
Fungal Transformation of Tree Stumps into a Suitable Resource for Xylophagous Beetles via Changes in Elemental Ratios


Abstract
:1. Introduction
1.1. Nutritional Scarcity of Pure Wood
1.2. Nutritional Enrichment of Decomposing Wood Meets the Needs of Wood-Eaters
1.3. Searching for the Mechanisms Underlying Wood Enrichment During Decomposition
1.4. Aims of This Study
2. Materials and Methods
2.1. Dead-Wood Nutritional Enrichment
2.2. Nutritional Limitation of Xylophages
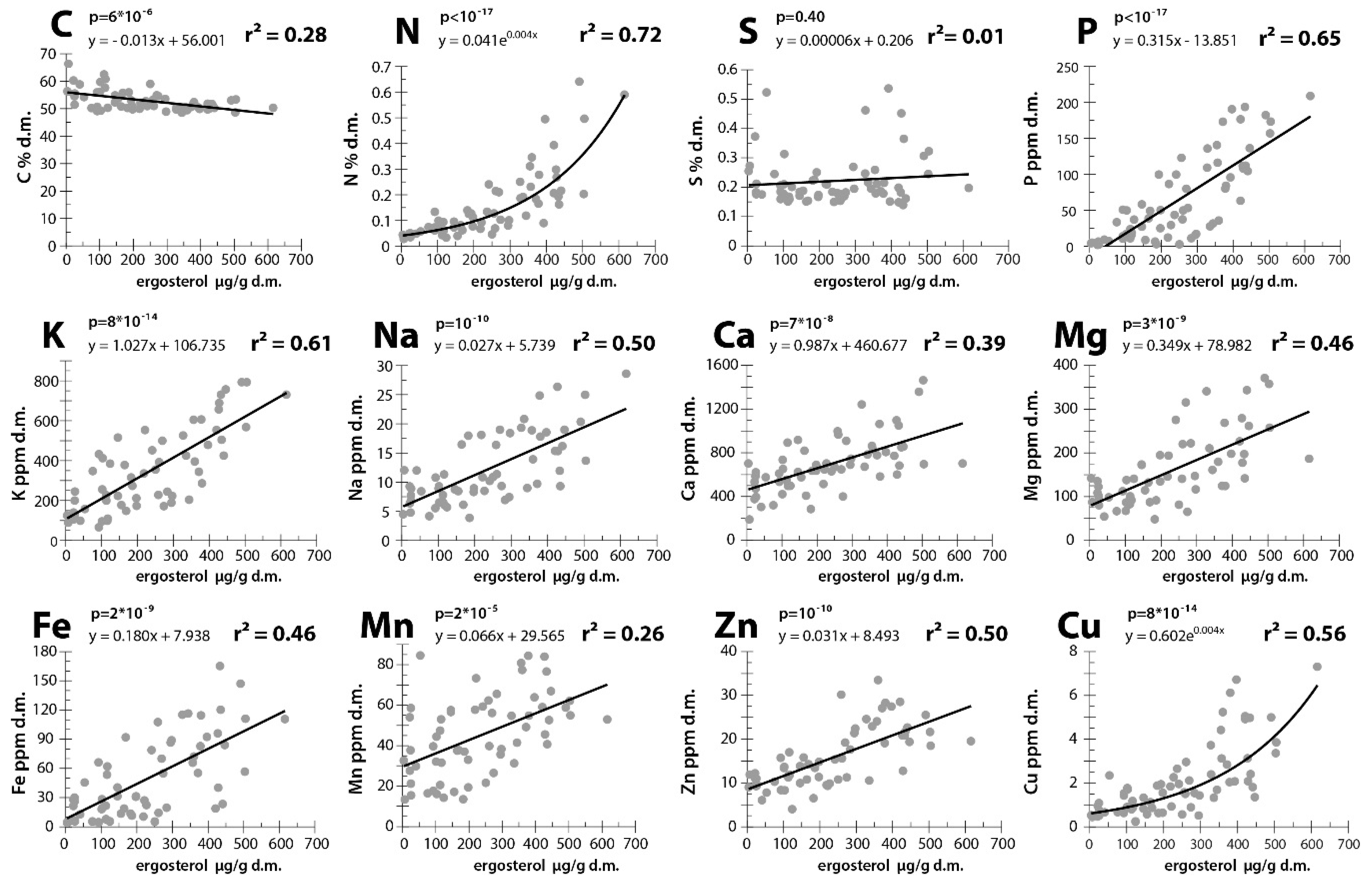
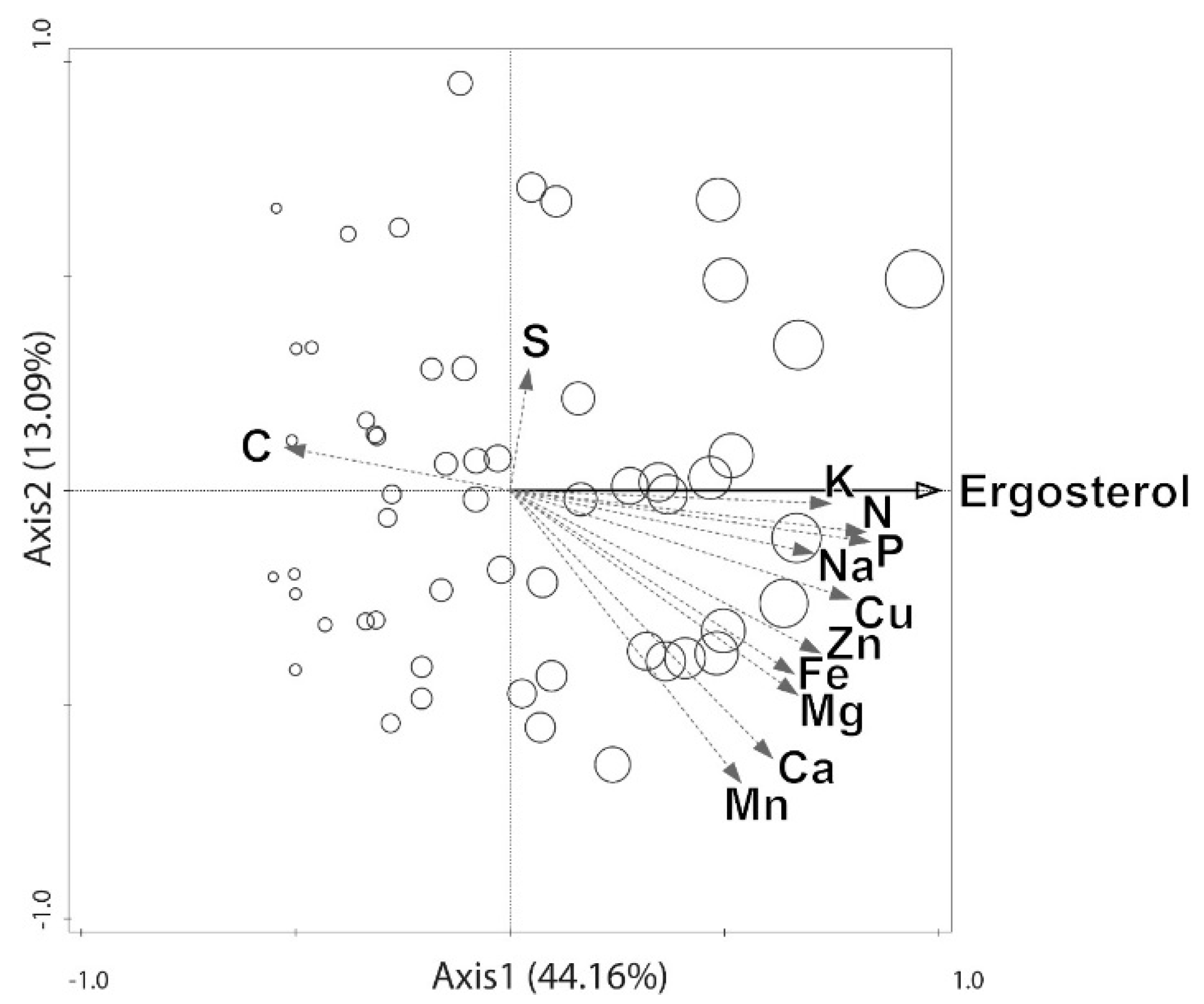
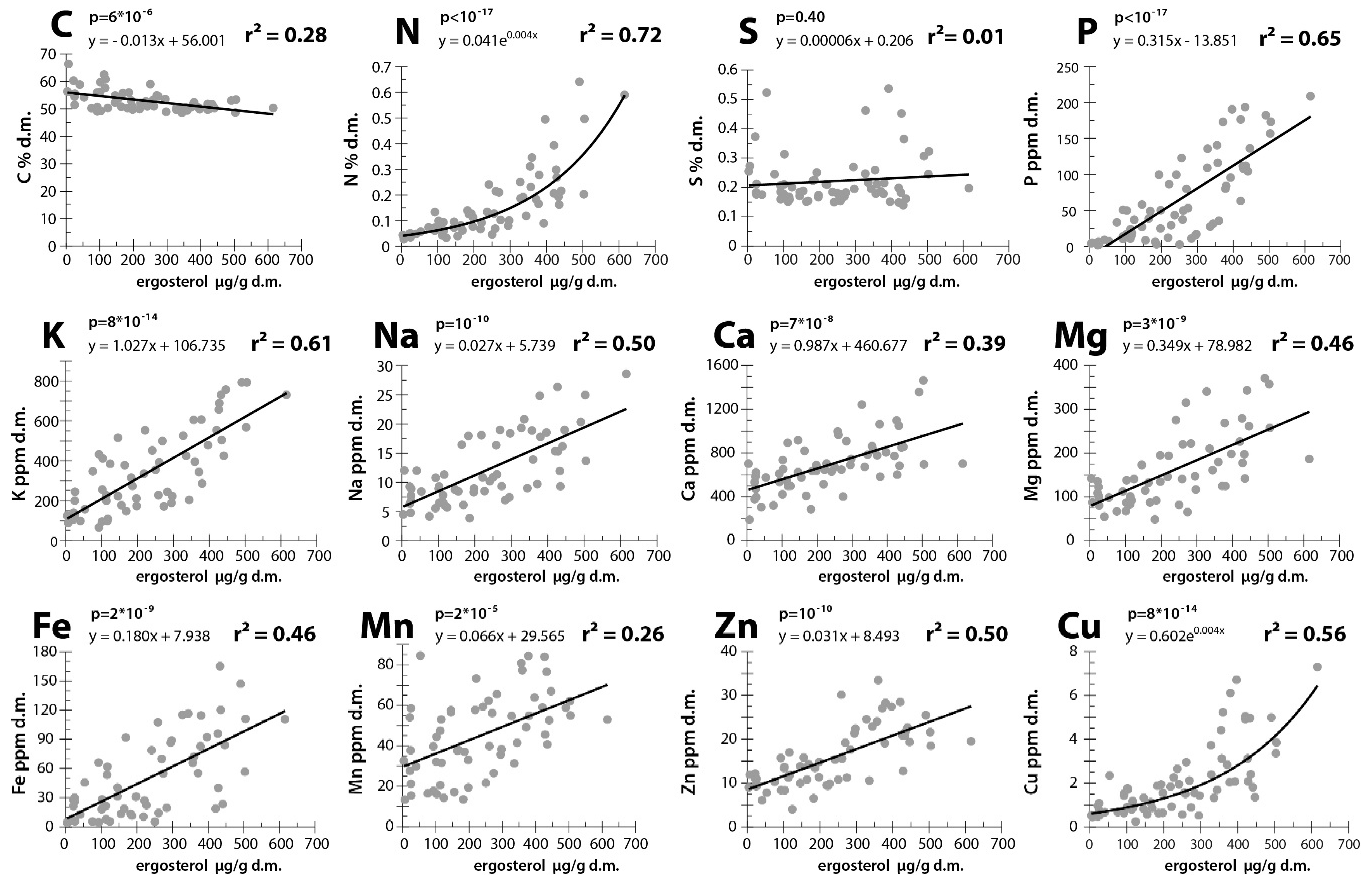
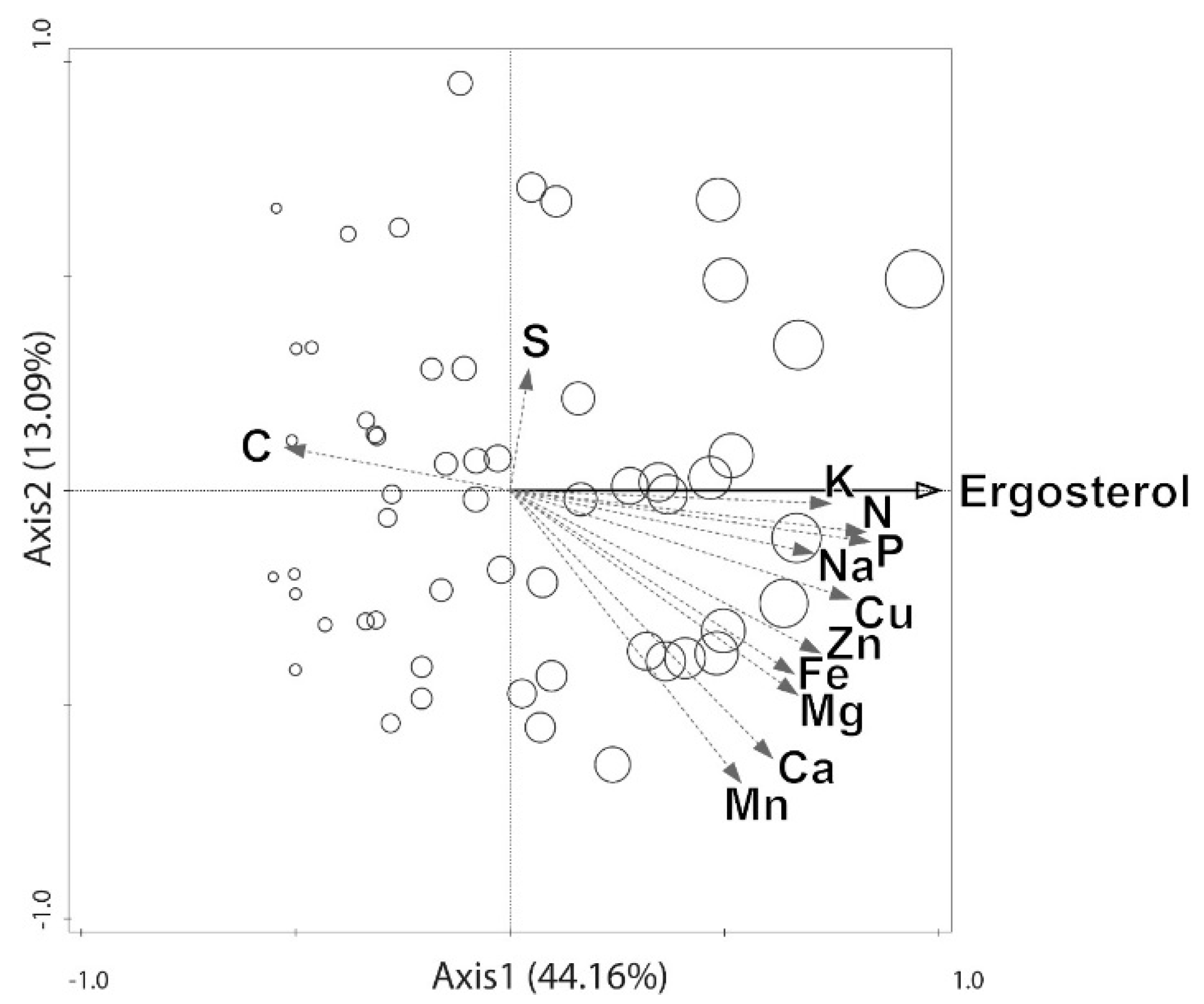
3. Results
4. Discussion
4.1. Elements That May be Transported by Fungi to Dead Wood From the Outside Environment
4.2. Nutritional Enrichment of Dead Wood by Fungi Allows for the Development of Wood-Eaters
4.3. Nutrient Cycling Pathways and Dead Wood Fungi
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| RDA | Multivariate redundancy analysis |
| AIC | Akaike information criterion |
References
- Filipiak, M.; Weiner, J. How to make a beetle out of wood: Multi-elemental stoichiometry of wood decay, xylophagy and fungivory. PLoS ONE 2014, 9, e115104. [Google Scholar]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of Coarse Woody Debris in Temperate Ecosystems. Adv. Ecol. Res. 1986. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Phosphorus and base cation accumulation and release patterns in decomposing Scots pine, Norway spruce and silver birch stumps. For. Ecol. Manage. 2010, 260, 1478–1489. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Carbon and nitrogen release from decomposing Scots pine, Norway spruce and silver birch stumps. For. Ecol. Manage. 2010, 259, 390–398. [Google Scholar] [CrossRef]
- Meerts, P. Mineral nutrient concentrations in sapwood and heartwood: A literature review. Ann. For. Sci. 2002, 59, 713–722. [Google Scholar] [CrossRef]
- Köster, K.; Metslaid, M.; Engelhart, J.; Köster, E. Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests. For. Ecol. Manage. 2015, 354, 35–42. [Google Scholar] [CrossRef]
- Denno, R.F.; Fagan, W.F. Might nitrogen limitation promote omnivory among carnivorous arthropods? Ecology 2003, 84, 2522–2531. [Google Scholar] [CrossRef]
- Hessen, D.O.; Elser, J.J.; Sterner, R.W.; Urabe, J. Ecological stoichiometry: An elementary approach using basic principles. Limnol. Oceanogr. 2013, 58, 2219–2236. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Fagan, W.F.; Siemann, E.; Mitter, C.; Denno, R.F.; Huberty, A.F.; Woods, H.A.; Elser, J.J. Nitrogen in insects: Implications for trophic complexity and species diversification. Am. Nat. 2002, 160, 784–802. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Kay, A.D.; Fagan, W.F. Adaptation to a limiting environment: the phosphorus content of terrestrial cave arthropods. Ecol. Res. 2010, 25, 565–577. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food. Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Ljungdahl, L.G.; Eriksson, K.-E. Ecology of Microbial Cellulose Degradation. Adv. Microb. Ecol. 1985, 8, 237–299. [Google Scholar]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Roskoski, J.P. Nitrogen fixation in hardwood forests of the northeastern United States. Plant Soil 1980, 54, 33–44. [Google Scholar] [CrossRef]
- Boddy, L.; Watkinson, S.C. Wood decomposition, higher fungi, and their role in nutrient redistribution. Can. J. Bot. 1995, 73, 1377–1383. [Google Scholar] [CrossRef]
- Stark, N. Nutrient Cycling Pathways and Litter Fungi. Bioscience 1972, 22, 355–360. [Google Scholar] [CrossRef]
- Foster, J.R.; Lang, G.E. Decomposition of red spruce and balsam fir boles in the White Mountains of New Hampshire. Can. J. For. Res. 1982, 12, 617–626. [Google Scholar]
- Dighton, J. Fungi in Ecosystem Processes; CRC Press: New York, NY, USA, 2003. [Google Scholar]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014. [Google Scholar] [CrossRef]
- Clinton, P.W.; Buchanan, P.K.; Wilkie, J.P.; Smaill, S.J.; Kimberley, M.O. Decomposition of Nothofagus wood in vitro and nutrient mobilization by fungi. Can. J. For. Res. 2009, 39, 2193–2202. [Google Scholar] [CrossRef]
- Dighton, J. Nutrient cycling by saprotrophic fungi in terrestrial habitats. In Environmental and Microbial Relationships; Kubicek, C.P., Druzhinina, I.S., Eds.; Springer Berlin Heidelberg: Berlin, Germany, 2007; pp. 287–300. [Google Scholar]
- Cairney, J. Basidiomycete mycelia in forest soils: Dimensions, dynamics and roles in nutrient distribution. Mycol. Res. 2005, 109, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Watkinson, S.C.; Bebber, D.; Darrah, P.; Fricker, M.; Tlalka, M.; Boddy, L. The role of wood decay fungi in the carbon and nitrogen dynamics of the forest floor. In Fungi in Biogeochemical Cycles; Gadd, G.M., Ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 151–181. [Google Scholar]
- Lodge, D.J. Nutrient concentrations, percentage moisture and density of field-collected fungal mycelia. Soil Biol. Biochem. 1987, 19, 727–733. [Google Scholar] [CrossRef]
- Rudawska, M.; Leski, T. Macro- and micro-element contents in fruiting bodies of wild mushrooms from the Notecka forest in west-central Poland. Food Chem. 2005, 92, 499–506. [Google Scholar] [CrossRef]
- Dursun, N.; Özcan, M.M.; Kaşık, G.; Öztürk, C. Mineral contents of 34 species of edible mushrooms growing wild in Turkey. J. Sci. Food. Agric. 2006, 86, 1087–1094. [Google Scholar] [CrossRef]
- Falandysz, J.; Borovička, J. Macro and trace mineral constituents and radionuclides in mushrooms: Health benefits and risks. Appl. Microbiol. Biotechnol. 2013, 97, 477–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zhang, J.; Wu, L.H.; Zhao, Y.L.; Li, T.; Li, J.Q.; Wang, Y.Z.; Liu, H.G. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Palviainen, M.; Finér, L. Decomposition and nutrient release from Norway spruce coarse roots and stumps––A 40-year chronosequence study. For. Ecol. Manage. 2015, 358, 1–11. [Google Scholar] [CrossRef]
- Gessner, M.O. Ergosterol as a measure of fungal biomass. In Methods to Study Litter Decomposition; Graça, M.A.S., Bärlocher, F., Gessner, M.O., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2005; pp. 189–195. [Google Scholar]
- Mille-Lindblom, C.; von Wachenfeldt, E.; Tranvik, L.J. Ergosterol as a measure of living fungal biomass: Persistence in environmental samples after fungal death. J. Microbiol. Methods. 2004, 59, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Niemenmaa, O.; Galkin, S.; Hatakka, A. Ergosterol contents of some wood-rotting basidiomycete fungi grown in liquid and solid culture conditions. Int. Biodeterior. Biodegradation 2008, 62, 125–134. [Google Scholar] [CrossRef]
- Stahl, P.D.; Parkin, T.B. Relationship of soil ergosterol concentration and fungal biomass. Soil Biol. Biochem. 1996, 28, 847–855. [Google Scholar] [CrossRef]
- Russell, M.B.; Woodall, C.W.; Fraver, S.; D’Amato, A.W.; Domke, G.M.; Skog, K.E. Residence times and decay rates of downed woody debris biomass/Carbon in Eastern US forests. Ecosystems 2014, 17, 765–777. [Google Scholar] [CrossRef]
- Fukasawa, Y.; Katsumata, S.; Mori, A.S.; Osono, T.; Takeda, H. Accumulation and decay dynamics of coarse woody debris in a Japanese old-growth subalpine coniferous forest. Ecol. Res. 2014, 29, 257–269. [Google Scholar] [CrossRef]
- Tobin, B.; Black, K.; McGurdy, L.; Nieuwenhuis, M. Estimates of decay rates of components of coarse woody debris in thinned Sitka spruce forests. Forestry 2007, 80, 455–469. [Google Scholar] [CrossRef]
- Garrett, L.G.; Oliver, G.R.; Pearce, S.H.; Davis, M.R. Decomposition of Pinus radiata coarse woody debris in New Zealand. For. Ecol. Manage. 2008, 255, 3839–3845. [Google Scholar] [CrossRef]
- Hessen, D.O.; Anderson, T.R. Excess carbon in aquatic organisms and ecosystems: Physiological, ecological, and evolutionary implications. Limnol. Oceanogr. 2008, 53, 1685–1696. [Google Scholar] [CrossRef]
- Anderson, T.R.; Hessen, D.O.; Elser, J.J.; Urabe, J. Metabolic stoichiometry and the fate of excess carbon and nutrients in consumers. Am. Nat. 2005, 165, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Frost, P.C.; Benstead, J.P.; Cross, W.F.; Hillebrand, H.; Larson, J.H.; Xenopoulos, M.A.; Yoshida, T. Threshold elemental ratios of carbon and phosphorus in aquatic consumers. Ecol. Lett. 2006, 9, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Walczyńska, A. Is wood safe for its inhabitants? Bull. Entomol. Res. 2010, 100, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Walczyńska, A. How does a xylem-feeder maximize its fitness? Bull. Entomol. Res. 2012, 102, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Graça, M.A.S. The role of invertebrates on leaf litter decomposition in streams––A review. Int. Rev. Hydrobiol. 2001, 86, 383–393. [Google Scholar] [CrossRef]
- Martius, C. Decomposition of wood. In The Central Amazon Floodplain; Springer Berlin Heidelberg: Berlin, Germany, 1997; pp. 267–276. [Google Scholar]

{kind=link}
{kind=link}
| Stoichiometric Mismatch (TSR) | N | P | K | Na | Ca | Mg | Fe | Zn | Mn | Cu | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Females | S. rubra TSRL | 258.6 | 1772.9 | 70.6 | 133.5 | 2.6 | 15.9 | 11.0 | 11.6 | 0.7 | 78.7 |
| S. rubra TSRH | 19.5 | 36.9 | 9.7 | 39.8 | 1.0 | 5.7 | 1.2 | 5.1 | 0.4 | 9.3 | |
| A. rusticus TSRL | 133.3 | 1252.1 | 54.8 | 104.7 | 1.6 | 12.9 | 6.2 | 12.6 | 0.5 | 68.5 | |
| A. rusticus TSRH | 10.0 | 26.1 | 7.5 | 31.2 | 0.7 | 4.6 | 0.7 | 5.5 | 0.2 | 8.1 | |
| C. mariana TSRL | 207.5 | 1790.2 | 89.0 | 85.2 | 2.1 | 25.4 | 2.8 | 8.0 | 0.8 | 12.2 | |
| C. mariana TSRH | 15.6 | 37.3 | 12.2 | 25.4 | 0.9 | 9.1 | 0.3 | 3.5 | 0.4 | 1.5 | |
| Males | S. rubra TSRL | 206.3 | 1052.2 | 54.6 | 101.5 | 2.1 | 13.8 | 3.9 | 10.7 | 0.8 | 30.7 |
| S. rubra TSRH | 15.5 | 21.9 | 7.5 | 30.3 | 0.8 | 4.9 | 0.4 | 4.7 | 0.4 | 3.6 | |
| A. rusticus TSRL | 177.6 | 1458.0 | 72.9 | 90.7 | 1.7 | 20.3 | 3.3 | 7.5 | 0.9 | 11.7 | |
| A. rusticus TSRH | 13.4 | 30.3 | 10.0 | 27.1 | 0.7 | 7.3 | 0.4 | 3.3 | 0.4 | 1.4 | |
| C. mariana TSRL | 244.4 | 1669.5 | 70.3 | 176.2 | 1.9 | 18.3 | 4.1 | 8.7 | 0.7 | 15.2 | |
| C. mariana TSRH | 18.4 | 34.7 | 9.6 | 52.6 | 0.8 | 6.6 | 0.5 | 3.8 | 0.3 | 1.8 | |
| α | 13.3 | 48.1 | 7.3 | 3.4 | 2.5 | 2.8 | 9.1 | 2.3 | 2.1 | 8.4 |
| Value of MF/MI | Values of β for the Measured Elements and Ergosterol | ||||||||||||
| C | N | S | P | K | Na | Ca | Mg | Fe | Zn | Mn | Cu | erg | |
| 0.90 | 0.8 | 10.3 | 0.9 | 37.3 | 5.7 | 2.6 | 1.9 | 2.2 | 7.0 | 1.8 | 1.6 | 6.5 | 28.8 |
| 0.58 | 0.5 | 6.6 | 0.6 | 24.0 | 3.7 | 1.7 | 1.2 | 1.4 | 4.5 | 1.1 | 1.0 | 4.2 | 18.6 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filipiak, M.; Sobczyk, Ł.; Weiner, J. Fungal Transformation of Tree Stumps into a Suitable Resource for Xylophagous Beetles via Changes in Elemental Ratios. Insects 2016, 7, 13. https://doi.org/10.3390/insects7020013
Filipiak M, Sobczyk Ł, Weiner J. Fungal Transformation of Tree Stumps into a Suitable Resource for Xylophagous Beetles via Changes in Elemental Ratios. Insects. 2016; 7(2):13. https://doi.org/10.3390/insects7020013
Chicago/Turabian StyleFilipiak, Michał, Łukasz Sobczyk, and January Weiner. 2016. "Fungal Transformation of Tree Stumps into a Suitable Resource for Xylophagous Beetles via Changes in Elemental Ratios" Insects 7, no. 2: 13. https://doi.org/10.3390/insects7020013
APA StyleFilipiak, M., Sobczyk, Ł., & Weiner, J. (2016). Fungal Transformation of Tree Stumps into a Suitable Resource for Xylophagous Beetles via Changes in Elemental Ratios. Insects, 7(2), 13. https://doi.org/10.3390/insects7020013