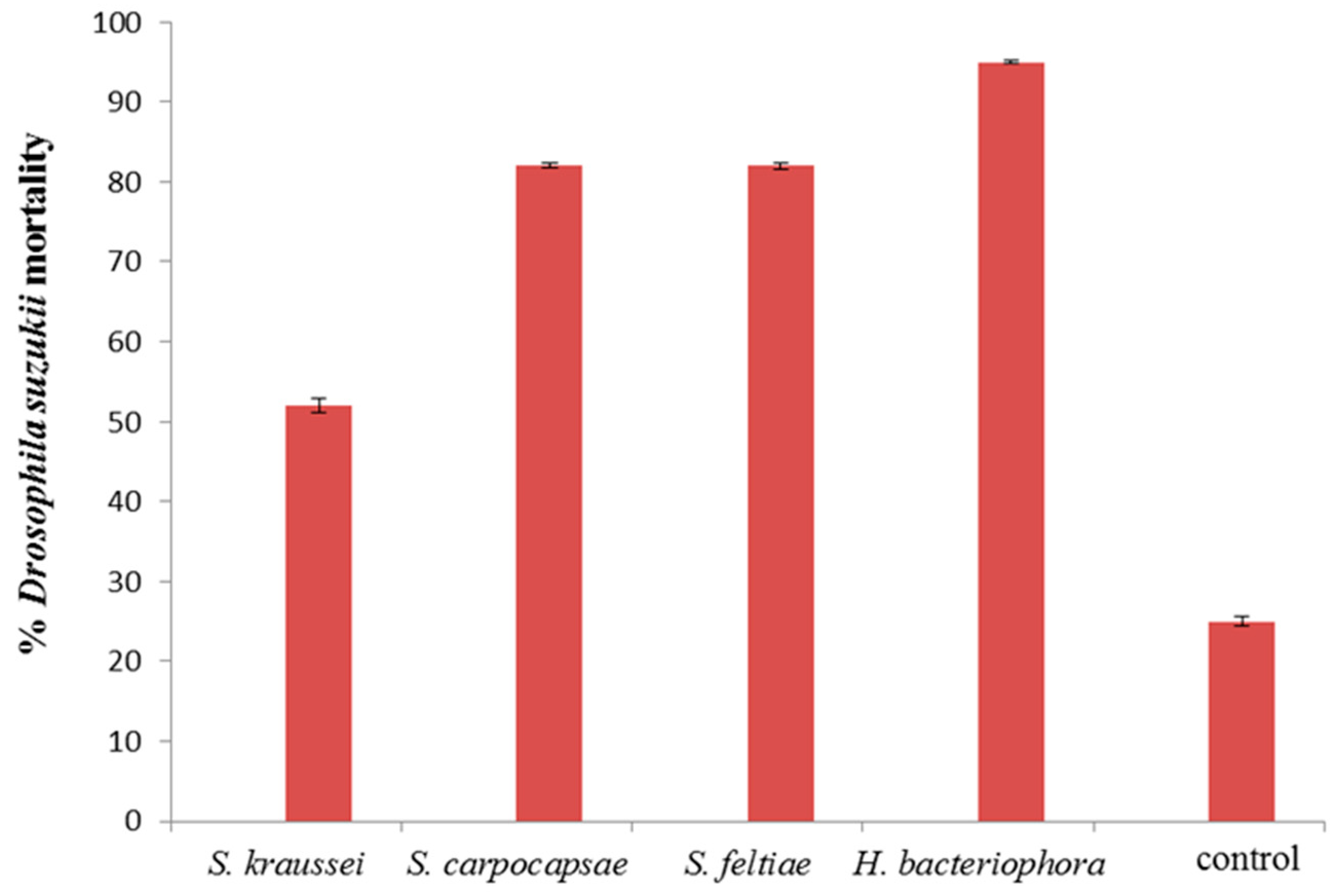
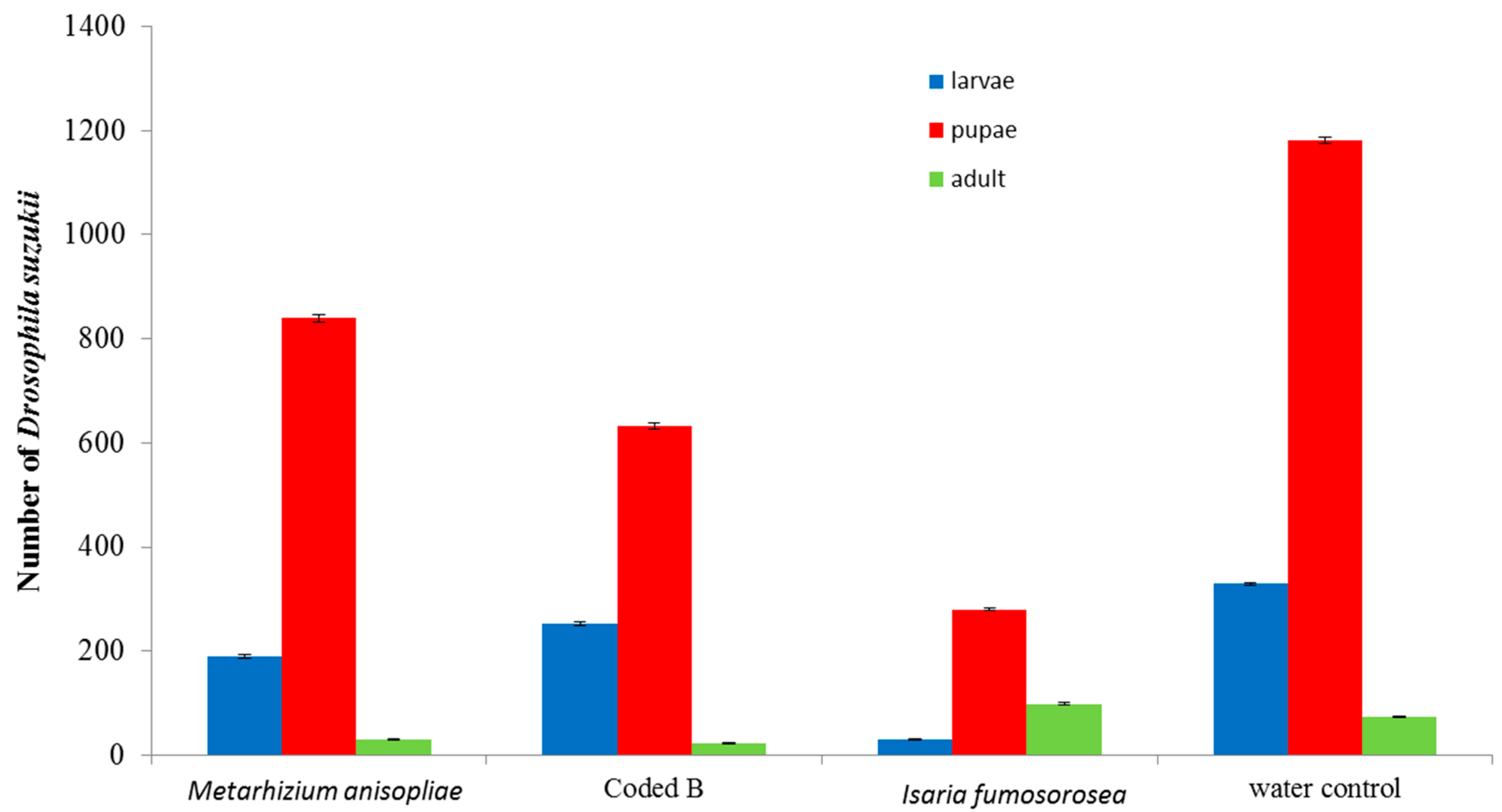
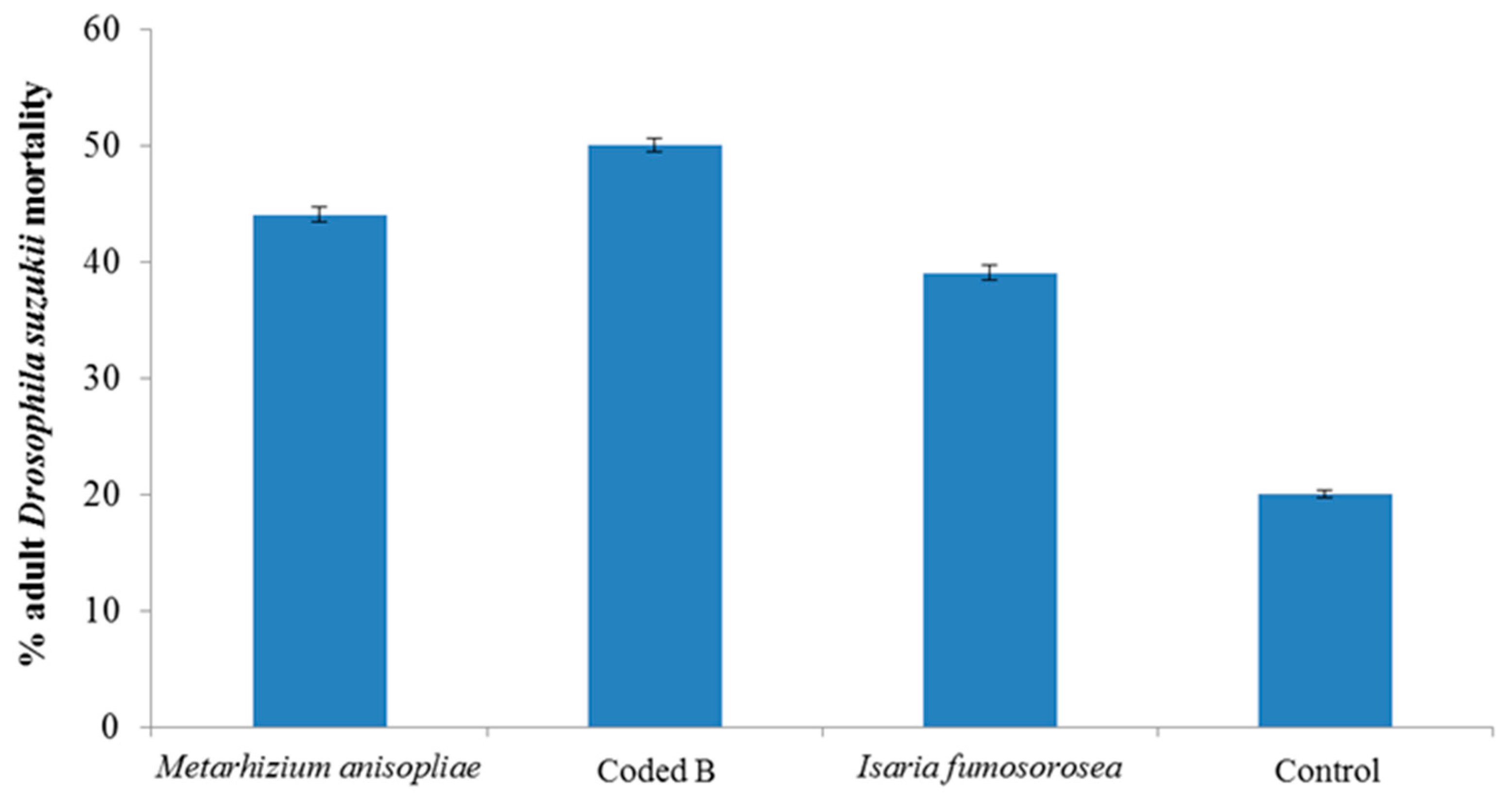
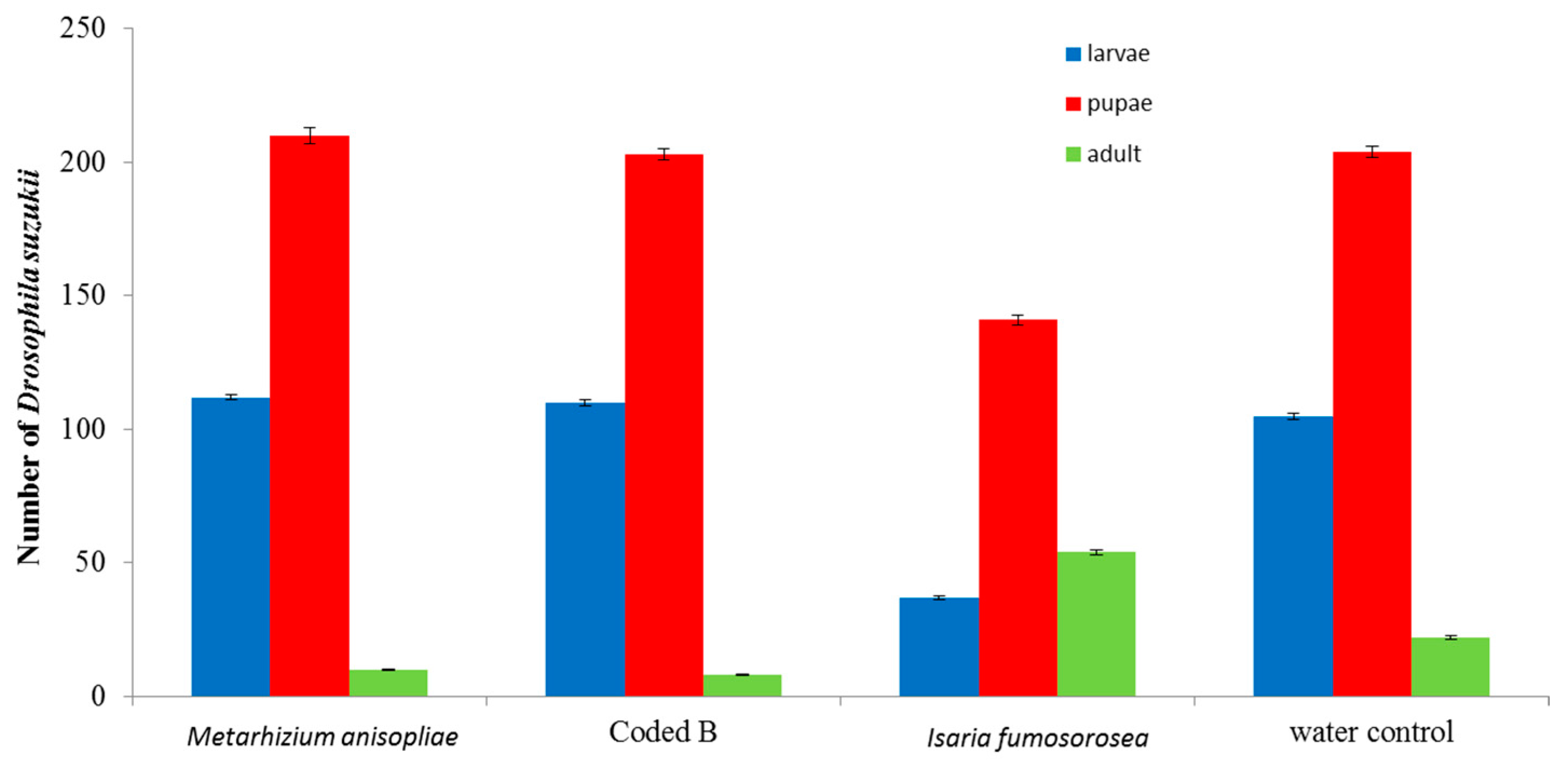
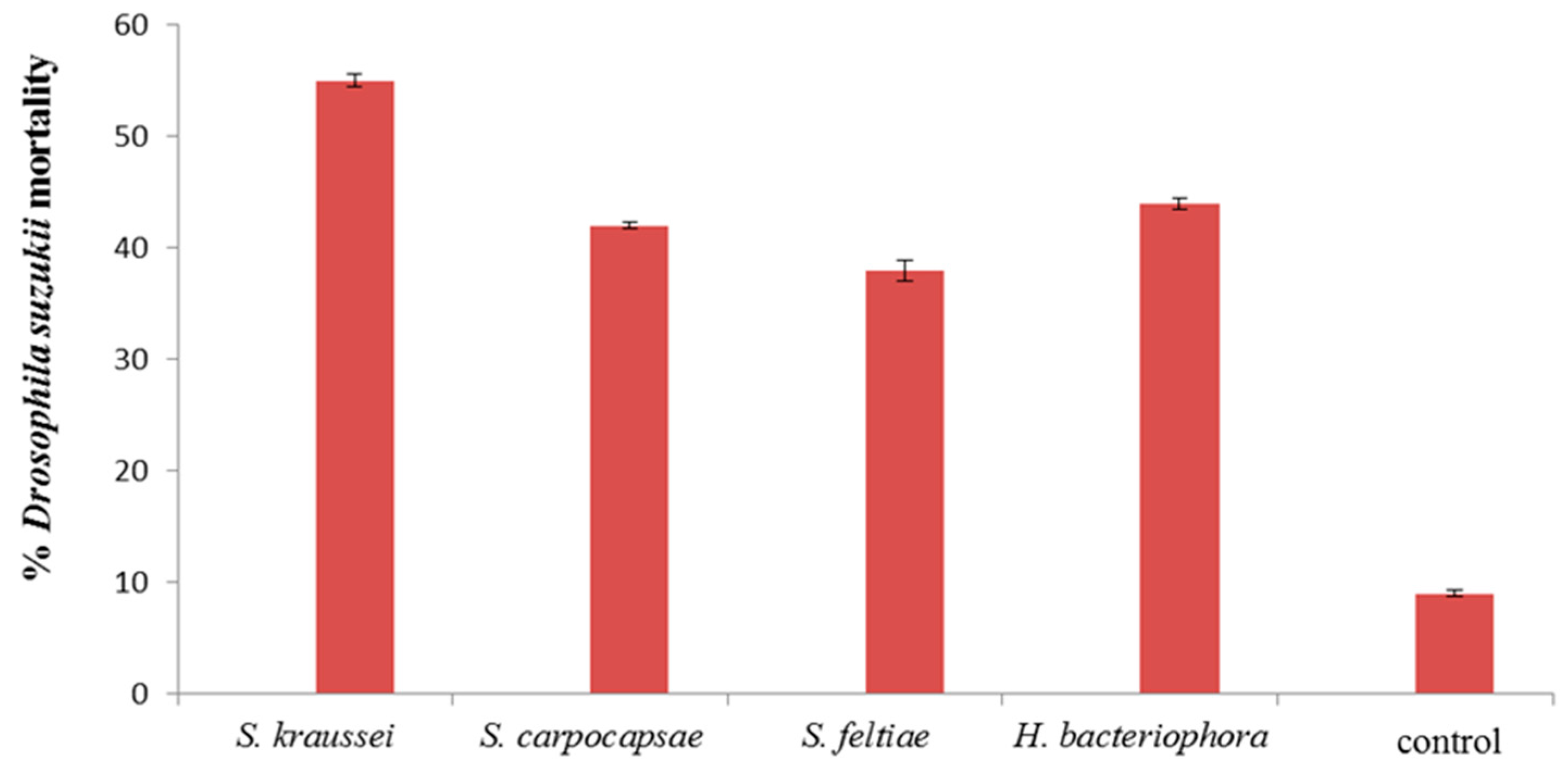
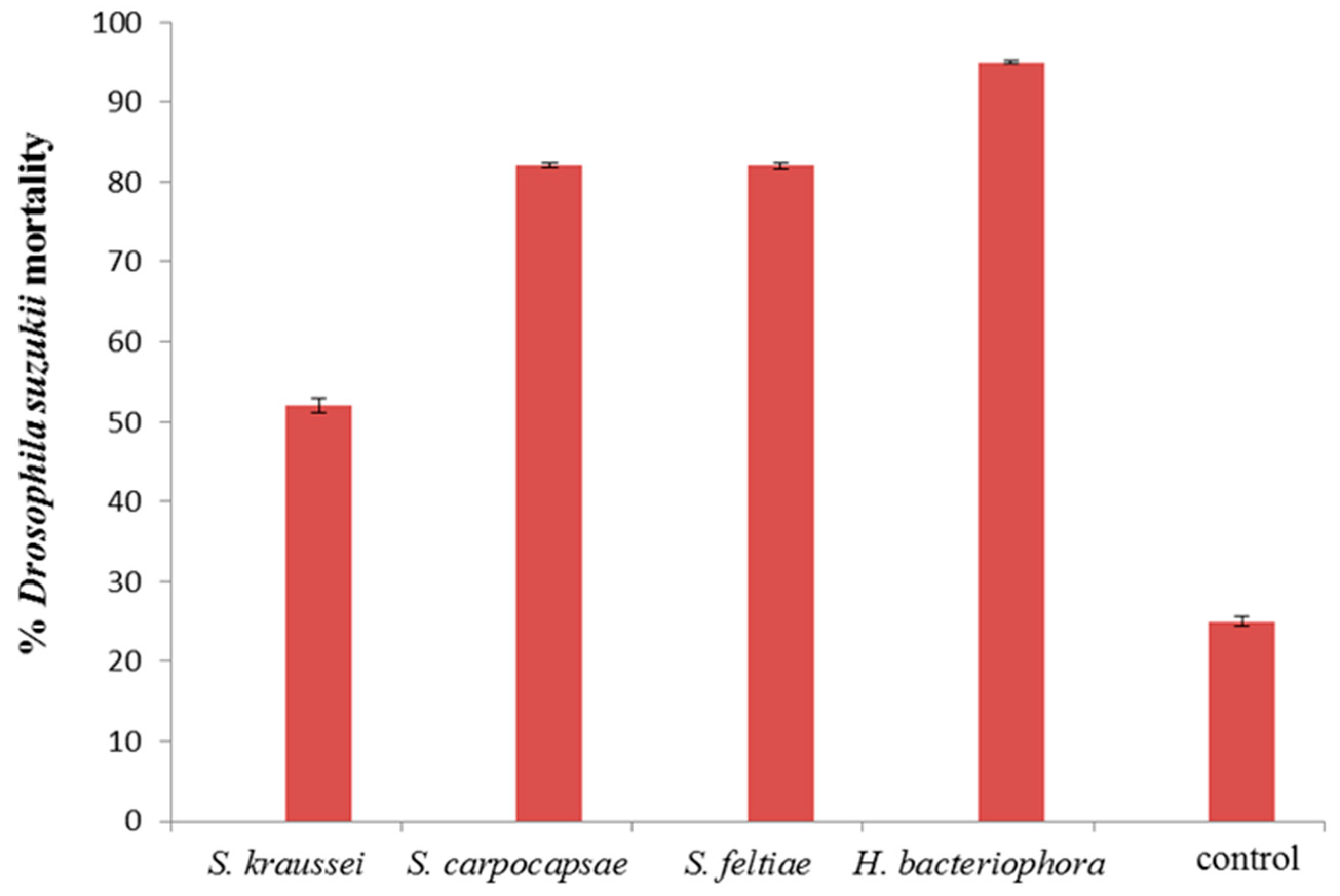
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Toda, M.J. Vertical microdistribution of Drosophilidae (Diptera) within various forests in Hokkaido. III. The Tomakomai Experiment Forest, Hokkaido University. Res. Bull. Coll. Exper. For. 1987, 44, 611–632. [Google Scholar]
- Oku, T. SWD: Drosophila suzukii (Matsumura) in Japan. Agricultural Pest Encyclopedia; Zenkoku Noson Kyoiku: Kyokai, Japan, 2003; pp. 1–381. [Google Scholar]
- Hauser, M.; Gaimari, S.; Damus, M. Drosophila suzukii new to North America. North American Dipterists Society. Available online: http://www.nadsdiptera.org/News/FlyTimes/issue43.pdf (accessed on 1 June 2015).
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest. Manag. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.C.; Valente, V.L. The first records of the invasive pest Drosophila suzukii in the South American continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Calabria, G.; Máca, J.; Bächli, G.; Serra, L.; Pascual, M. First records of the potential pest species Drosophila suzukii (Diptera: Drosophilidae) in Europe. J. Appl. Entomol. 2012, 136, 139–147. [Google Scholar] [CrossRef]
- Goodhue, R. E.; Bolda, M.; Farnsworth, D.; Williams, J. C.; Zalom, F. G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Collins, D.A.; Blackburn, L.F.; Audsley, N.; Bell, H.A. Preliminary screening of potential control products against Drosophila suzukii. Insects 2014, 5, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S; Audsley, N.; Fera, Sand Hutton, York, UK. Personal Observations, 2015.
- Woltz, J.M.; Lee, J.C. Biological control of spotted wing drosophila (Drosophila suzukii), Poster Presentation. In Proceedings of 62nd American Entomological Society Congress, Portland, OR, USA, 16th–19th November 2014.
- Renkema, J.M.; Telfer, Z.; Gariepy, T.; Hallett, R.H. Dalotia coriaria as a predator of Drosophila suzukii: Functional responses, reduced fruit infestation and molecular diagnostics. Biol. Control 2015, 89, 1–10. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Audsley, N. Efficacy of commercially available invertebrate predators against Drosophila suzukii. Insects 2014, 5, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM tactics and their practical implementation in fruit crops across different regions around the world. J. Pest Sci. 2016. [Google Scholar] [CrossRef]
- Mar, T.T.; Lumyong, S. Evaluation of effective entomopathogenic fungi to fruit fly pupa, Bactrocera spp. and their antimicrobial activity. Chiang Mai J. Sci. 2012, 39, 464–477. [Google Scholar]
- Elbashir, M.I.; Bishwajeet, P.; Shankarganesh, K.; Gautam, R.D.; Sharma, P. Pathogenicity of Indian isolates of entomopathogenic fungi against important insect pests and natural enemies. Ind. J. Entomol. 2014, 76, 37–43. [Google Scholar]
- Soliman, N.A.; Ibrahim, A.A.; El-Deen, M.M.; Shams Ramadan, N.F.; Farag, S.R. Entomopathogenic nematodes and fungi as bioControl agents for the peach fruit fly, Bactrocera zonata (Saunders) and the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) soil borne-stages. Egypt. J. Biol. Pest Co. 2014, 24, 497–502. [Google Scholar]
- Yousef, M.; Garrido-Jurado, I.; Quesada-Moraga, E. One Metarhizium brunneum Strain; Two uses to control Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2014, 107, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Poprawski, T.J.; Greenberg, S.M.; Ciomperlik, M.A. Effect of host plant on Beauveria bassiana and Paecilomyces fumosoroseus induced mortality of Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Environ. Entomol. 2000, 29, 1048–1053. [Google Scholar] [CrossRef]
- Naranjo-Lazaro, J.M.; Mellin-Rosas, M.A.; Gonzalez-Padilla, V.D.; Sanchez-Gonzalez, J.A.; Moreno-Carrillo, G.; Arredondo-Bernal, H.C. Susceptibility of Drosophila suzukii Matsumura (Diptera: Drosophilidae) to entomopathogenic fungi. Southwest. Entomol. 2014, 39, 201–203. [Google Scholar] [CrossRef]
- Poinar, G.O. JR Biology and taxonomy of Steinernematidae and Heterorhabditidae. In Entomopathogenic Nematodes in Biological Control; Gaugler, R., Kaya, H.K., Eds.; CRC Press Inc.: Boca Raton, FL, USA, 1990; pp. 23–61. [Google Scholar]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Cross, J.V.; Solomon, M.G.; Chandler, D.; Jarrett, P.; Richardson, P.N.; Winstanley, D.; Bathon, H.; Huber, J.; Keller, B.; Langenbruch, G.A.; et al. Biocontrol of pests of apples and pears in Northern and Central Europe: 1. Microbial agents and nematodes. Biocontrol Sci. Techn. 1999, 9, 125–149. [Google Scholar] [CrossRef]
- Krishnayya, P.V.; Grewal, P.S. Effect of neem and selected fungicides on viability and virulence of the entomopathogenic nematode Steinernema feltiae. Biocontrol Sci. Techn. 2002, 12, 259–266. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Head, J.; Walters, K.F.A.; Murray, A.W.A. The integrated use of chemical insecticides and the entomopathogenic nematode, Steinernema feltiae, for the control of the sweetpotato whitefly, Bemisia tabaci. Nematology 2003, 5, 713–720. [Google Scholar] [CrossRef]
- Williams, E.C.; Walters, K.F.A. Foliar application of the entomopathogenic nematode Steinernema feltiae against leafminers on vegetables. Biocontrol Sci. Technol. 2000, 10, 61–70. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Northing, P.; Luo, W. Efficacy of the entomopathogenic nematode, Steinernema feltiae, against sweetpotato whitefly, Bemisia tabaci, under laboratory and glasshouse conditions. Bull. Entomol. Res. 2007, 97, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Mathers, J.J.; Northing, P.; Luo, W.; Walters, K.F.A. The susceptibility of immature stages of Bemisia tabaci to infection by the entomopathogenic nematode Steinernema carpocapsae. Russ. J. Nematol. 2007, 15, 153–156. [Google Scholar]
- Chyzik, R.; Glazer, O.; Klein, M. Virulence and efficacy of different entomopathogenic nematode species against western flower thrips (Frankliniella occidentalis). Phytoparasitica 1996, 24, 103–110. [Google Scholar] [CrossRef]
- Ebssa, L.; Borgemister, C.; Berndt, O.; Poehling, H.M. Efficacy of entomopathogenic nematodes against soil-dwelling life stages of western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). J. Invertebr. Pathol. 2001, 78, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Funderburk, J.; Stavisky, J.; Tipping, C.; Gorbet, D.; Momol, T.; Berger, R. Infection of Frankliniella fusca (Thysanoptera: Thripidae) in peanut by the parasitic nematode Thripinema fuscum (Tylenchidae: Allantonematidae). Environ. Entomol. 2002, 31, 558–563. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; North, J.P.; Walters, K.F.A. Effect of temperature and host plant leaf morphology on the efficacy of two entomopathogenic biocontrol agents of Thrips palmi (Thysanoptera: Thripidae). Bull. Entomol. Res. 2005, 95, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Mathers, J.J.; Blackburn, L.F.; Powell, M.E.; Marris, G.; Pietravalle, S.; Brown, M.A.; Budge, G.E. Screening commercially available entomopathogenic biocontrol agents for the control of Aethina tumida (Coleoptera: Nitidulidae) in the UK. Insects 2012, 3, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, A.G.S.; Walters, K.F.A. Evaluation of exposure time of Steinernema feltiae against second instar Bemisia tabaci. Tests Agrochem. Cult. 2005, 26, 34–35. [Google Scholar]
- Castillo, M.A.; Moya, P.; Herna´ndez, E.; Primo-Yu´fera, E. Susceptibility of Ceratitis capitata Wiedemann (Diptera: Tephritidae) to entomopathogenic fungi and their extracts. Biol. Control 2000, 19, 274–282. [Google Scholar] [CrossRef]
- Ekesi, S.; Maniania, N.K.; Lux, S.A. Mortality in three African Tephritid fruit fly puparia and adults caused by the entomopathogenic fungi, Metarhizium anisopliae and Beauveria bassiana. Biocontrol Sci. Techn. 2002, 12, 7–17. [Google Scholar] [CrossRef]
- Dimbi, S.; Maniania, N.K.; Lux, S.A.; Ekesi, S.; Mueke, J.K. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathologia 2003, 156, 375–382. [Google Scholar] [PubMed]
- Konstantopoulou, M.A.; Mazomenos, B.E. Evaluation of Beauveria bassiana and B. brongniartii strains and four wild-type fungal species against adults of Bactrocera oleae and Ceratitis capitata. Biocontrol 2005, 50, 293–305. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Ruiz-García, A.; Santiago-Álvarez, C. Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2006, 99, 1955–1966. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.J.; Willis, O.R. Some factors affecting parasitism by mermithid nematodes in southern house mosquito larvae. J. Econ. Entomol. 1970, 63, 175–178. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Head, J.; Walters, K.F.A.; Gregory, S.A. The efficacy of the entomopathogenic nematode, Steinernema feltiae, against the immature stages of Bemisia tabaci. J. Invertebr. Pathol. 2003, 83, 267–269. [Google Scholar] [CrossRef]
- Woltz, J.M.; Donahue, K.M.; Bruck, D.J.; Lee, J.C. Efficacy of commercially available predators, nematodes and fungal entomopathogens for augmentative control of Drosophila suzukii. J. Appl. Entomol. 2015, 139, 759–770. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S. Update on the status of Bemisia tabaci in the UK and the use of entomopathogenic fungi within eradication programmes. Insects 2013, 4, 198–205. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuthbertson, A.G.S.; Audsley, N. Further Screening of Entomopathogenic Fungi and Nematodes as Control Agents for Drosophila suzukii. Insects 2016, 7, 24. https://doi.org/10.3390/insects7020024
Cuthbertson AGS, Audsley N. Further Screening of Entomopathogenic Fungi and Nematodes as Control Agents for Drosophila suzukii. Insects. 2016; 7(2):24. https://doi.org/10.3390/insects7020024
Chicago/Turabian StyleCuthbertson, Andrew G. S., and Neil Audsley. 2016. "Further Screening of Entomopathogenic Fungi and Nematodes as Control Agents for Drosophila suzukii" Insects 7, no. 2: 24. https://doi.org/10.3390/insects7020024
APA StyleCuthbertson, A. G. S., & Audsley, N. (2016). Further Screening of Entomopathogenic Fungi and Nematodes as Control Agents for Drosophila suzukii. Insects, 7(2), 24. https://doi.org/10.3390/insects7020024