Environmental Engineering Approaches toward Sustainable Management of Spider Mites

Abstract
:1. Introduction
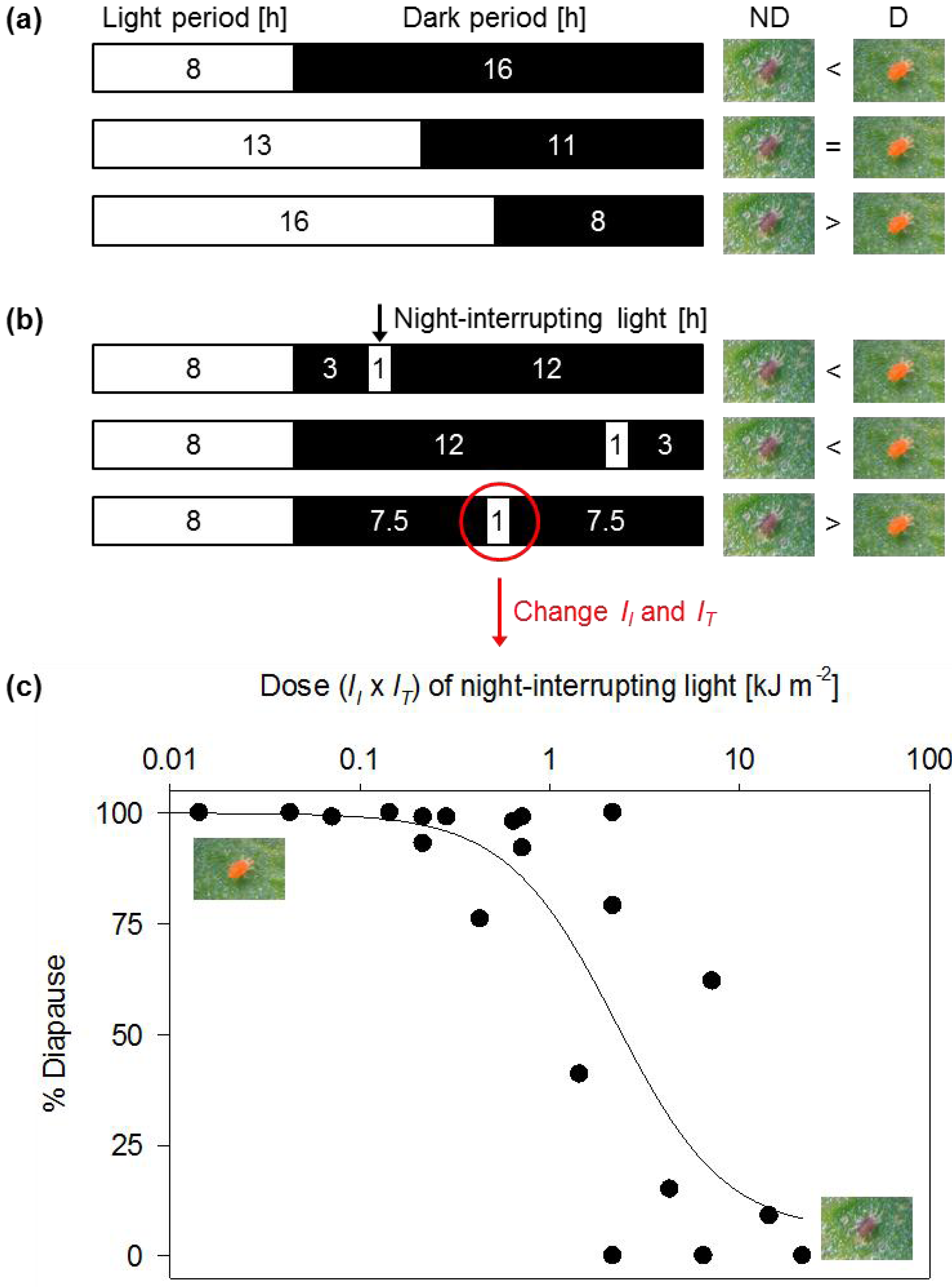
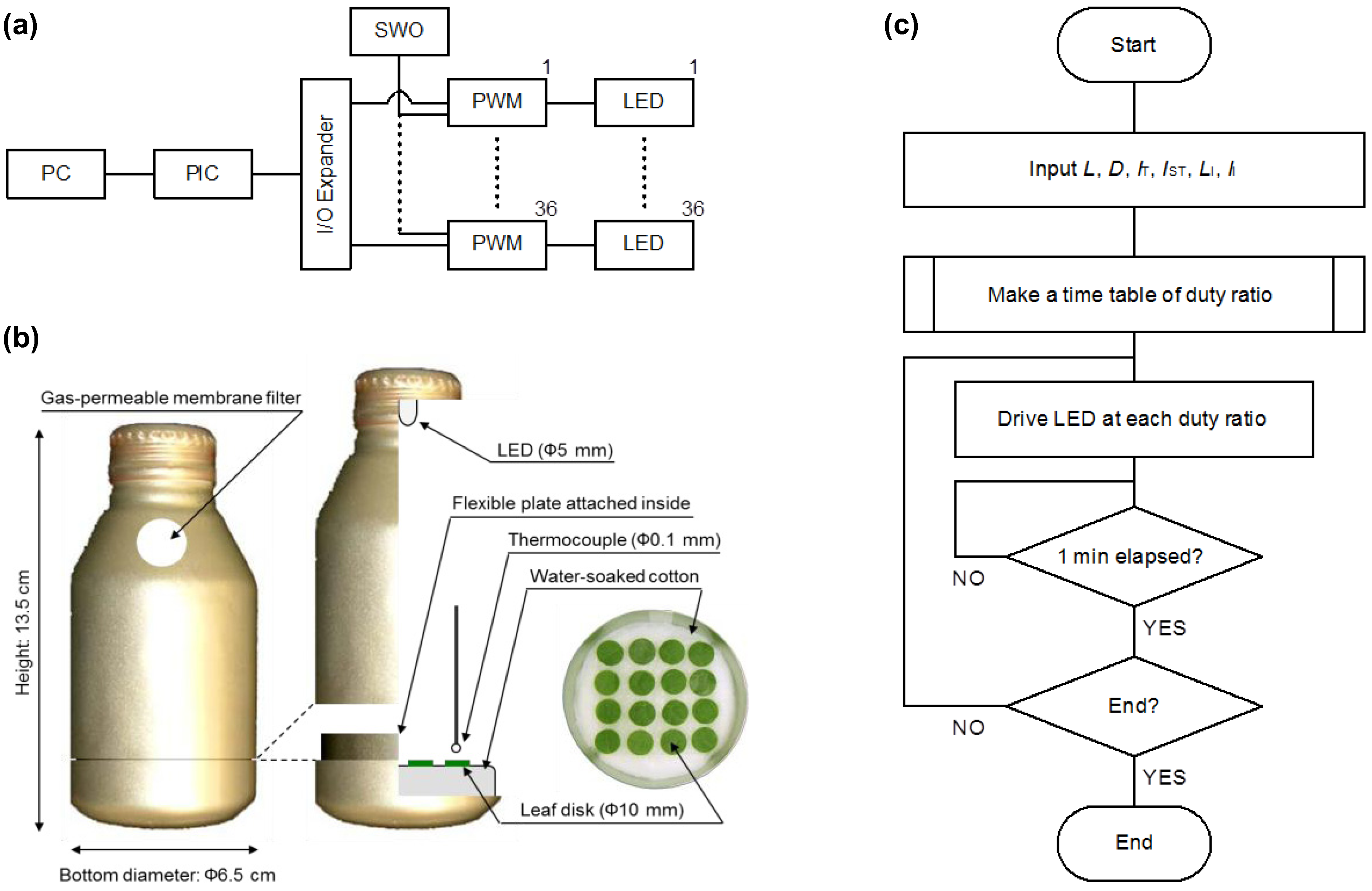
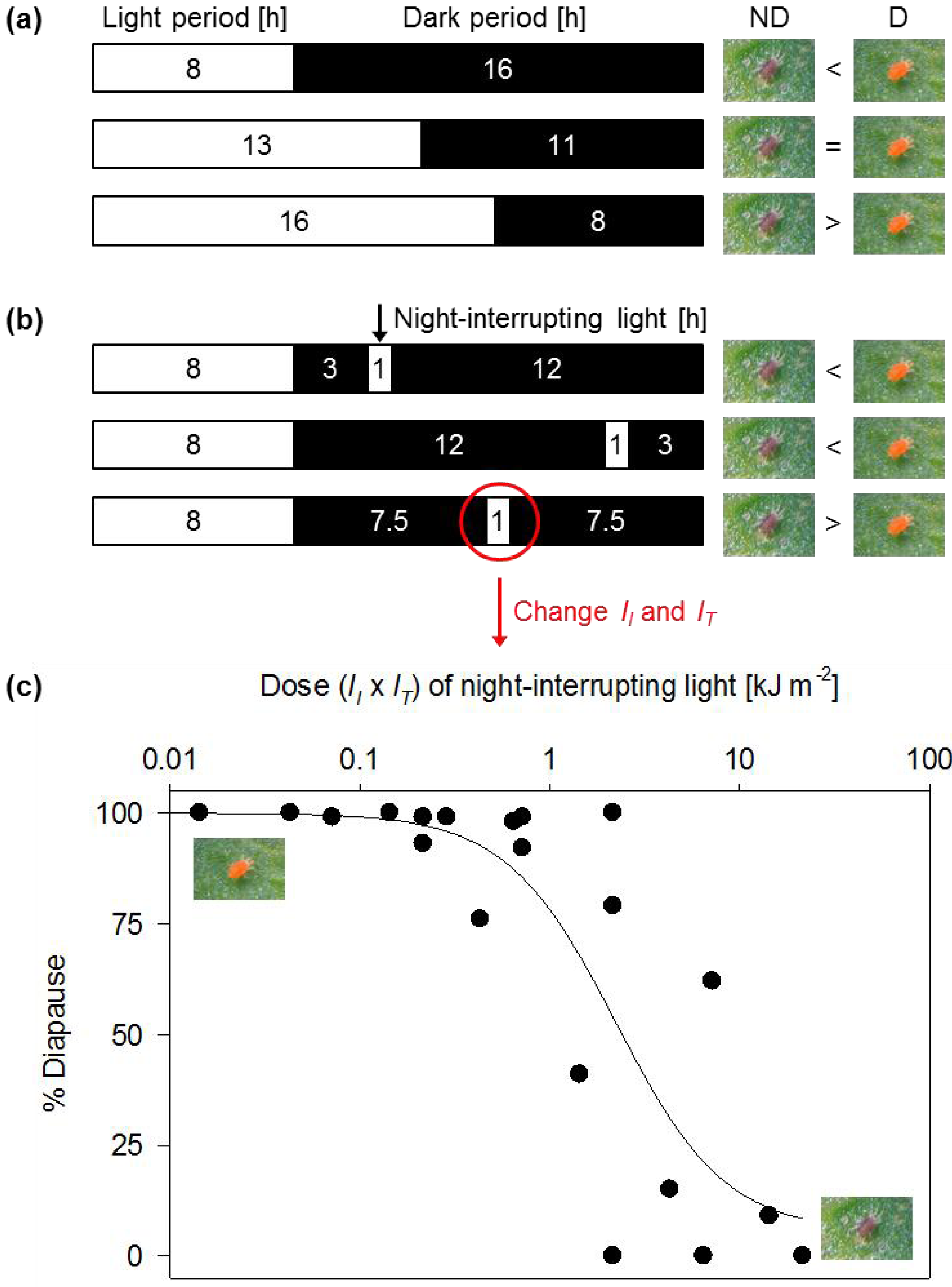
2. Photoperiod Control

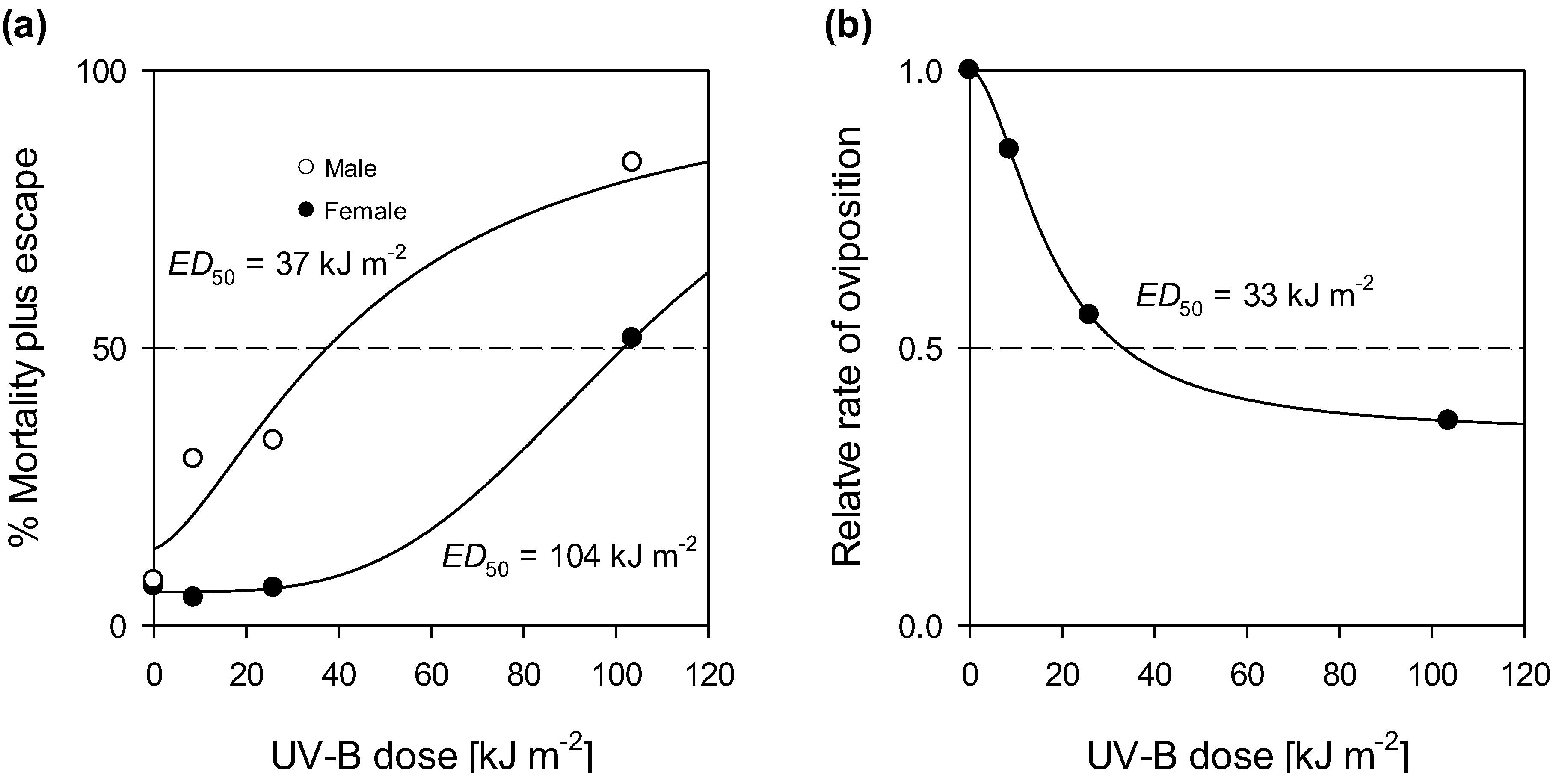
3. UV Control
4. Water Vapor Control

{kind=link}
{kind=link}
{kind=link}
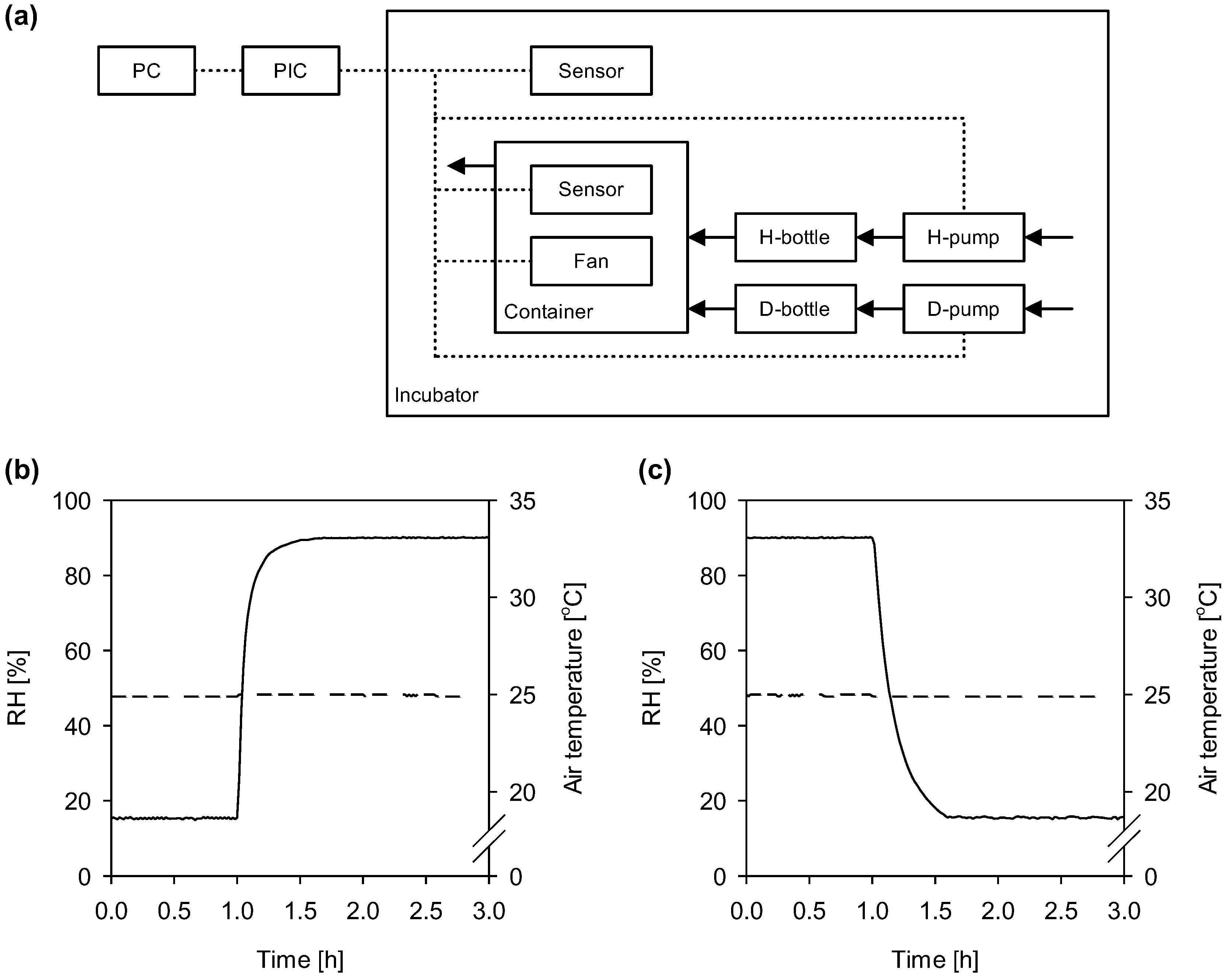
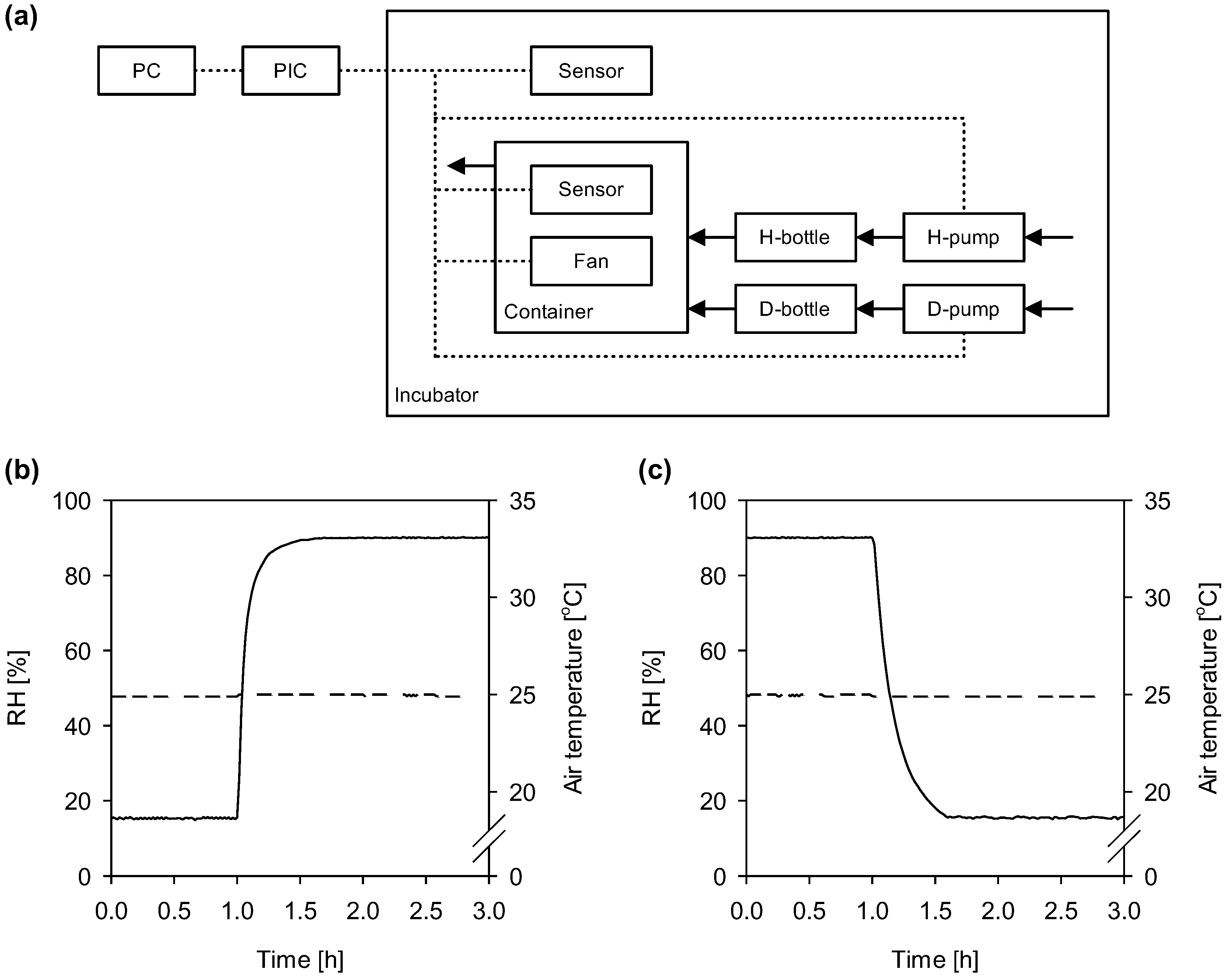{kind=link}
| Species | Stage (♀/♂) | Air temperature (°C) | RH (%) | VPD (kPa) | Food or chemical | Storage (d) | Survival (%) | Ref. |
|---|---|---|---|---|---|---|---|---|
| P. persimilis | Adult (♀) | 10 | 100 | 0.0 | T. kanzawai eggs | 70 | 93 | [96] |
| P. persimilis | Adult (♀) | 7.5 | 100 * | 0.0 * | T. urticae eggs | 56 | 43 | [91] |
| P. persimilis | Adult (♀) | 10 | 100 | 0.0 | – | 50 | 60 | [96] |
| P. persimilis | Adult (♀) | 8 | 70 | 0.3 | – | 35 | 51 | [97] |
| P. persimilis | Adult (♀) | 8 | 70–90 | 0.1–0.3 | Cryoprotectant | 29 | 46 | [98] |
| P. persimilis | Adult (♀) | 8 | 100 | 0.0 | – | 14 | 80 | [99] |
| P. persimilis | Egg | 10 | 100 | 0.0 | – | 25 | 98 | [96] |
| N. californicus | Adult (♀) | 5 | 100 | 0.0 | – | 75 | 35 | [93] |
| N. californicus | Adult (♀) | 5 | 100 | 0.0 | – | 65 | 50 | [92] |
| N. californicus | Adult (♀) | 10 | 92 | 0.1 | – | 30 | 83 | [85] |
| N. californicus | Adult (♂) | 5 | 100 | 0.0 | – | 32 | 50 | [100] |
| A. cucumeris | 9 | 100* | 0.0* | – | 70 | 63 | [101] | |
5. Conclusions
Acknowledgements
References
- March, R.B. Properties and actions of bridged diphenyl acaricides. Environ. Health Perspect. 1976, 14, 83–91. [Google Scholar] [CrossRef]
- Chant, D.C. Integrated control systems. In Scientific Aspects of Pest Control; Natural Academy Scientific Publications: Washington, DC, USA, 1966; pp. 193–218. [Google Scholar]
- Bartlett, B.R. Outbreaks of two-spotted spider mites and cotton aphids following pesticide treatment. I. Pest stimulation vs. natural enemy destruction as the cause of outbreaks. J. Econ. Entomol. 1968, 61, 297–303. [Google Scholar]
- Dittrich, V.; Streibert, P.S.; Bathe, P.A. An old case reopened: mite stimulation by insecticide residues. Environ. Entomol. 1974, 3, 534–540. [Google Scholar]
- McMurtry, J.A.; Huffaker, C.B.; van de Vrie, M. Ecology of tetranychid mites and their natural enemies: A review. I. Tetranychid enemies: Their biological characters and the impact of spray practices. Hilgardia. 1970, 40, 331–390. [Google Scholar]
- Huffaker, C.B.; van de Vrie, M.; McMurtry, J.A. Ecology of Tetranychid mites and their natural enemies: A review. II. Tetranychid populations and their possible control by predators: An evaluation. Hilgardia. 1970, 40, 391–458. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Integrated Pest Management. Available online: http://www.fao.org/agriculture/crops/core-themes/theme/pests/ipm/en (accessed on 15 October 2012).
- Lewis, W.J.; van Lenteren, J.C.; Phatak, S.C.; Tumlinson, J.H. A total system approach to sustainable pest management. Proc. Natl. Acad. Sci. USA 1997, 94, 12243–12248. [Google Scholar] [CrossRef]
- Vänninen, I.; Pinto, D.M.; Nissinen, A.I.; Johansen, N.S.; Shipp, L. In the light of new greenhouse technologies: 1. Plant mediated effects of artificial lighting on arthropods and tritrophic interactions. Ann. Appl. Biol. 2010, 157, 393–414. [Google Scholar] [CrossRef]
- Johansen, N.S.; Vänninen, I.; Pinto, D.M.; Nissinen, A.I.; Shipp, L. In the light of new greenhouse technologies: 2. Direct effects of artificial lighting on arthropods and integrated pest management in greenhouse crops. Ann. Appl. Biol. 2011, 159, 1–27. [Google Scholar] [CrossRef]
- Herms, W.B. A field test of the effect of artificial light on the behavior of the codling moth Carpocapsa. pomonella Linn. J. Econ. Entomol. 1929, 22, 78–88. [Google Scholar]
- Nomura, K.; Oya, S.; Watanabe, I.; Kawamura, H. Studies on orchard illumination against fruit-piercing moths. I. Analysis of illumination effects, and influence of light elements on moths' activities. Jpn. J. Appl. Entomol. Zool. 1965, 9, 179–186. [Google Scholar] [CrossRef]
- Nemec, S.J. Use of artificial lighting to reduce Heliothis spp. populations in cotton fields. J. Econ. Entomol. 1969, 62, 1138–1140. [Google Scholar]
- Yeh, N.; Chung, J.P. High-brightness LEDs—Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sust. Energy Rev. 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Watanabe, H. Light-controlled plant cultivation system in Japan—development of a vegetable factory using LEDs as a light source for plants. Acta. Hort. 2011, 907, 37–44. [Google Scholar]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: Oxford, UK, 1986; p. 411. [Google Scholar]
- Burkett, D.A.; Butler, J.F.; Kline, D.L. Field evaluation of colored light-emitting diodes as attractants for woodland mosquitoes and other Diptera in north central Florida. J. Am. Mosq. Control. Assoc. 1998, 14, 186–195. [Google Scholar]
- Chu, C.C.; Jackson, C.G.; Alexander, P.J.; Karut, K.; Henneberry, T.J. Plastic cup traps equipped with light-emitting diodes for monitoring adult Bemisia. tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 2003, 96, 543–546. [Google Scholar] [CrossRef]
- Chen, T.Y.; Chu, C.C.; Fitzgerald, G.; Natwick, E.T.; Henneberry, T.J. Trap evaluation for thrips (Thysanoptera: Thripidae) and hoverflies (Diptera: Syrphidae). Environ. Entomol. 2004, 33, 1416–1420. [Google Scholar] [CrossRef]
- Nakamoto, Y.; Kuba, H. The effectiveness of a green light emitting diode (LED) trap at capturing the West Indian sweet potato weevil, Euscepes. postfasciatus (Fairmaire) (Coleoptera: Curculionidae) in a sweet potato field. Appl. Entomol. Zool. 2004, 39, 491–495. [Google Scholar] [CrossRef]
- Bishop, A.L.; Bellis, G.A.; McKenzie, H.J.; Spohr, L.J.; Worrall, R.J.; Harris, A.M.; Melville, L. Light trapping of biting midges Culicoides. spp. (Diptera: Ceratopogonidae) with green light-emitting diodes. Aust. J. Entomol. 2006, 45, 202–205. [Google Scholar] [CrossRef]
- Hoel, D.F.; Butler, J.F.; Fawaz, E.Y.; Watany, N.; El-Hossary, S.S.; Villinski, J. Response of phlebotomine sand flies to light-emitting diode-modified light traps in southern Egypt. J. Vector Ecol. 2007, 32, 302–308. [Google Scholar] [CrossRef]
- Mann, R.S.; Kaufman, P.E.; Butler, J.F. Lutzomyia. spp. (Diptera: Psychodidae) response to olfactory attractant- and light emitting diode-modified Mosquito Magnet X (MM-X) traps. J. Med. Entomol. 2009, 46, 1052–1061. [Google Scholar] [CrossRef]
- Duehl, A.J.; Cohnstaedt, L.W.; Arbogast, R.T.; Teal, P.E.A. Evaluating light attraction to increase trap efficiency for Tribolium. castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2011, 104, 1430–1435. [Google Scholar] [CrossRef]
- Oh, M.S.; Lee, C.H.; Lee, S.G.; Lee, H.S. Evaluation of high power light emitting diodes (HPLEDs) as potential attractants for adult Spodoptera. exigua (Hübner) (Lepidoptera: Noctuidae). J. Kor. Soc. Appl. Biol. Chem. 2011, 54, 416–422. [Google Scholar] [CrossRef]
- Jeon, J.H.; Oh, M.S.; Cho, K.S.; Lee, H.S. Phototactic response of the rice weevil, Sitophilus. oryzae Linnaeus (Coleoptera: Curculionidae), to light-emitting diodes. J. Kor. Soc. Appl. Biol. Chem. 2012, 55, 35–39. [Google Scholar] [CrossRef]
- Greathead, D.J. Benefits and risks of classical biological control. In Biological Control: Benefits and Risks; Hokkanen, H.M.T., Lynch, J.M., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 53–63. [Google Scholar]
- van Lenteren, J.C.; Babendreier, D.; Bigler, F.; Burgio, G.; Hokkanen, H.M.T.; Kuske, S.; Loomans, A.J.M.; Menzler-Hokkanen, I.; van Rijn, P.C.J.; Thomas, M.B.; Tommasini, M.G.; Zeng, Q.Q. Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl 2003, 48, 3–38. [Google Scholar] [CrossRef]
- Yano, E. Recent development of biological control and IPM in greenhouses in Japan. J. Asia Pac. Entomol. 2004, 7, 5–11. [Google Scholar] [CrossRef]
- McMurtry, J.A.; Croft, B.A. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol. 1997, 42, 291–321. [Google Scholar]
- Abad-Moyano, R.; Pina, T.; Pérez-Panadés, J.; Carbonell, E.A.; Urbaneja, A. Efficacy of Neoseiulus. californicus and Phytoseiulus. persimilis in suppression of Tetranychus. urticae in young clementine plants. Exp. Appl. Acarol. 2010, 50, 317–328. [Google Scholar] [CrossRef]
- Tauber, C.A.; Tauber, M.J. Insect seasonal cycles: Genetics and evolution. Annu. Rev. Ecol. Syst. 1981, 12, 281–308. [Google Scholar]
- Denlinger, D.L. Why study diapause? Entomol. Res. 2008, 38, 1–9. [Google Scholar]
- Barker, R.J.; Cohen, C.F.; Mayer, A. Photoflashes: a potential new tool for control of insect populations. Science 1964, 145, 1195–1197. [Google Scholar]
- Ankersmit, G.W. The photoperiod as a control agent against Adoxophyes. reticulana (Lepidoptera; Tortricidae). Entomol. Exp. Appl. 1968, 11, 231–240. [Google Scholar] [CrossRef]
- Berlinger, M.J.; Ankersmit, G.W. Manipulation with the photoperiod as a method of control of Adoxophyes. orana (Lepidoptera; Tortricidae). Entomol. Exp. Appl. 1976, 19, 96–107. [Google Scholar] [CrossRef]
- Hayes, D.K.; Sullivan, W.N.; Oliver, M.Z.; Schechter, M.S. Photoperiod manipulation of insect diapause: A method of pest control? Science 1970, 169, 382–383. [Google Scholar]
- Hayes, D.K.; Hewing, A.N.; Odesser, D.B.; Sullivan, W.N.; Schechter, M.S. The effect on diapause of photoperiod manipulation at different temperatures. In Chronobiology; Scheving, L.E., Halberg, F., Pauly, J.E., Eds.; Igaku-shoin: Tokyo, Japan, 1974; pp. 593–596. [Google Scholar]
- Hayes, D.K.; Sullivan, W.N.; Schechter, M.S.; Cawley, B.M.; Campbell, L.E. European corn borer: effect of manipulated photoperiods on survival in the field. J. Econ. Entomol. 1979, 72, 61–63. [Google Scholar]
- Schechter, M.S.; Hayes, D.K.; Sullivan, W.N. Manipulation of photoperiod to control insects. Israel J. Entomol. 1971, 6, 143–166. [Google Scholar]
- Hayes, D.K.; Sullivan, W.N.; Adler, V.E.; Schechter, M.S. The effect of added light pulses on overwintering and diapause, under natural light and temperature conditions, of four species of Lepidoptera. Environ. Entomol. 1974, 3, 863–865. [Google Scholar]
- Sáringer, G. Photoperiod as a potential control against Cydia pomonella Linné (Lep.; Tortricidae). In Proc. 2nd Egyptian-Hungarian Conf. Plant Prot.; Univ. Alexandria: Alexandria, Egypt, 1982; pp. 127–132. [Google Scholar]
- Sullivan, W.N.; Oliver, M.Z.; Hayes, D.K.; Schechter, M.S. Photoperiod manipulation to control diapause in the pink bollworm, Pectinophora. gossypiella. Experientia 1970, 26, 1101–1102. [Google Scholar] [CrossRef]
- Beach, R.F.; Craig, C.B. Photoinhibition of diapause in field populations of Aedes. atropalpus. Environ. Entomol. 1979, 8, 392–396. [Google Scholar]
- Sáringer, G. Illumination for half an hour at a time in autumn, in the scotophase of the photoperiod, as a possible ecological method of controlling the turnip sawfly Athalia. rosae L. (Hym., Tenthredinidae). Z. Angew. Entomol. 1983, 96, 278–291. [Google Scholar]
- Suzuki, T.; Takeda, M. Utilizing LED technology for arthropod pest control. In Handbook of Light Emitting and Schottky Diode Research; Chen, N.P., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 345–360. [Google Scholar]
- Veerman, A. Diapause. In Spider Mites. Their biology,Natural Enemies and Control; Helle, W., Sabelis, M.W., Eds.; Elsevier: Amsterdam, Netherlands, 1985; Volume 1A, pp. 279–316. [Google Scholar]
- Suzuki, T.; Takeda, M. Diapause-inducing signals prolong nymphal development in the two-spotted spider mite, Tetranychus. urticae. Physiol. Entomol. 2009, 34, 278–283. [Google Scholar] [CrossRef]
- Shah, M.; Suzuki, T.; Ghazy, N.A.; Amano, H.; Ohyama, K. Effect of photoperiod on immature development and diapause induction in the Kanzawa spider mite, Tetranychus. kanzawai (Acari: Tetranychidae). Exp. Appl. Acarol. 2011, 55, 183–190. [Google Scholar] [CrossRef]
- Vaz Nunes, M.; Veerman, A. Light-break experiments and photoperiodic time measurement in the spider mite Tetranychus. urticae. J. Insect Physiol. 1984, 30, 891–897. [Google Scholar] [CrossRef]
- Shah, M.; Suzuki, T.; Ghazy, N.A.; Amano, H.; Ohyama, K. Night-interrupting light inhibits diapause induction in the Kanzawa spider mite, Tetranychus. kanzawai Kishida (Acari: Tetranychidae). J. Insect Physiol. 2011, 57, 1185–1189. [Google Scholar] [CrossRef]
- Suzuki, T.; Shah, M.; Ghazy, N.A.; Takeda, M.; Amano, H.; Ohyama, K. n improved space-saving system for testing photoperiodic responses of insects and mites: Its use in diapause experiments for the two-spotted spider mite, Tetranychus. urticae (Acari: Tetranychidae). Appl. Entomol. Zool. 2011, 46, 449–454. [Google Scholar] [CrossRef]
- Koveos, D.S.; Kroon, A.; Veerman, A. The same photoperiodic clock may control induction and maintenance of diapause in the spider mite Tetranychus. urticae. J. Biol. Rhythms. 1993, 8, 265–282. [Google Scholar] [CrossRef]
- Koveos, D.S.; Veerman, A. Accumulation of photoperiodic information during diapause development in the spider mite Tetranychus. urticae. J. Insect Physiol. 1994, 40, 701–707. [Google Scholar] [CrossRef]
- Beck, S.D. Insect thermoperiodism. Annu. Rev. Entomol. 1983, 28, 91–108. [Google Scholar] [CrossRef]
- Insect Dormancy: An Eclogical Perspective; Danks, H.V. (Ed.) Biological Survey of Canada (Terrestrial arthropods): Ottawa, Canada, 1987; p. 439.
- van Houten, Y.M.; Overmeer, W.P.J.; van Zon, A.Q.; Veerman, A. Thermoperiodic induction of diapause in the predacious mite, Amblyseius. potentillae. J. Insect Physiol. 1988, 34, 285–290. [Google Scholar] [CrossRef]
- Uchida, M. Appearance time of diapausing females and termination of diapause in the two-spotted spider mite, Tetranychus. urticae Koch and the Kanzawa spider mite Tetranychus. kanzawai Kishida on pear tree in Tottori District (Acarina: Tetranychidae) (In Japanese). Jpn. J. Appl. Entomol. Zool. 1980, 24, 175–183. [Google Scholar] [CrossRef]
- Morishita, M.; Takafuji, A. Diapause characteristics of the Kanzawa spider mite, Tetranychus. kanzawai Kishida, in pea fields of central Wakayama Prefecture, Japan (In Japanese). Jpn. J. Appl. Entomol. Zool. 1999, 43, 185–188. [Google Scholar] [CrossRef]
- Parr, W.J.; Hussey, N.W. Diapause in the glasshouse red spider mite (Tetranychus. urticae Koch): A synthesis of present knowledge. Hort. Res. 1966, 6, 1–21. [Google Scholar]
- Veerman, A. Aspects of the induction of diapause in a laboratory strain of the mite Tetranychus. urticae. J. Insect Physiol. 1977, 23, 703–711. [Google Scholar]
- Gotoh, T. Annual life cycle of the two-spotted spider mite, Tetranychus. urticae Koch (Acarina: Tetranychidae), on Ribes. rubrum L. in Sapporo: The presence of non-diapausing individuals. Appl. Entomol. Zool. 1986, 21, 454–460. [Google Scholar]
- Takafuji, A.; So, P.M.; Tuno, N. Inter- and intra-population variations in diapause attribute of the two-spotted spider mite, Tetranychus. urticae Koch, in Japan. Res. Popul. Ecol. 1991, 33, 331–344. [Google Scholar] [CrossRef]
- Suzuki, T.; Fukunaga, Y.; Amano, H.; Takeda, M.; Goto, E. Effects of light quality and intensity on diapause induction in the two-spotted spider mite, Tetranychus. urticae. Appl. Entomol. Zool. 2008, 43, 213–218. [Google Scholar] [CrossRef]
- Veerman, A.; Vaz Nunes, M. Analysis of the operation of the photoperiodic counter provides evidence in the spider mite Tetranychus. urticae. J. Comp. Physiol. A 1987, 160, 421–430. [Google Scholar] [CrossRef]
- Lees, A.D. Action spectra for the photoperiodic control of polymorphism in the aphid Megoura. viciae. J. Insect Physiol. 1981, 27, 761–771. [Google Scholar] [CrossRef]
- Goto, S.G.; Numata, H. Possible involvement of distinct photoreceptors in the photoperiodic induction of diapause in the flesh fly Sarcophaga. similis. J. Insect Physiol. 2009, 55, 401–407. [Google Scholar] [CrossRef]
- Lubbock, J. Observations on ants, bees, and wasps. IX. Color of flowers as an attraction to bees: Experiments and considerations thereon. J. Linn. Soc. Lond. Zool. 1882, 16, 110–112. [Google Scholar] [CrossRef]
- Silberglied, R.E. Communication in the ultraviolet. Annu. Rev. Ecol. Syst. 1979, 10, 373–398. [Google Scholar]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef]
- Costa, H.S.; Robb, K.L. Effects of ultraviolet-absorbing greenhouse plastic films on flight behavior of Bemisia. argentifolii (Homoptera: Aleyrodidae) and Frankliniella. occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 1999, 92, 557–562. [Google Scholar]
- Costa, H.S.; Robb, K.L.; Wilen, C.A. Field trials measuring the effects of ultraviolet-absorbing greenhouse plastic films on insect populations. J. Econ. Entomol. 2002, 95, 113–120. [Google Scholar] [CrossRef]
- Antignus, Y.; Nestel, D.; Cohen, S.; Lapidot, M. Ultraviolet-deficient greenhouse environment affects whitefly attraction and flight-behavior. Environ. Entomol. 2001, 30, 394–399. [Google Scholar] [CrossRef]
- Beehler, B.C.; Przybyszewski, J.; Box, H.B.; Kulesz-Martin, M.F. Formation of 8-hydroxydeoxyguanosine within DNA of mouse keratinocytes exposed in culture to UVB and H2O2. Carcinogenesis 1992, 13, 2003–2007. [Google Scholar] [CrossRef]
- Barcelo, J.A. Photoeffects of visible and ultraviolet radiation on the two-spotted spider mite, Tetranychus. urticae. Photochem. Photobiol. 1981, 33, 703–706. [Google Scholar] [CrossRef]
- Ohtsuka, K.; Osakabe, M. Deleterious effects of UV-B radiation on herbivorous spider mites: they can avoid it by remaining on lower leaf surfaces. Environ. Entomol. 2009, 38, 920–929. [Google Scholar] [CrossRef]
- Suzuki, T.; Watanabe, M.; Takeda, M. UV tolerance in the two-spotted spider mite, Tetranychus. urticae. J. Insect Physiol. 2009, 55, 649–654. [Google Scholar] [CrossRef]
- Sakai, Y.; Osakabe, M. Spectrum-specific damage and solar ultraviolet radiation avoidance in the two-spotted spider mite. Photochem. Photobiol. 2010, 86, 925–932. [Google Scholar]
- Sakai, Y.; Sudo, M.; Osakabe, M. Seasonal changes in the deleterious effects of solar ultraviolet-B radiation on eggs of the twospotted spider mite, Tetranychus. urticae (Acari: Tetranychidae). Appl. Entomol. Zool. 2012, 47, 67–73. [Google Scholar] [CrossRef]
- Watanabe, M.; Furuya, M.; Miyoshi, Y.; Inoue, Y.; Iwahashi, I.; Matsumoto, K. Design and performance of the Okazaki Large Spectrograph for photobiological research. Photochem. Photobiol. 1982, 36, 491–498. [Google Scholar] [CrossRef]
- Mégevand, B.; Klay, A.; Gnanvossou, D.; Paraiso, G. Maintenance and mass rearing of phytoseiid predators of the cassava green mite. Exp. Appl. Acarol. 1993, 17, 115–128. [Google Scholar]
- Mites (Acari.) for Pest Control; Gerson, U.; Smiley, R.L.; Ochoa, R. (Eds.) Blackwell: Oxford, UK, 2003; p. 539.
- Mites of Greenhouses: Identification, Biology and Control; Zhang, Z.Q. (Ed.) CABI Publishing: Wallingford, UK, 2003; p. 244.
- Plant. Mites and Sociality: Diversity and Evolution; Saito, Y. (Ed.) Springer: Tokyo, Japan, 2010; p. 187.
- Ghazy, N.A.; Suzuki, T.; Amano, H.; Ohyama, K. Effects of air temperature and water vapor pressure deficit on storage of the predatory mite Neoseiulus. californicus (Acari: Phytoseiidae). Exp. Appl. Acarol. 2012, 58, 111–120. [Google Scholar] [CrossRef]
- Morrison, R.K.; King, E.G. Mass production of natural enemies. In Biological Control by Augmentation of Natural Enemies; Ridgway, R.L., Vinson, S.B., Eds.; Plenum Press: New York, NY, USA, 1977; pp. 183–217. [Google Scholar]
- Stinner, R.E. Efficacy of inundative releases. Annu. Rev. Entomol. 1977, 22, 515–531. [Google Scholar]
- Bueno, R., Jr.; Van Cleave, H.W. The effect of temperature and host density on the reproduction of Aphelinus. perpallidus. Southwest Entomol. 1997, 22, 39–51. [Google Scholar]
- Leopold, R.A. Cold storage of insects for integrated pest management. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; Hallman, G.J., Denlinger, D.L., Eds.; Westview Press: New York, NY, USA, 1998; pp. 235–267. [Google Scholar]
- López, S.N.; Botto, E. Effect of cold storage on some biological parameters of Eretmocerus. corni and Encarsia. formosa (Hymenoptera: Aphelinidae). Biol. Control. 2005, 33, 123–130. [Google Scholar] [CrossRef]
- Morewood, W.D. Cold storage of Phytoseiulus. persimilis (Phytoseiidae). Exp. Appl. Acarol. 1992, 13, 231–236. [Google Scholar] [CrossRef]
- Ghazy, N.A.; Suzuki, T.; Shah, M.; Amano, H.; Ohyama, K. Using high relative humidity and low air temperature as a long-term storage strategy for the predatory mite Neoseiulus. californicus (Gamasida: Phytoseiidae). Biol. Control. 2012, 60, 241–246. [Google Scholar] [CrossRef]
- Ghazy, N.A.; Suzuki, T.; Shah, M.; Amano, H.; Ohyama, K. Effect of long-term cold storage of the predatory mite Neoseiulus. californicus at high relative humidity on post-storage biological traits. BioControl 2012, 57, 635–641. [Google Scholar] [CrossRef]
- Colinet, H.; Boivin, G. Insect parasitoids cold storage: a comprehensive review of factors of variability and consequences. Biol. Control. 2011, 58, 83–95. [Google Scholar]
- Warthon, G.W. Water balance in insects. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gibert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; Volume 4, pp. 565–603. [Google Scholar]
- Hamamura, T.; Shinkaji, N.; Ashihara, W. Studies on the low temperature storage of Phytoseiulus. persimilis Athias-Henriot (Acarina: Phytoseiidae). Bull. Fruit Tree Res. Stn. 1978, E2, 83–90. [Google Scholar]
- Kim, J.H.; Kim, H.Y.; Han, M.J.; Choi, M.Y.; Hwang, S.J.; Lee, M.S. Cold storage effect on the biological characteristics of Orius. laevigatus (Fieber) (Hemiptera: Anthocoridae) and Phytoseiulus. persimilis Athias-Henriot (Acari: Phytoseiidae). Kor. J. Appl. Entomol. 2009, 48, 361–368. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z. Potential long-term storage of the predatory mite Phytoseiulus. persimilis. BioControl 2010, 55, 639–644. [Google Scholar] [CrossRef]
- Nicoli, G.; Galazzi, D. Quality control of cold stored Phytoseiulus persimilis Athias-Henriot (Acarina: Phytoseiidae). Boll. Ist. Ent. Univ. Bologna 1998, 52, 61–73. [Google Scholar]
- Ghazy, N.A.; Suzuki, T.; Amano, H.; Ohyama, K. Humidity-controlled cold storage of Neoseiulus. californicus (Acari: Phytoseiidae): Effects on male survival and reproductive ability. J. Appl. Entomol. 2012. [Google Scholar] [CrossRef]
- Gillespie, D.R.; Ramey, C.A. Life history and cold storage of Amblyseius. cucumeris (Acarina: Phytoseiidae). J. Entomol. Soc. B.C. 1988, 85, 71–76. [Google Scholar]
- Suzuki, T.; Ghazy, N.A.; Amano, H.; Ohyama, K. A high-performance humidity control system for tiny animals: Demonstration of its usefulness in testing egg hatchability of the two-spotted spider mite, Tetranychus. urticae. Exp. Appl. Acarol. 2012, 58, 101–110. [Google Scholar] [CrossRef]
- Kawashima, M.; Jung, C. Overwintering sites of the predacious mite Neoseiulus. californicus (McGregor) (Acari: Phytoseiidae) in Satsuma mandarin orchards on Jeju Island, Korea. Appl. Entomol. Zool. 2010, 45, 191–199. [Google Scholar] [CrossRef]
- Kawashima, M.; Jung, C. Effects of sheltered ground habitats on the overwintering potential of the predacious mite Neoseiulus. californicus (Acari: Phytoseiidae) in apple orchards on mainland Korea. Exp. Appl. Acarol. 2011, 55, 375–388. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Suzuki, T. Environmental Engineering Approaches toward Sustainable Management of Spider Mites. Insects 2012, 3, 1126-1142. https://doi.org/10.3390/insects3041126
Suzuki T. Environmental Engineering Approaches toward Sustainable Management of Spider Mites. Insects. 2012; 3(4):1126-1142. https://doi.org/10.3390/insects3041126
Chicago/Turabian StyleSuzuki, Takeshi. 2012. "Environmental Engineering Approaches toward Sustainable Management of Spider Mites" Insects 3, no. 4: 1126-1142. https://doi.org/10.3390/insects3041126
APA StyleSuzuki, T. (2012). Environmental Engineering Approaches toward Sustainable Management of Spider Mites. Insects, 3(4), 1126-1142. https://doi.org/10.3390/insects3041126