
Feeding Behaviour on Host Plants May Influence Potential Exposure to Bt Maize Pollen of Aglais Urticae Larvae (Lepidoptera, Nymphalidae)

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Sites and Nettle Patches

{kind=link}
{kind=link}
{kind=link}
| Sites | Sample Years | No. of Nettle Patches | Size of Patches (m2) | Nettle Density (Plants/m2) |
|---|---|---|---|---|
| Steinen (G) | 2011, 2013 | 6 | 22.67 ± 4.58 | 118.17 ± 11.32 |
| Todtnau (G) | 2013 | 1 | 20 | 45 |
| St. Sigmund (A) | 2012 | 1 | 8 | 50 |
| Praxmar (A) | 2012 | 1 | 3 | 120 |
2.2. Butterfly Larvae in the Host Plant Patches
2.3 Feeding Patterns of Butterfly Larvae

3. Results
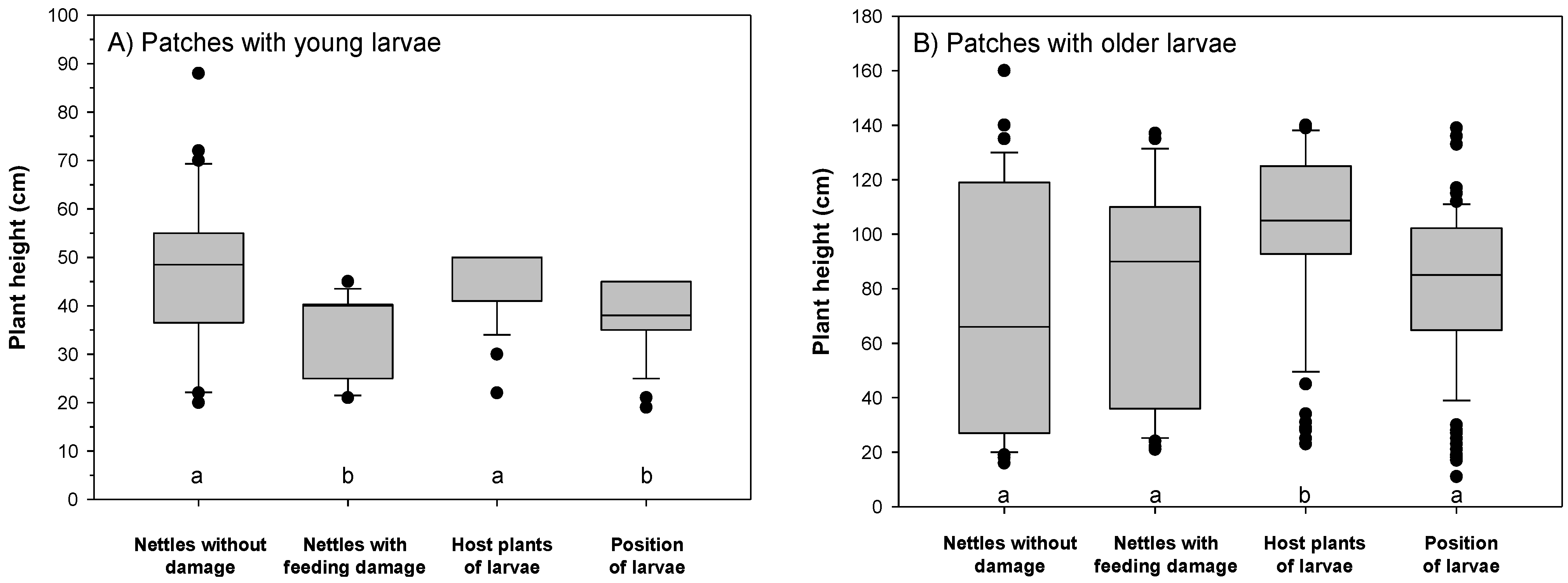
3.1. Larvae in the Host Plant Patches
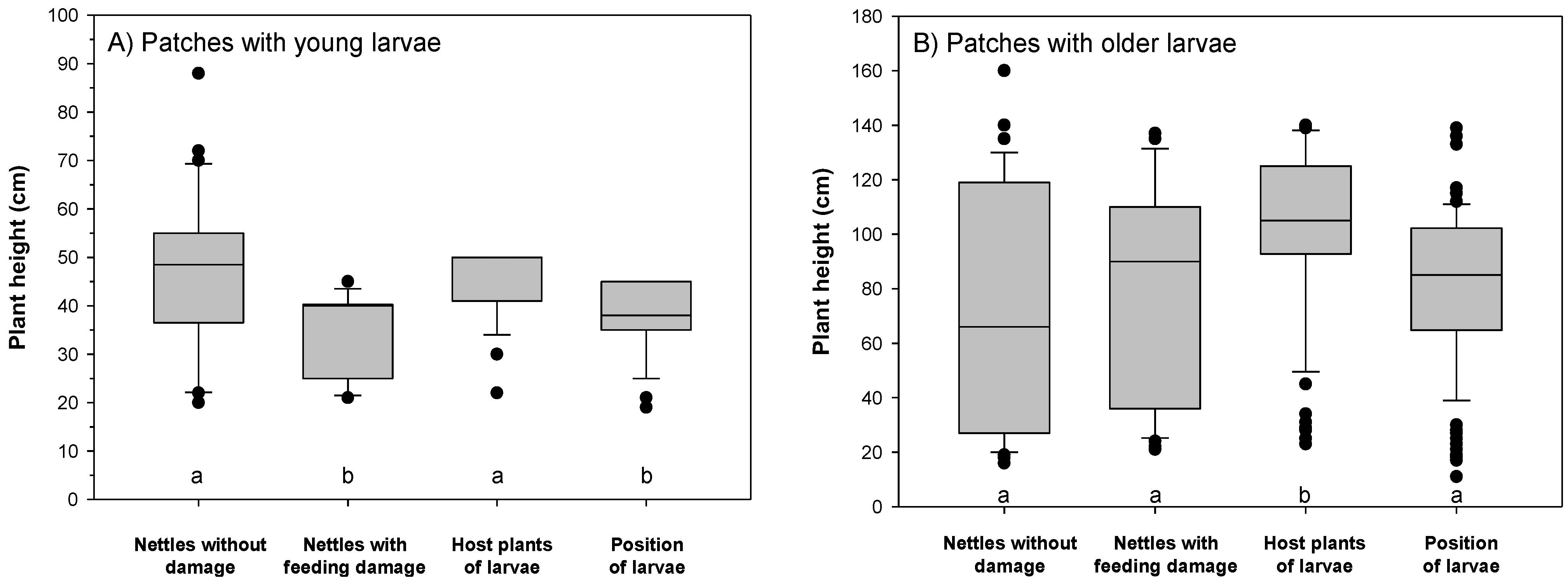
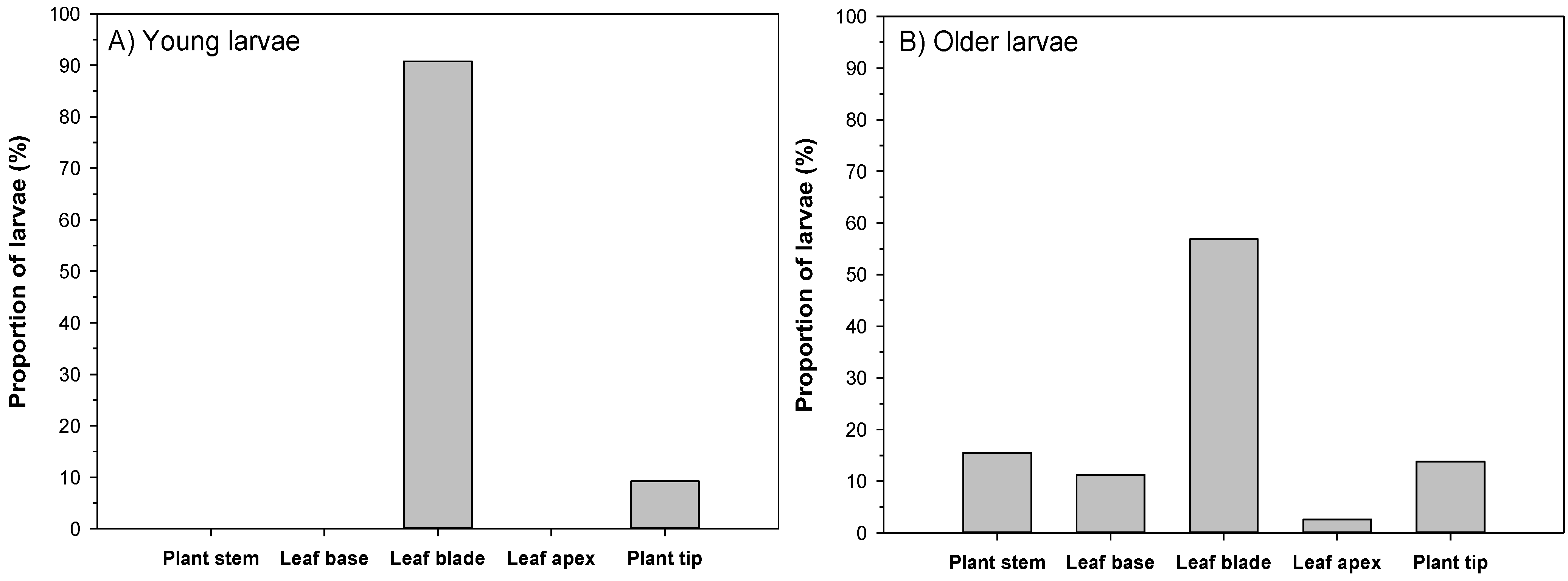
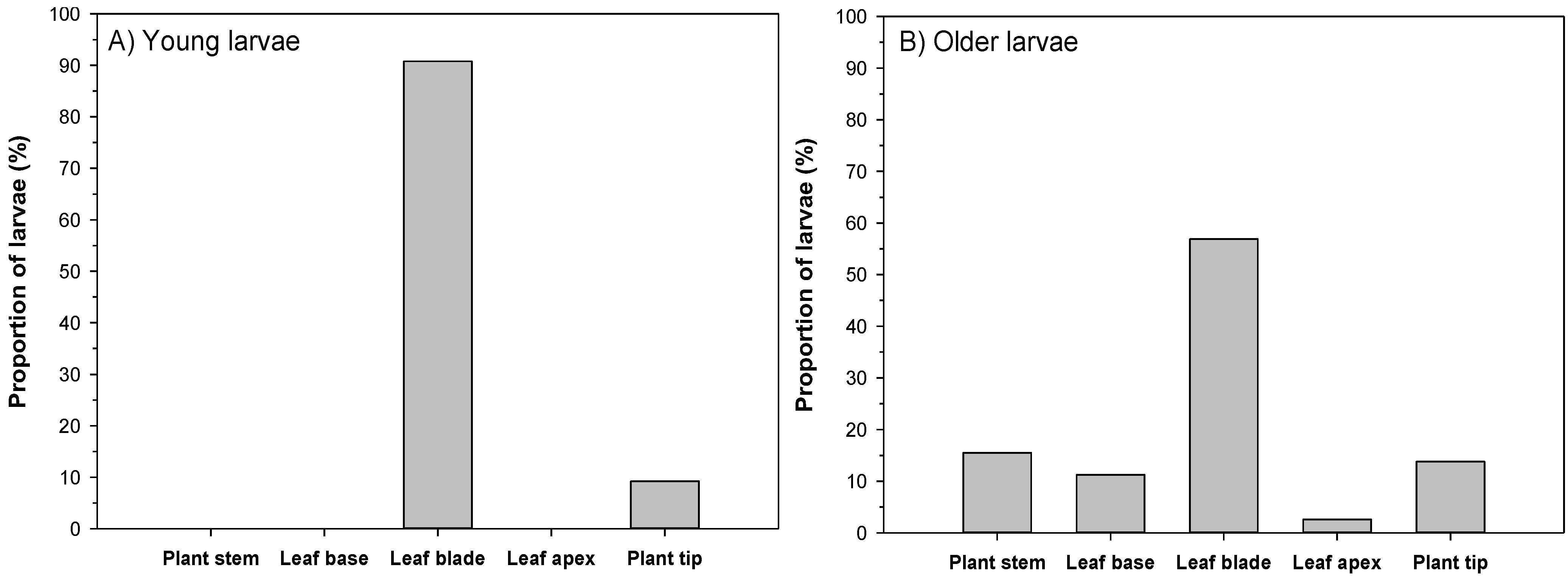
3.2. Feeding Patterns of Larvae
| Leaf Structure | Young Larvae | Older Larvae |
|---|---|---|
| Leaf base | 86% | 61% |
| Leaf blade | 92% | 87% |
| Leaf apex | 91% | 85% |
| Midrib | 17% | 26% |
| Primary vein | 68% | 82% |
| Secondary vein | 98% | 98% |
| Tertiary vein | 89% | 87% |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- EC. Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC. Off. J. Eur. Communities L 2001, 106, 1–38. [Google Scholar]
- James, C. Global Status of Commercialized Biotech/GM Crops: 2014; ISAAA Briefs 49-2014; ISAAA: Ithaca, NY, USA, 2014. [Google Scholar]
- Van Rie, J. Bacillus thuringiensis and its use in transgenic insect control technologies. Int. J. Med. Microbiol. 2000, 290, 463–469. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Jehle, J.A. Quantitative analysis of the seasonal and tissue-specific expression of Cry1Ab in transgenic maize Mon810. J. Plant Dis. Prot. 2007, 114, 82–87. [Google Scholar]
- Székács, A.; Lauber, E.; Juracsek, J.; Darvas, B. Cry1ab toxin production of MON 810 transgenic maize. Environ. Toxicol. Chem. 2010, 29, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Pleasants, J.M.; Hellmich, R.L.; Dively, G.P.; Sears, M.K.; Stanley-Horn, D.E.; Mattila, H.R.; Foster, J.E.; Clark, T.L.; Jones, G.D. Corn pollen distribution on milkweeds in and near cornfields. Proc. Natl. Acad. Sci. USA 2001, 98, 11919–11924. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, F.; Otto, M.; Kuhn, U.; Ober, S.; Schlechtriemen, U.; Vögel, R. A new method for in situ measurement of Bt-maize pollen deposition on host-plant leaves. Insects 2011, 2, 12–21. [Google Scholar] [CrossRef]
- Hofmann, F.; Otto, M.; Wosniok, W. Maize pollen deposition in relation to distance from the nearest pollen source under common cultivation—Results of 10 years of monitoring (2001 to 2010). Environ. Sci. Eur. 2014, 26. [Google Scholar] [CrossRef]
- Masetti, A.; Perry, J.N.; Dinelli, G.; Burgio, G. Phenology of Inachis io larvae and maize pollen deposition on nettles in Northern Italy field margins. IOBC-WPRS Bull. 2013, 97, 73–79. [Google Scholar]
- Losey, J.E.; Rayor, L.S.; Carter, M.E. Transgenic pollen harms monarch larvae. Available online: http://www.readcube.com/articles/10.1038%2F20338 (accessed on 5 March 2015).
- Dively, G.P.; Rose, R.; Sears, M.K.; Hellmich, R.L.; Stanley-Horn, D.E.; Calvin, D.D.; Russoe, J.M.; Anderson, P.L. Effects on monarch butterfly larvae (Lepidoptera: Danaidae) after continuous exposure to Cry1Ab-expressing corn during anthesis. Environ. Entomol. 2004, 33, 1116–1125. [Google Scholar] [CrossRef]
- Lang, A.; Vojtech, E. The effects of pollen consumption of transgenic Bt maize on the common swallowtail, Papilio machaon L. (Lepidoptera, Papilionidae). Basic Appl. Ecol. 2006, 7, 296–306. [Google Scholar] [CrossRef]
- Felke, M.; Langenbruch, G.A.; Feiertag, S.; Kassa, A. Effect of Bt-176 maize pollen on first instar larvae of the Peacock butterfly (Inachis io) (Lepidoptera; Nymphalidae). Environ. Biosaf. Res. 2010, 9, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Otto, M. A synthesis of laboratory and field studies on the effects of transgenic Bt-maize on non-target Lepidoptera. Entomol. Exp. Appl. 2010, 135, 121–134. [Google Scholar] [CrossRef]
- Sears, M.K.; Hellmich, R.L.; Stanley-Horn, D.E.; Oberhauser, K.S.; Pleasants, J.M.; Mattila, H. R Siegfried, B.D.; Dively, G.P. Impact of Bt corn pollen on monarch butterfly populations: A risk assessment. Proc. Natl. Acad. Sci. USA 2001, 98, 11937–11942. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.K.D.; Meyer, S.J.; Wolf, A.T.; Wolt, J.D.; Davis, P.M. Genetically engineered plants, endangered species, and risk: A temporal and spatial exposure assessment for Karner Blue butterfly larvae and Bt maize pollen. Risk Anal. 2006, 26, 845–858. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority). Scientific Opinion updating the evaluation of the environmental risk assessment and risk management recommendations on insect resistant genetically modified maize 1507 for cultivation. EFSA J. 2011, 2429, 1–7. [Google Scholar]
- EFSA (European Food Safety Authority). Scientific Opinion supplementing the conclusions of the environmental risk assessment and risk management recommendations for the cultivation of the genetically modified insect resistant maize Bt11 and MON 810. EFSA J. 2012, 3016, 1–98. [Google Scholar]
- Andow, D.A.; Zwahlen, C. Assessing environmental risks of transgenic plants. Ecol. Lett. 2006, 9, 196–214. [Google Scholar] [CrossRef] [PubMed]
- Andow, D.A.; Lövei, G.L.; Arpaia, S.; Wilson, L.; Fontes, E.M.G.; Hilbeck, A.; Lang, A.; van Tuat, N.; Pires, C.S.S.; Sujii, E.R.; et al. An ecologically-based method for selecting ecological indicators for assessing risks to biological diversity from genetically-engineered plants. J. Biosaf. 2014, 22, 141–156. [Google Scholar]
- Oberhauser, K.S.; Prysby, M.D.; Mattila, H.R.; Stanley-Horn, D.E.; Sears, M.K.; Dively, G.; Olson, E.; Pleasants, J.M.; Lami, W.K.F.; Hellmich, R.L. Temporal and spatial overlap between monarch larvae and corn pollen. Proc. Natl. Acad. Sci. USA 2001, 98, 11913–11918. [Google Scholar] [CrossRef] [PubMed]
- Losey, J.E.; Hufbauer, R.A.; Hartzler, R.G. Enumerating lepidopteran species associated with maize as a first step in risk assessment in the USA. Environ. Biosaf. Res. 2003, 2, 247–261. [Google Scholar] [CrossRef]
- Traxler, A.; Minarz, E.; Höttinger, H.; Pennerstorfer, J.; Schmatzberger, A.; Banko, G.; Placer, K.; Hadrobolec, M.; Gaugitsch, H. Biodiversitäts-Hotspots der Agrarlandschaft als Eckpfeiler für Risikoabschätzung und Monitoring von GVO. Available online: http://bmg.gv.at/cms/home/attachments/5/3/0/CH1050/CMS1340177559357/5_-_2005.pdf (accessed on 2 March 2015). (In German)
- Prasifka, P.L.; Hellmich, R.L.; Prasifka, J.R.; Lewis, L.C. Effects of Cry1Ab-expressing corn anthers on the movement of Monarch butterfly larvae. Environ. Entomol. 2007, 36, 228–233. [Google Scholar] [CrossRef]
- Holst, N.; Lang, A.; Lövei, G.; Otto, M. Increased mortality is predicted of Inachis io larvae caused by Bt-maize pollen in European farmland. Ecol. Model. 2013, 250, 126–133. [Google Scholar] [CrossRef]
- Zangerl, A.R.; McKenna, D.; Wraight, C.L.; Carroll, M.; Ficarello, P.; Berenbaum, M.R. Effects of exposure to event 176 Bacillus thuringiensis corn pollen on monarch and black swallowtail caterpillars under field conditions. Proc. Natl. Acad. Sci. USA 2001, 98, 11908–11912. [Google Scholar] [CrossRef] [PubMed]
- Jesse, L.C.H.; Obrycki, J.J. Field deposition of Bt transgenic corn pollen: Lethal effects on the monarch butterfly. Oecologia 2000, 125, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Gathmann, A.; Wirooks, L.; Eckert, J.; Schuphan, I. Spatial distribution of Aglais urticae (L.) in an agricultural landscape: Implications for Bt maize risk assessment and post-market monitoring. Environ. Biosaf. Res. 2006, 5, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Schuppener, M.; Mühlhauser, J.; Müller, A.-K.; Rauschen, S. Environmental risk assessment for the small tortoiseshell Aglais urticae and a stacked Bt-maize with combined resistances against Lepidoptera and Chrysomelidae in central European agrarian landscapes. Mol. Ecol. 2012, 21, 4646–4662. [Google Scholar] [CrossRef] [PubMed]
- Pollard, E.; Graetorex-Davies, J.N.; Thomas, J.A. Drought reduces breeding success of the butterfly Aglais urticae. Ecol. Entomol. 1997, 22, 315–318. [Google Scholar] [CrossRef]
- Lang, A.; Dolek, M.; Theißen, B.; Zapp, A. Are adult Crambid Snout Moths (Crambinae) and larval stages of Lepidoptera suitable tools for an environmental monitoring of transgenic crops?—Implications of a field test. Insects 2011, 2, 400–411. [Google Scholar] [CrossRef]
- Perry, J.N.; Devos, Y.; Arpaia, S.; Bartsch, D.; Gathmann, A.; Hails, R.S.; Kiss, J.; Lheureux, K.; Manachini, B.; Mestdagh, S.; et al. A mathematical model of exposure of nontarget Lepidoptera to Bt-maize pollen expressing Cry1Ab within Europe. Proc. R. Soc. B 2010, 277, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Ebert, G.; Rennwald, E. Die Schmetterlinge Baden-Württembergs, Band 1: Tagfalter I; Verlag Eugen Ulmer: Stuttgart, Deutschland, 1991. (In German) [Google Scholar]
- Kolbeck, H. Kleiner Fuchs, Aglais urticae (Linnaeus, 1758). In Tagfalter in Bayern; Bräu, M., Bolz, R., Kolbeck, H., Nummer, A., Voith, J., Wolf, W., Eds.; Verlag Eugen Ulmer: Stuttgart, Deutschland, 2013; pp. 377–378. (In German) [Google Scholar]
- Felke, M.; Langenbruch, G.A. Auswirkungen des Pollens von transgenem Bt-Mais auf ausgewählte Schmetterlingsarten. BfN Skr. 2005, 157, 1–143. (In German) [Google Scholar]
- Felke, M. Massenzucht von Aglais urticae (Linnaeus, 1758) und Inachis io (Linnaeus, 1758) (Lepidoptera, Nymphalidae) unter freilandnahen Bedingungen. Entomol. Z. 2003, 113, 211–213. (In German) [Google Scholar]
- Bryant, S.R.; Thomas, C.D.; Bale, J.S. Thermal ecology of gregarious and solitary nettle-feeding nymphalid butterfly larvae. Oecologia 2000, 122, 1–10. [Google Scholar] [CrossRef]
- Bryant, S.R.; Thomas, C.D.; Bale, J.S. The influence of thermal ecology on the distribution of three nymphalid butterflies. J. Appl. Ecol. 2002, 39, 43–55. [Google Scholar] [CrossRef]
- Pullin, A.S. Influence of food plant, Urtica dioica, on larval development, feeding efficiencies, and voltinism of a specialist insect, Inachis io. Holarct. Ecol. 1986, 9, 72–78. [Google Scholar]
- Pullin, A.S. Changes in leaf quality following clipping and regrowth of Urtica dioica, and consequences for a specialist herbivore, Aglais urticae. Oikos 1987, 49, 39–45. [Google Scholar] [CrossRef]
- Settele, J.; Steiner, R.; Reinhardt, R.; Feldmann, R.; Hermann, G. Schmetterlinge. In Die Tagfalter Deutschlands; Ulmer: Stuttgart, Deutschland, 2000. (In German) [Google Scholar]
- Lang, A.; Ludy, C.; Vojtech, E. Dispersion and distribution of Bt maize pollen in field margins. J. Plant Dis. Prot. 2004, 111, 417–428. [Google Scholar]
- Hofmann, F.; Schlechtriemen, U.; Kuhn, U.; Boessenkool, B.; Haase, S.; Niemann, E.; Schiebel, L.; Schütz, R.; Schröder, B.; Mante, C.; et al. Variabilität der Maispollendeposition auf Blättern und Exposition von Gewässerökosystemen durch Pollen und Streu aus dem Maisanbau. Fachbeiträge LUGV 2013, 133, 65–77. (In German) [Google Scholar]
- Lang, A.; Oehen, B.; Ross, J.-H.; Bieri, K.; Steinbrich, A. Environmental Geosciences, University of Basel, Switzerland. Potential exposure of butterflies in protected habitats by Bt maize cultivation: A case study in Switzerland. Biol. Conserv. 2015. submitted. [Google Scholar]
- Ohlfest, J.R.; Jesse, L.C.; Jurenka, R.; Obrycki, J.J. Stability of insecticidal CryIAb protein in transgenic Bt corn pollen exposed to UV radiation. J. Kans. Entomol. Soc. 2002, 75, 48–51. [Google Scholar]
- Stanley-Horn, D.E.; Dively, G.P.; Hellmich, R.L.; Mattila, H.R.; Sears, M.K.; Rose, R.; Jesse, L.C.H.; Losey, J.E.; Obrycki, J.J.; Lewis, L. Assessing the impact of Cry1Ab-expressing corn pollen on monarch butterfly larvae in field studies. Proc. Natl. Acad. Sci. USA 2001, 98, 11931–11936. [Google Scholar] [CrossRef] [PubMed]
- Pullin, A.S.; Bale, J.S. Effects of low temperature on diapausing Aglais urticae and Inachis io (Lepidoptera: Nympahlidae): Overwintering physiology. J. Insect Physiol. 1989, 35, 283–290. [Google Scholar] [CrossRef]
- Bryant, S.R.; Thomas, C.D.; Bale, J.S. Nettle-feeding nymphalid butterflies: Temperature, development and distribution. Ecol. Entomol. 1997, 22, 390–398. [Google Scholar] [CrossRef]
- Bryant, S.R.; Bale, J.S.; Thomas, C.D. Comparison of development and growth of nettle-feeding larvae of Nymphalidae (Lepidoptera) under constant and alternating temperature regimes. Eur. J. Entomol. 1999, 96, 143–148. [Google Scholar]
- Dennis, R.L.H.; Sparks, T.H. Landscape resources for the territorial nymphalid butterfly Inachis io: Microsite landform selection and behavioral responses to environmental conditions. J. Insect Behav. 2005, 18, 725–742. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, A.; Otto, M. Feeding Behaviour on Host Plants May Influence Potential Exposure to Bt Maize Pollen of Aglais Urticae Larvae (Lepidoptera, Nymphalidae). Insects 2015, 6, 760-771. https://doi.org/10.3390/insects6030760
Lang A, Otto M. Feeding Behaviour on Host Plants May Influence Potential Exposure to Bt Maize Pollen of Aglais Urticae Larvae (Lepidoptera, Nymphalidae). Insects. 2015; 6(3):760-771. https://doi.org/10.3390/insects6030760
Chicago/Turabian StyleLang, Andreas, and Mathias Otto. 2015. "Feeding Behaviour on Host Plants May Influence Potential Exposure to Bt Maize Pollen of Aglais Urticae Larvae (Lepidoptera, Nymphalidae)" Insects 6, no. 3: 760-771. https://doi.org/10.3390/insects6030760
APA StyleLang, A., & Otto, M. (2015). Feeding Behaviour on Host Plants May Influence Potential Exposure to Bt Maize Pollen of Aglais Urticae Larvae (Lepidoptera, Nymphalidae). Insects, 6(3), 760-771. https://doi.org/10.3390/insects6030760