Insects as a Nitrogen Source for Plants


{kind=link}
Abstract
1. Introduction
2. Nitrogen in the Soil—Why Is It a Deficit?
3. Insects as a Nitrogen Reservoir
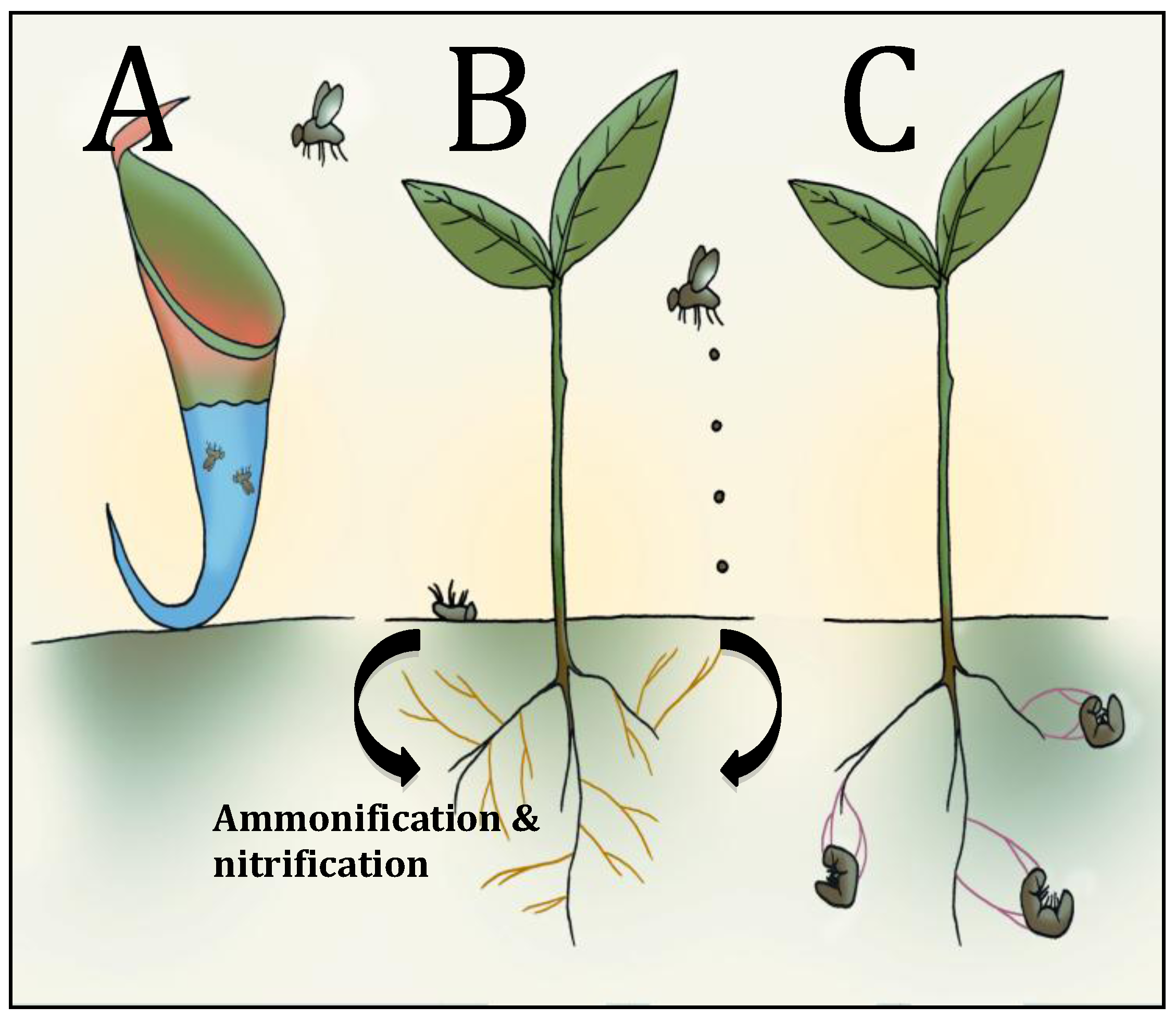
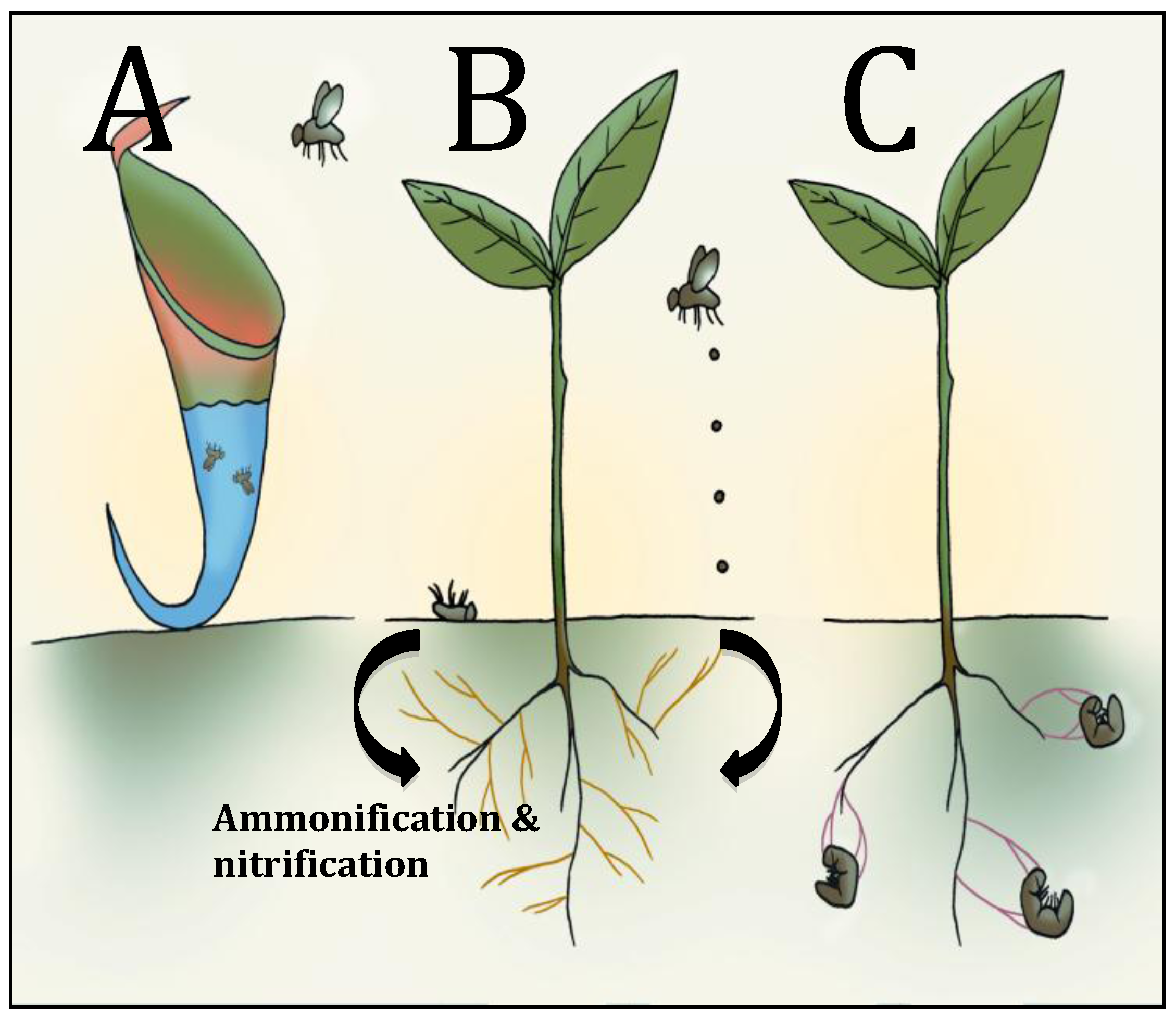
4. Carnivorous Plants
5. Insect Cadavers and Insect Frass: Potential Involvement of Mycorrhizal Fungi
5.1. Insect Cadavers/Insect Frass
5.2. Potential Role of Myccorhizal Fungi in Insect Nitrogen Transfer
6. Endophytic Insect Pathogenic Fungi
7. Evolutionary Implication of Nitrogen Transfer by Fungal Symbionts
8. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Schulten, H.R.; Schnitzer, M. The chemistry of soil organic nitrogen: A review. Biol. Fertil. Soils 1998, 26, 1–15. [Google Scholar] [CrossRef]
- Cocking, E.C. Endophytic colonization of plant roots by nitrogen-fixing bacteria. Plant Soil 2003, 252, 169–175. [Google Scholar] [CrossRef]
- Mary, B.; Recous, S.; Darwis, D.; Robin, D. Interactions between decomposition of plant residues and nitrogen cycling in soil. Plant Soil 1996, 181, 71–82. [Google Scholar] [CrossRef]
- Behie, S.W.; Zelisko, P.M.; Bidochka, M.J. Endophytic insect-parasitic fungi translocate nitrogen directly from insects to plants. Science 2012, 336, 1576–1577. [Google Scholar] [CrossRef]
- Ellison, A.M.; Gotelli, N.J. Evolutionary ecology of carnivorous plants. Trends Ecol. Evol. 2001, 16, 623–629. [Google Scholar] [CrossRef]
- Sprent, J.I. Benefits of Rhizobium to agriculture. Trends Biotechnol. 1986, 4, 124–129. [Google Scholar] [CrossRef]
- Johansson, J.F.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 2006, 48, 1–13. [Google Scholar] [CrossRef]
- Koller, R.; Rodriguez, A.; Robin, C.; Scheu, S.; Bonkowski, M. Protozoa enhance foraging efficiency of arbuscular mycorrhizal fungi for mineral nitrogen from organic matter in soil to the benefit of host plants. New Phytol. 2013, 199, 203–211. [Google Scholar] [CrossRef]
- Liaw, Y.P. Comparison of field, laboratory, and theoretical estimates of global nitrogen fixation by lightning. J. Geophys. Res. 1990, 95, 22489–22494. [Google Scholar] [CrossRef]
- Blossey, B.; Hunt-Joshi, T.R. Belowground herbivory by insects: Influence on plants and aboveground herbivores. Annu. Rev. Entomol. 2003, 48, 521–547. [Google Scholar] [CrossRef]
- Post, W.M.; Pastor, J.; Zinke, P.J.; Stangenberger, A.G. Global patterns of soil nitrogen storage. Nature 1985, 317, 613–616. [Google Scholar] [CrossRef]
- Parsons, L.L.; Smith, M.S.; Murray, R.E. Soil denitrification dynamics: Spatial and temporal variations of enzyme activity, populations, and nitrogen gas loss. Soil Sci. Soc. Am. J. 1991, 55, 90–95. [Google Scholar] [CrossRef]
- Goulding, K.W.Y.; Bailey, N.J.; Bradbury, N.J.; Hargreaves, P.; Howe, M.; Murphy, D.V.; Poulton, P.R.; Willison, T.W. Nitrogen deposition and its contribution to nitrogen cycling and associated soil processes. New Phytol. 1998, 139, 49–58. [Google Scholar]
- Whittaker, J.B. Insects and plants in a changing atmosphere. J. Ecol. 2001, 89, 507–518. [Google Scholar] [CrossRef]
- Wardle, D.A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen in the soil. Biol. Rev. 1992, 67, 321–358. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; SeabloomE, W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phos-phorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef]
- Gutschick, V.P. Evolved strategies in nitrogen acquisition by plants. Am. Nat. 1981, 118, 607–637. [Google Scholar]
- Miller, H.G. Forest fertilization: some guiding concepts. Forestry 1981, 54, 157–167. [Google Scholar] [CrossRef]
- Agren, G.I. Nitrogen productivity of some conifers. Can. J. For. Res. 1983, 13, 494–500. [Google Scholar] [CrossRef]
- Huenneke, L.F.; Hamburg, S.P.; Koide, R.; Mooney, H.A.; Vitousek, P.M. Effects of soil resources on plant invasion and community structure in Californian serpentine grassland. Ecology 1990, 71, 478–491. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F.S. Response to fertilization by various plant growth forms in an Alaskan tundra: Nutrient accumulation and growth. Ecology 1980, 61, 662–675. [Google Scholar] [CrossRef]
- Field, C.; Mooney, H.A. The photosynthesis-nitrogen relationship in wild plants. In On the Economy of Plant Form and Function; Givnish, T.J., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 25–55. [Google Scholar]
- Swank, W.T.; Waide, J.B.; Crossley, D.A.; Todd, R.L. Insect defoliation enhances nitrate export from forested ecosystems. Oecologia 1981, 51, 297–299. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Melilo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and nitrogen dynamics long the decay continuum: Plant litter to soil organic matter. Dev. Plant Soil Sci. 1989, 39, 53–62. [Google Scholar]
- Kelley, K.R.; Stevenson, F.J. Forms and nature of organic N in soil. Fert. Res. 1995, 42, 1–11. [Google Scholar] [CrossRef]
- Ayers, E.; Dromph, K.M.; Cook, R.; Ostle, N.; Bardgett, R.D. The influence of below-ground herbivory and defoliation of a legume on nitrogen transfer to neighbouring plants. Funct. Ecol. 2007, 21, 256–263. [Google Scholar] [CrossRef]
- Mattson, W.J. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar]
- Maron, J.L. Insect herbivory above and belowground: Individual and joint effects on plant fitness. Ecology 1998, 79, 1281–1293. [Google Scholar] [CrossRef]
- Murray, P.J.; Hatch, J.B.; Cliquet, J.B. Impact of insect root herbivory on the growth and nitrogen and carbon contents of white clover (Trifolium repens) seedlings. Can. J. Bot. 1996, 74, 1591–1595. [Google Scholar] [CrossRef]
- Fagan, W.F.; Siemann, E.; Mitter, C.; Denno, R.F.; Huberty, A.F.; Woods, A.; Elser, J.J. Nitrogen in Insects: Implications for Trophic Complexity and Species Diversification. Am. Nat. 2002, 160, 784–802. [Google Scholar]
- Van Emden, H.F. Pest Control, 2nd ed. ed.; Edward Arnold Publication: London, UK, New York, NY, USA, 1989. [Google Scholar]
- Kummerow, J.; Alexander, J.V.; Neel, J.W.; Fishbeck, K. Symbiotic nitrogen fixation in Ceanothus roots. Am. J. Bot. 1978, 65, 63–69. [Google Scholar] [CrossRef]
- Brewer, J.S. A demographic analysis of fire-stimulated seedling establishment of Sarracenia alata (Sarraceniacae). Am. J. Bot. 2001, 88, 1250–1257. [Google Scholar] [CrossRef]
- Müller, K.F.; Borsch, T.; Legendre, L.; Porembski, S.; Barthlott, W. Recent progress in understanding the evolution of carnivorous Lentibulariaceae (Lamiales). Plant Biol. 2006, 8, 748–757. [Google Scholar] [CrossRef]
- Moran, J.A.; Clarke, C.; Gowen, B.E. The use of light in prey capture by the tropical pitcher plant Nepenthes aristolochioides. Plant Signal. Behav. 2012, 7, 957–960. [Google Scholar] [CrossRef]
- Jaffi, K.; Blum, M.S.; Fales, H.M.; Mason, R.T.; Cabrera, A. Insect attractants from pitcher plants of the genus Heliamphora (Sarraceniacae). J. Chem. Ecol. 1995, 21, 379–384. [Google Scholar] [CrossRef]
- Adamec, L. Mineral nutrition of carnivorous plants: A review. Bot. J. 1997, 63, 273–299. [Google Scholar]
- Ellison, A.M.; Gotelli, N.J. Nitrogen availability alters the expression of carnivory in the northern pitcher plant, Sarraceniapurpurea. Proc. Natl. Acad. Sci. USA 2002, 99, 4409–4412. [Google Scholar] [CrossRef]
- Volkov, A.G.; Adesina, T.; Markin, V.S.; Jovanov, E. Kinetics and mechanism of Dionaea muscipula trap closing. Plant Physiol. 2008, 146, 694–702. [Google Scholar]
- Volkov, A.G.; Carrell, H.; Baldwin, A.; Markin, V.S. Electrical memory in Venus flytrap. Bioelectrochemistry 2009, 75, 142–147. [Google Scholar] [CrossRef]
- Jaffe, K.; Michelangeli, F.; Gonzalez, J.M.; Miras, B.; Ruiz, M.C. Carnivory in Pitcher Plants of the Genus Heliamphora (Sarraceniaceae). New Phytol. 1992, 122, 733–744. [Google Scholar]
- Gaume, L.; Forterre, Y. A viscoelastic deadly fluid in carnivorous pitcher plants. PLoS One 2007, 2, e1185. [Google Scholar] [CrossRef]
- Joel, D.M. Mimicry and mutualism in carnivorous pitcher plants (Sarraceniaceae, Nepenthaceae, Cephalotaceae, Bromeliaceae). Biol. J. Linn. Soc. 1988, 35, 185–197. [Google Scholar] [CrossRef]
- Fielding, D.J.; Trainor, E.; Zhang, M. Diet influences rates of carbon and nitrogen mineralization from decomposing grasshopper frass and cadavers. Biol. Fertil. Soils 2012. [Google Scholar] [CrossRef]
- Hunter, M.D. Insect population dynamics meets ecosystem ecology: Effects of herbivory on soil nutrient dynamics. Agr. For. Entomol. 2001, 3, 77–84. [Google Scholar] [CrossRef]
- Frost, C.J.; Hunter, M.D. Recycling of nitrogen in herbivore feces: Plant recovery, herbivore assimilation, soil retention, and leaching losses. Oecologia 2007, 151, 42–53. [Google Scholar] [CrossRef]
- Zaady, E.; Groffman, M.; Shachak, M.; Wilby, A. Consumption and release ofnitrogen by the harvester termite Anacanthotermes ubachi navas in the northern Negev desert, Israel. Soil Biol. Biochem. 2003, 35, 1299–1303. [Google Scholar] [CrossRef]
- Lovett, G.M.; Ruesink, A.E. Carbon and nitrogen mineralization from decomposing gypsy moth frass. Oecologia 1995, 104, 133–138. [Google Scholar] [CrossRef]
- Hollinger, D.Y. Herbivory and the cycling of nitrogen and phosphorus in isolated California oak trees. Oecologia 1986, 70, 291–297. [Google Scholar] [CrossRef]
- Antonie, B.J. Diversity of ammonifying bacteria. Hydrobiologia 1981, 83, 343–350. [Google Scholar] [CrossRef]
- Sánchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef]
- George, E.; Marschner, H.; Jakobsen, I. Role of arbuscular mycorrhizal fungi in uptake of phosphorus and nitrogen from soil. Crit. Rev. Biotechnol. 1995, 15, 257–270. [Google Scholar] [CrossRef]
- Marschner, H.; Dell, B. Nutrient uptake in mycorrhizal symbiosis. Plant Soil 1994, 159, 89–102. [Google Scholar]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bucking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal symbiosis, 2nd ed. ed.; Academic Press: London, UK, 1997. [Google Scholar]
- Klironomos, J.N.; Hart, M.M. Food-web dynamics: Animal nitrogen swap for plant carbon. Nature 2001, 410, 651–652. [Google Scholar] [CrossRef]
- Talbot, J.M.; Allison, S.D.; Treseder, K.K. Decomposers in disguise: Mycorrhizal fungi as regulators of soil C dynamics in ecosystems under global change. Funct. Ecol. 2008, 22, 955–963. [Google Scholar] [CrossRef]
- Harrison, M.J. Biotrophic interfaces and nutrient transport in plant/fungal symbioses. J. Exp. Bot. 1999, 63, 1013–1022. [Google Scholar]
- Gaude, N.; Bortfeld, S.; Duensing, N.; Lohse, M.; Franziska, K. Arbuscule-containing and non-colonized cortical cells of mycorrhizal roots undergo a massive and specific reprogramming during arbuscular mycorrhizal development. Plant J. 2012, 69, 510–528. [Google Scholar] [CrossRef]
- Kobae, Y.; Tamura, Y.; Takai, S.; Banba, M.; Hata, S. Localized expression of arbus-cular mycorrhiza-inducible ammonium trans-porters in soybean. Plant Cell Physiol. 2010, 51, 1411–1415. [Google Scholar] [CrossRef]
- Wulf, A.; Manthey, K.; Doll, J.; Perlick, A.M.; Linke, B.; Bekel, T.; Folker, M.; Franken, P.; Kuster, H.; Kranjinski, F. Transcriptional changes in response to arbuscular mycorrhiza development in the model plant Medicago truncatula. Am. Phytopath. Soc. 2003, 16, 306–314. [Google Scholar]
- Benedetto, A.; Magurno, F.; Bonfante, P.; Lanfranco, L. Expression profiles of a phosphate transporter gene (GmosPT) from the endomycorrhizal fungus Glomus mosseae. Mycorrhiza 2005, 15, 620–627. [Google Scholar] [CrossRef]
- Harrison, M.J. A sugar transporter from Medicago truncatula: Altered expression pat-terns in roots during vesicular-arbuscular (VA) mycorrhizal associations. Plant J. 1996, 9, 491–503. [Google Scholar]
- Pearson, J.N.; Jakobsen, I. Symbiotic exchange of carbon and phosphorus between cucumber and three arbuscular mycorrhizal fungi. New Phytol. 1993, 124, 481–488. [Google Scholar] [CrossRef]
- Hobbie, J.E.; Hobbie, E.A. 15N in symbiotic fungi and plants estimates nitrogen and carbon flux rates in arctic tundra. Ecology 2006, 87, 816–822. [Google Scholar] [CrossRef]
- Newsham, K.K. A meta-analysis of plant responses to dark septate root endophytes. New Phytol. 2011, 190, 783–193. [Google Scholar] [CrossRef]
- Usuki, F.; Narisawa, K. A mutualistic symbiosis between a dark septate endophytic fungus, Heteroconium chae-tospira, and a nonmycorrhizal plant, Chinese cabbage. Mycologia 2007, 99, 175–184. [Google Scholar] [CrossRef]
- Hashiba, T.; Narisawa, K. The development and endophytic nature of the fungus Heteroconium chaetospira. FEMS Microbiol. 2005, 252, 191–196. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef]
- Sasan, R.K.; Bidochka, M.J. The insect-pathogenic fungus Metarhizium robertsii (Claviciptaceae) is alos in endophyte that stimulates plant root development. Am. J. Bot. 2012, 99, 101–107. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Gold, C.S.; Coyne, D.; Nakavuma, J.; Paparu, P. Beauveria bassiana (Balsamo) Vuillemin as an endophyte in tissue culture banana (Musa spp.). J. Invertebr. Pathol. 2007, 96, 34–42. [Google Scholar] [CrossRef]
- Roberts, D.W.; Hajek, A.E. Entomopathogenic fungi as bioinsecticides. In Frontiers of Industrial Mycology; Leatham, G.F., Ed.; Chapman and Hall: New York, NY, USA, 1992; pp. 114–159. [Google Scholar]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? In Ecology; 2007; Volume 88, pp. 541–549. [Google Scholar]
- Porras-Alfaro, A.; Herrera, J.; Sinsabaugh, R.L.; Odenbach, K.J.; Lowrey, T.; Natvig, D.O. Novel root fungal consortium associated with a dominant desert grass. Appl. Environ. Microbiol. 2008, 74, 2805–2813. [Google Scholar] [CrossRef]
- Behie, S.W.; Padilla-Guerrero, I.E.; Bidochka, M.J. Nutrient transfer to plants by phylogenetically diverse fungi suggests convergent evolutionary strategies in rhizospheric symbionts. Commun. Integr. Biol. 2013, 6, 1–4. [Google Scholar]
- Plummer, G.L.; Kethley, J.B. Foliar absorption of amino acids, peptides, and other nutrients by the pitcher plant, Sarracenia flava. Bot. Gaz. 1964, 125, 245–260. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Behie, S.W.; Bidochka, M.J. Insects as a Nitrogen Source for Plants. Insects 2013, 4, 413-424. https://doi.org/10.3390/insects4030413
Behie SW, Bidochka MJ. Insects as a Nitrogen Source for Plants. Insects. 2013; 4(3):413-424. https://doi.org/10.3390/insects4030413
Chicago/Turabian StyleBehie, Scott W., and Michael J. Bidochka. 2013. "Insects as a Nitrogen Source for Plants" Insects 4, no. 3: 413-424. https://doi.org/10.3390/insects4030413
APA StyleBehie, S. W., & Bidochka, M. J. (2013). Insects as a Nitrogen Source for Plants. Insects, 4(3), 413-424. https://doi.org/10.3390/insects4030413