
Functional Elucidation of Vitellogenin receptor Activity in Apis mellifera in Response to Abiotic Stress


Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insect Sample Collection and Processing
2.2. RNA Extraction, cDNA Synthesis, and AmVgR Gene Cloning
2.3. Real-Time Quantitative Polymerase Chain Reaction Analysis (RT-qPCR)
2.4. Anti-AmVgR Antibody Preparation
2.5. Western Blot Analyses (WB)
2.6. Bioinformatics Analysis of the AmVgR Gene
2.7. RNA Interference of the AmVgR Gene
2.8. Expression Profiling of Antioxidant Genes After RNAi Treatment
2.9. Determination of Antioxidant Enzyme Activity and Metabolites After RNAi Treatment
2.10. Detection of Oxidative Stress Tolerance in Bees After RNAi Treatment
2.11. Data Analysis
3. Results
3.1. AmVgR Gene Isolation and Structure Specificity Analysis
3.2. Identification and Analysis of the Promoter Region
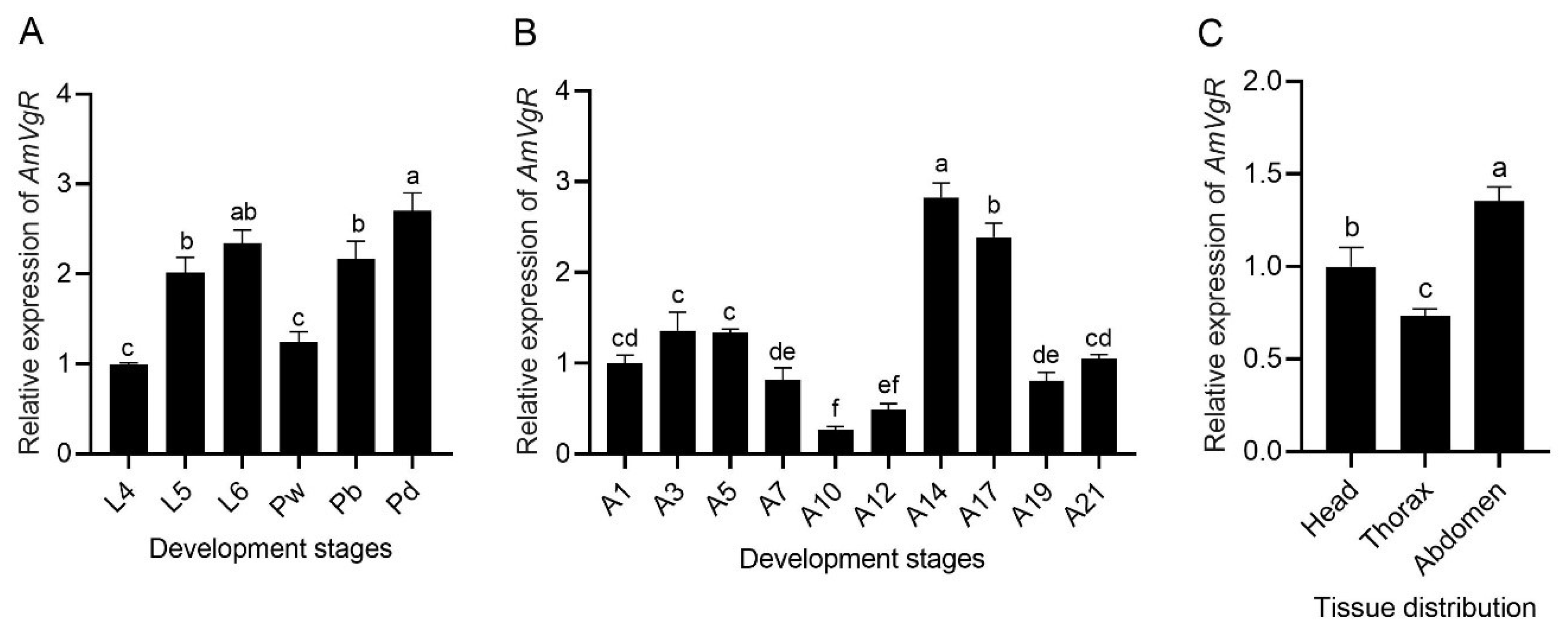
3.3. Analysis of the Spatiotemporal Expression Profiles of AmVgR
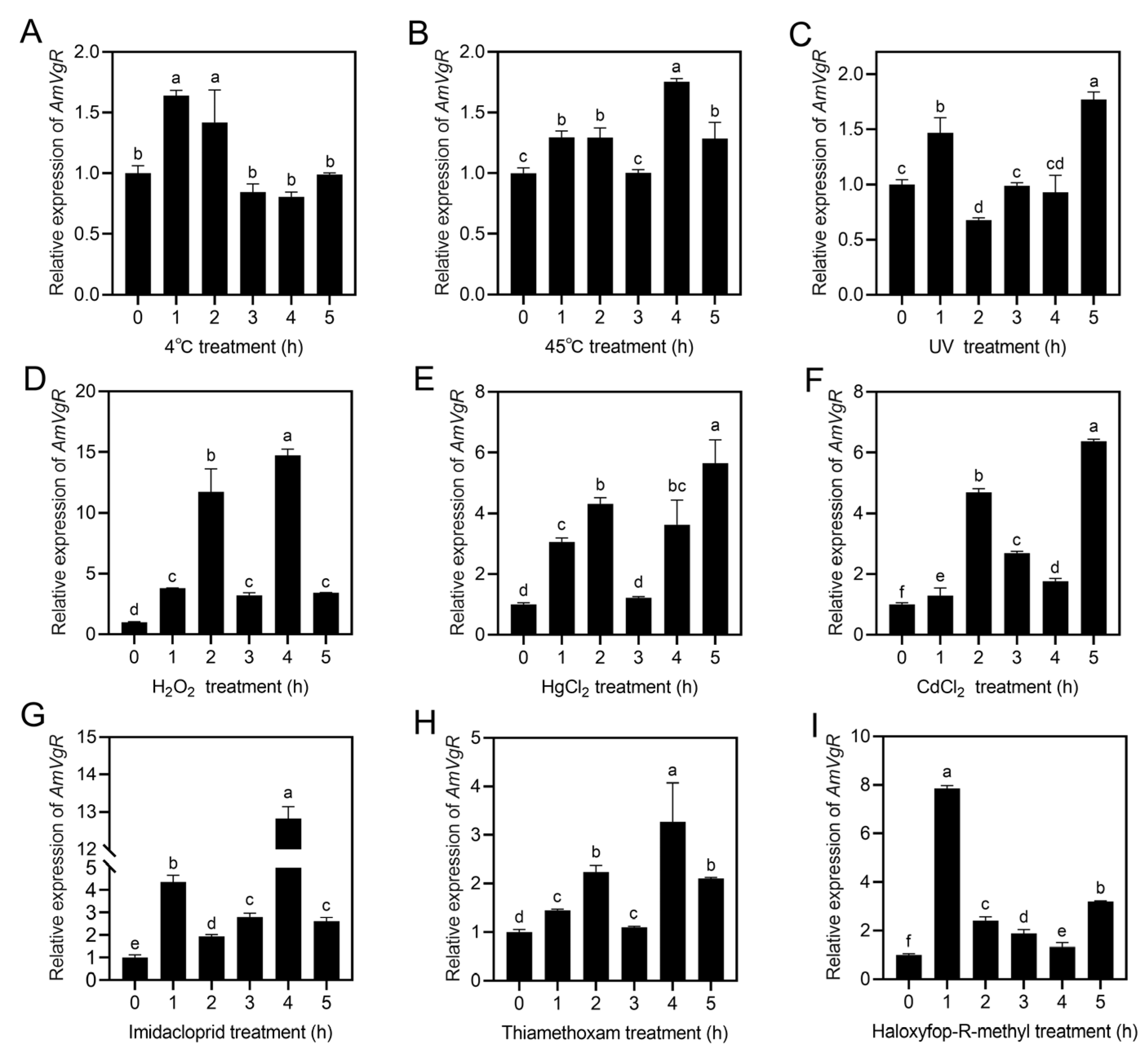
3.4. AmVgR Gene Expression Profile Under Different Stress Conditions
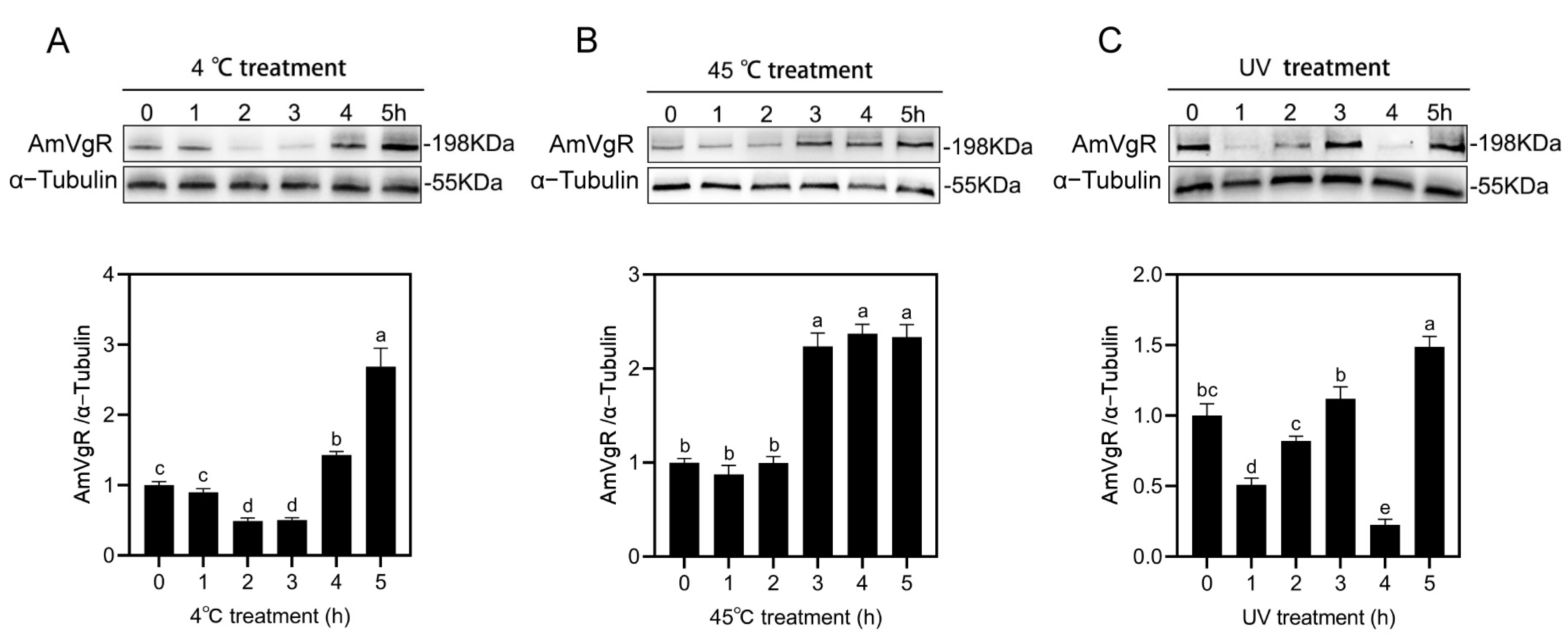
3.5. AmVgR Protein Expression Under Abiotic Stress Conditions
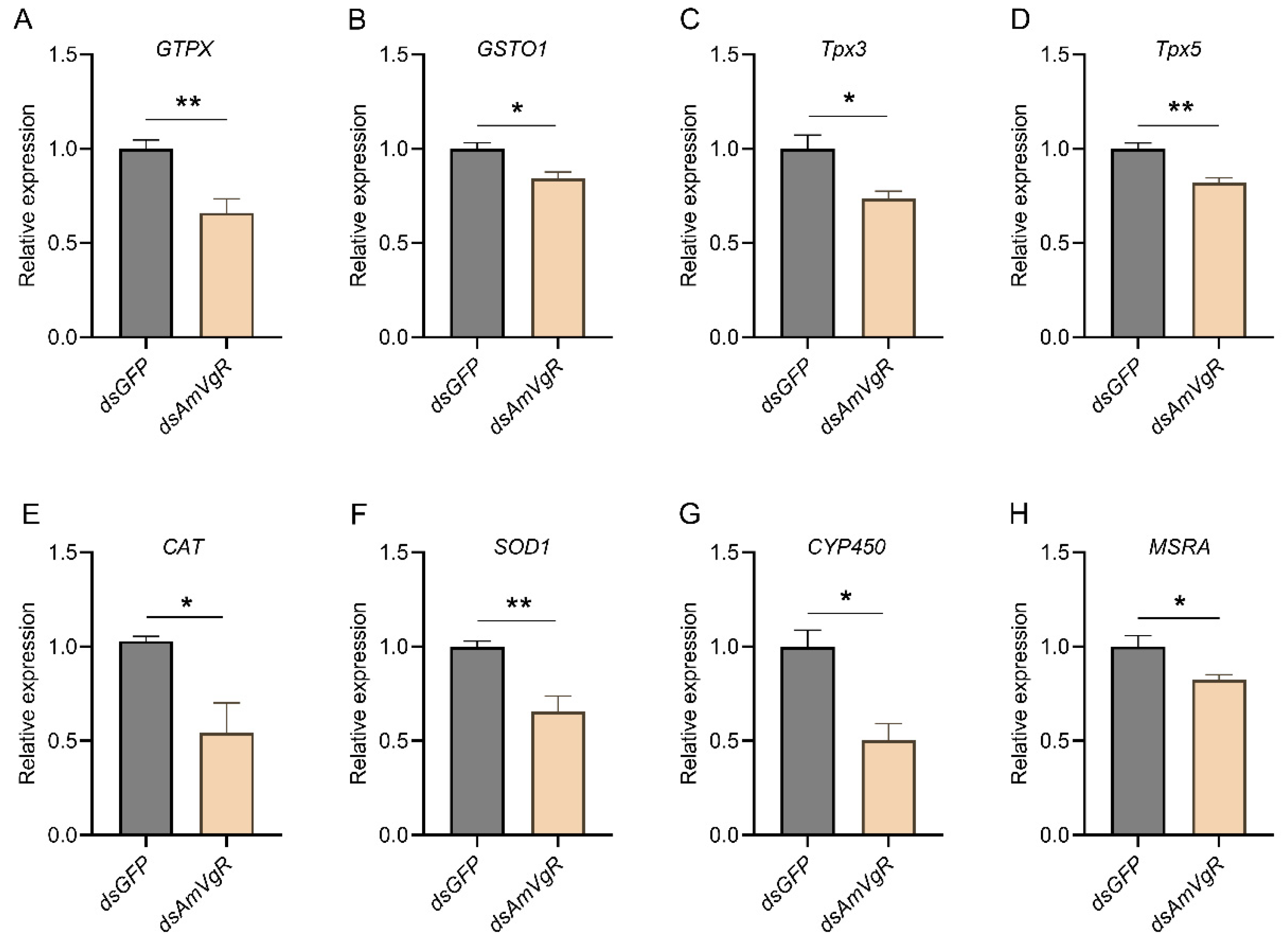
3.6. Study of the Effects of Knocking Down AmVgR on the Antioxidant Capacity in Apis mellifera
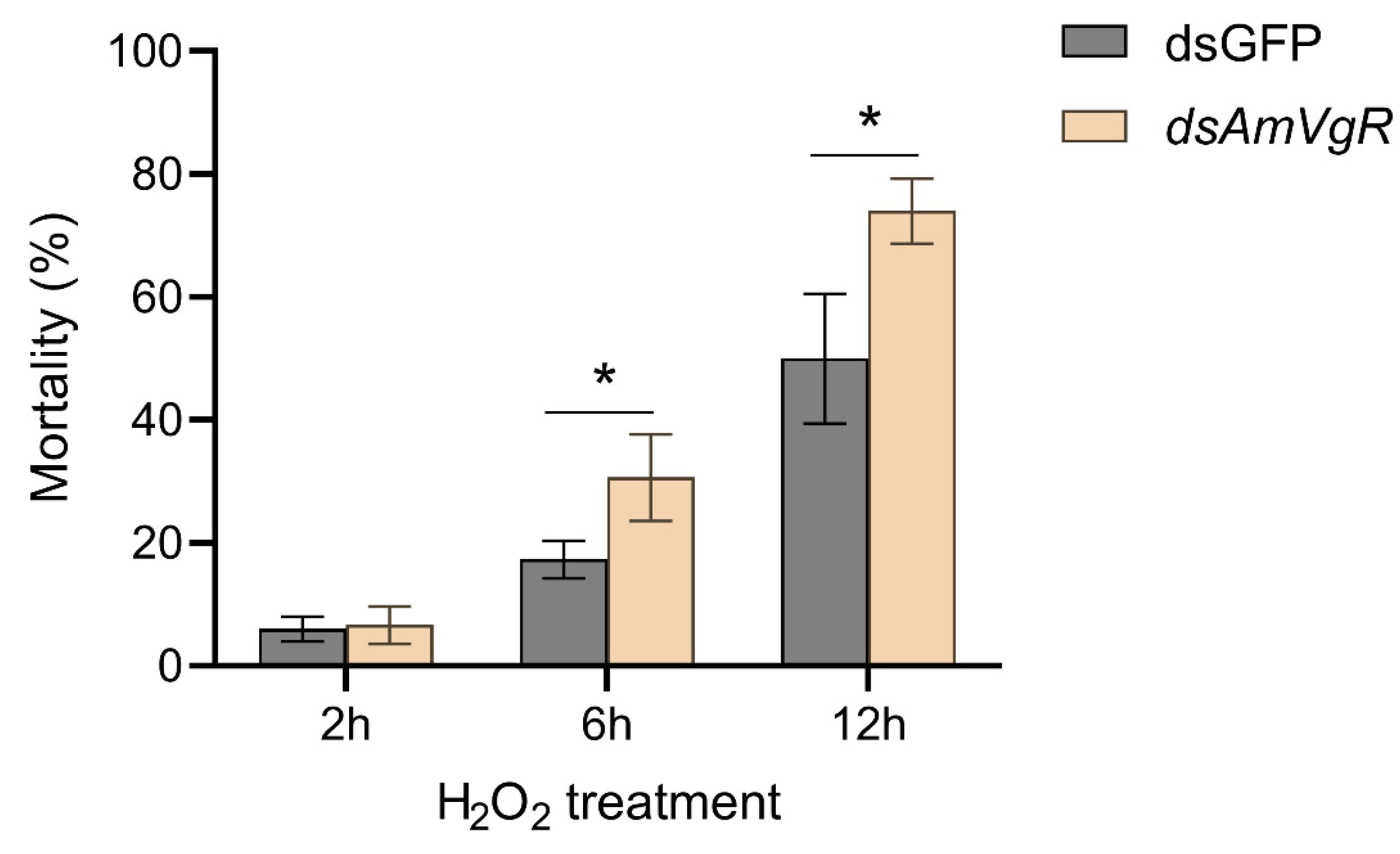
3.7. The Knockdown of AmVgR Reduces Antioxidant Stress Resistance in Apis mellifera
4. Discussion
5. Conclusions
6. Limitations and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Catarino, R.; Bretagnolle, V.; Perrot, T.; Vialloux, F.; Gaba, S. Bee pollination outperforms pesticides for oilseed crop production and profitability. Proc. Biol. Sci. 2019, 286, 20191550. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef]
- Evans, J.D.; Schwarz, R.S. Bees brought to their knees: Microbes affecting honey bee health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Grassl, J.; Holt, S.; Cremen, N.; Peso, M.; Hahne, D.; Baer, B. Synergistic effects of pathogen and pesticide exposure on honey bee (Apis mellifera) survival and immunity. J. Invertebr. Pathol. 2018, 159, 78–86. [Google Scholar] [CrossRef]
- Li, G.; Zhao, H.; Liu, Z.; Wang, H.; Xu, B.; Guo, X. The Wisdom of Honeybee Defenses Against Environmental Stresses. Front. Microbiol. 2018, 9, 722. [Google Scholar] [CrossRef]
- Kodrík, D.; Bednářová, A.; Zemanová, M.; Krishnan, N. Hormonal Regulation of Response to Oxidative Stress in Insects-An Update. Int. J. Mol. Sci. 2015, 16, 25788–25816. [Google Scholar] [CrossRef]
- Harvey, J.A.; Heinen, R.; Gols, R.; Thakur, M.P. Climate change-mediated temperature extremes and insects: From outbreaks to breakdowns. Glob. Change Biol. 2020, 26, 6685–6701. [Google Scholar] [CrossRef]
- Laycock, I.; Lenthall, K.M.; Barratt, A.T.; Cresswell, J.E. Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris). Ecotoxicology 2012, 21, 1937–1945. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Luo, Q.H.; Hou, C.S.; Wang, Q.; Dai, P.L.; Gao, J.; Liu, Y.J.; Diao, Q.Y. Sublethal effects of imidacloprid on targeting muscle and ribosomal protein related genes in the honey bee Apis mellifera L. Sci. Rep. 2017, 7, 15943. [Google Scholar] [CrossRef]
- Suchail, S.; Debrauwer, L.; Belzunces, L.P. Metabolism of imidacloprid in Apis mellifera. Pest Manag. Sci. 2004, 60, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Synergistic toxicity and physiological impact of imidacloprid alone and binary mixtures with seven representative pesticides on honey bee (Apis mellifera). PLoS ONE 2017, 12, e0176837. [Google Scholar] [CrossRef] [PubMed]
- Gregorc, A.; Alburaki, M.; Rinderer, N.; Sampson, B.; Knight, P.R.; Karim, S.; Adamczyk, J. Effects of coumaphos and imidacloprid on honey bee (Hymenoptera: Apidae) lifespan and antioxidant gene regulations in laboratory experiments. Sci. Rep. 2018, 8, 15003. [Google Scholar] [CrossRef]
- Park, H.G.; Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Lee, K.Y.; Wan, H.; Li, J.; Jin, B.R. Honeybee (Apis cerana) vitellogenin acts as an antimicrobial and antioxidant agent in the body and venom. Dev. Comp. Immunol. 2018, 85, 51–60. [Google Scholar] [CrossRef]
- Jovanovic, N.M.; Glavinic, U.; Stevanovic, J.; Ristanic, M.; Vejnovic, B.; Dolasevic, S.; Stanimirovic, Z. A Field Trial to Demonstrate the Potential of a Vitamin B Diet Supplement in Reducing Oxidative Stress and Improving Hygienic and Grooming Behaviors in Honey Bees. Insects 2025, 16, 36. [Google Scholar] [CrossRef]
- Prado, A.; Brunet, J.L.; Peruzzi, M.; Bonnet, M.; Bordier, C.; Crauser, D.; Le Conte, Y.; Alaux, C. Warmer winters are associated with lower levels of the cryoprotectant glycerol, a slower decrease in vitellogenin expression and reduced virus infections in winter honeybees. J. Insect Physiol. 2022, 136, 104348. [Google Scholar] [CrossRef] [PubMed]
- Shephard, A.M.; Brown, N.S.; Snell-Rood, E.C. Anthropogenic Zinc Exposure Increases Mortality and Antioxidant Gene Expression in Monarch Butterflies with Low Access to Dietary Macronutrients. Environ. Toxicol. Chem. 2022, 41, 1286–1296. [Google Scholar] [CrossRef]
- Torson, A.S.; Yocum, G.D.; Rinehart, J.P.; Nash, S.A.; Bowsher, J.H. Fluctuating thermal regimes prevent chill injury but do not change patterns of oxidative stress in the alfalfa leafcutting bee, Megachile rotundata. J. Insect Physiol. 2019, 118, 103935. [Google Scholar] [CrossRef]
- Diedrick, W.A.; Kanga, L.H.B.; Mallinger, R.; Pescador, M.; Elsharkawy, I.; Zhang, Y. Molecular Assessment of Genes Linked to Honeybee Health Fed with Different Diets in Nuclear Colonies. Insects 2025, 16, 374. [Google Scholar] [CrossRef]
- Sagona, S.; Coppola, F.; Nanetti, A.; Cardaio, I.; Tafi, E.; Palego, L.; Betti, L.; Giannaccini, G.; Felicioli, A. Queen Caging and Oxalic Acid Treatment: Combined Effect on Vitellogenin Content and Enzyme Activities in the First Post-Treatment Workers and Drones, Apis mellifera L. Animals 2022, 12, 3121. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Rasool, K.G.; Tufail, M.; Aldawood, A.S. Molecular characterization, expression pattern and RNAi-mediated silencing of vitellogenin receptor gene in almond moth, Cadra cautella. Insect Mol. Biol. 2020, 29, 417–430. [Google Scholar] [CrossRef]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Ciudad, L.; Piulachs, M.D.; Bellés, X. Systemic RNAi of the cockroach vitellogenin receptor results in a phenotype similar to that of the Drosophila yolkless mutant. FEBS J. 2006, 273, 325–335. [Google Scholar] [CrossRef]
- Yang, Z.M.; Lu, T.Y.; Wu, Y.; Yu, N.; Xu, G.M.; Han, Q.Q.; Liu, Z.W. The importance of vitellogenin receptors in the oviposition of the pond wolf spider, Pardosa pseudoannulata. Insect Sci. 2022, 29, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Sang, D.; Dong, B.; Hu, H.; Ji, R.; Wang, H. Molecular Features and Expression Patterns of Vitellogenin Receptor in Calliptamus italicus (Orthoptera: Acrididae). J. Insect Sci. 2019, 19, 15. [Google Scholar] [CrossRef]
- Sappington, T.W.; Raikhel, A.S. Ligand-binding domains in vitellogenin receptors and other LDL-receptor family members share a common ancestral ordering of cysteine-rich repeats. J. Mol. Evol. 1998, 46, 476–487. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, K.; Jia, X.; Du, S.; Wang, D.; Wang, L.; Qu, H.; Zhu, S.; Wang, Y.; Wang, Z.; et al. A novel extrachromosomal circular DNA related genes signature for overall survival prediction in patients with ovarian cancer. BMC Med. Genom. 2023, 16, 140. [Google Scholar] [CrossRef]
- Guidugli-Lazzarini, K.R.; do Nascimento, A.M.; Tanaka, E.D.; Piulachs, M.D.; Hartfelder, K.; Bitondi, M.G.; Simões, Z.L. Expression analysis of putative vitellogenin and lipophorin receptors in honey bee (Apis mellifera L.) queens and workers. J. Insect Physiol. 2008, 54, 1138–1147. [Google Scholar] [CrossRef]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction-Immunity Trade-Offs in Insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef]
- Stahlschmidt, Z.R.; Rollinson, N.; Acker, M.; Adamo, S.A. Are all eggs created equal? Food availability and the fitness trade-off between reproduction and immunity. Funct. Ecol. 2013, 27, 800–806. [Google Scholar] [CrossRef]
- Kelly, C.D. Reproductive and physiological costs of repeated immune challenges in female Wellington tree weta (Orthoptera: Anostostomatidae). Biol. J. Linn. Soc. 2011, 104, 38–46. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Hurd, H. Immune stimulation and malaria infection impose reproductive costs in Anopheles gambiae via follicular apoptosis. Microbes Infect. 2006, 8, 308–315. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Guo, X.; Li, Y.; Gao, H.; Guo, X.; Xu, B. sHsp22.6, an intronless small heat shock protein gene, is involved in stress defence and development in Apis cerana cerana. Insect Biochem. Mol. Biol. 2014, 53, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, H.; Lu, W.; Li, Y.; Guo, X.; Xu, B. A novel Omega-class glutathione S-transferase gene in Apis cerana cerana: Molecular characterisation of GSTO2 and its protective effects in oxidative stress. Cell Stress Chaperones 2013, 18, 503–516. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, A.J.; Atcha, H.; Hocker, J.D.; Ren, B.; Engler, A.J. AP-1 signaling modulates cardiac fibroblast stress responses. J. Cell Sci. 2023, 136, jcs261152. [Google Scholar] [CrossRef]
- Nicholls, S.; MacCallum, D.M.; Kaffarnik, F.A.; Selway, L.; Peck, S.C.; Brown, A.J. Activation of the heat shock transcription factor Hsf1 is essential for the full virulence of the fungal pathogen Candida albicans. Fungal Genet. Biol. 2011, 48, 297–305. [Google Scholar] [CrossRef]
- Erinjeri, A.P.; Wang, X.; Williams, R.; Chiozzi, R.Z.; Thalassinos, K.; Labbadia, J. HSF-1 promotes longevity through ubiquilin-1-dependent mitochondrial network remodelling. Nat. Commun. 2024, 15, 9797. [Google Scholar] [CrossRef]
- Steven, A.; Friedrich, M.; Jank, P.; Heimer, N.; Budczies, J.; Denkert, C.; Seliger, B. What turns CREB on? And off? And why does it matter? Cell. Mol. Life Sci. 2020, 77, 4049–4067. [Google Scholar] [CrossRef]
- Raftery, R.M.; Gonzalez Vazquez, A.G.; Chen, G.; O’Brien, F.J. Activation of the SOX-5, SOX-6, and SOX-9 Trio of Transcription Factors Using a Gene-Activated Scaffold Stimulates Mesenchymal Stromal Cell Chondrogenesis and Inhibits Endochondral Ossification. Adv. Healthc Mater. 2020, 9, e1901827. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.E.; Tantin, D.; Chen, J.; Sharp, P.A. B cell development and immunoglobulin transcription in Oct-1-deficient mice. Proc. Natl. Acad. Sci. USA 2004, 101, 2005–2010. [Google Scholar] [CrossRef]
- Yang, C.; Lei, L.; Wang, Y.; Xu, B.; Liu, Z. The Ontogeny and Dietary Differences in Queen and Worker Castes of Honey Bee (Apis cerana cerana). Insects 2024, 15, 855. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.H.; Wang, J.W.; Lu, K.; Zhou, J.L.; Zhou, Q.; Zhang, G.R. The first vitellogenin receptor from a Lepidopteran insect: Molecular characterization, expression patterns and RNA interference analysis. Insect Mol. Biol. 2011, 20, 61–73. [Google Scholar] [CrossRef]
- Cong, L.; Yang, W.J.; Jiang, X.Z.; Niu, J.Z.; Shen, G.M.; Ran, C.; Wang, J.J. The Essential Role of Vitellogenin Receptor in Ovary Development and Vitellogenin Uptake in Bactrocera dorsalis (Hendel). Int. J. Mol. Sci. 2015, 16, 18368–18383. [Google Scholar] [CrossRef]
- Seehuus, S.C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef]
- Miyashita, T.; Takami, A.; Takagi, R. Molecular cloning and characterization of the 5’-flanking regulatory region of the carbonic anhydrase nacrein gene of the pearl oyster Pinctada fucata and its expression. Biochem. Genet. 2012, 50, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhou, L.; Li, Z.; Gui, J.F. Expression pattern, cellular localization and promoter activity analysis of ovarian aromatase (Cyp19a1a) in protogynous hermaphrodite red-spotted grouper. Mol. Cell. Endocrinol. 2009, 307, 224–236. [Google Scholar] [CrossRef]
- Zou, Y.; Huang, W.; Gu, Z.; Gu, X. Predominant gain of promoter TATA box after gene duplication associated with stress responses. Mol. Biol. Evol. 2011, 28, 2893–2904. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Duman, R.S.; Nestler, E.J. The many faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef]
- Aathmanathan, V.S.; Arumugam, V.; Krishnan, M. Computational approach to explore the inhibitory potential of biologically derived compounds against Spodoptera litura vitellogenin receptor (VgR) using structure based virtual screening and molecular dynamics. J. Biomol. Struct. Dyn. 2022, 40, 4954–4960. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.L.; Vinson, S.B.; Pietrantonio, P.V. Oocyte membrane localization of vitellogenin receptor coincides with queen flying age, and receptor silencing by RNAi disrupts egg formation in fire ant virgin queens. FEBS J. 2009, 276, 3110–3123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ma, L.; Xiao, H.; Xie, B.; Smagghe, G.; Guo, Y.; Liang, G. Molecular Characterization and Function Analysis of the Vitellogenin Receptor from the Cotton Bollworm, Helicoverpa armigera (Hübner) (Lepidoptera, Noctuidae). PLoS ONE 2016, 11, e0155785. [Google Scholar] [CrossRef]
- Sheng, Y.; Chen, J.; Jiang, H.; Lu, Y.; Dong, Z.; Pang, L.; Zhang, J.; Wang, Y.; Chen, X.; Huang, J. The vitellogenin receptor gene contributes to mating and host-searching behaviors in parasitoid wasps. iScience 2023, 26, 106298. [Google Scholar] [CrossRef]
- Hadiwinarta, C.; Blank, A.; Somers, J.; Schal, C.; Bronzert, C.; Sharma, K.; Strader, M.B.; Hamilton, R.G.; Slater, J.E. Vitellin/Vitellogenin Is an Important Allergen in German Cockroach. Int. Arch. Allergy Immunol. 2024, 185, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Siviter, H.; Brown, M.J.F.; Leadbeater, E. Sulfoxaflor exposure reduces bumblebee reproductive success. Nature 2018, 561, 109–112. [Google Scholar] [CrossRef]
- Penn, J.; Hu, W. Benefit-cost analysis of becoming certified pollinator friendly. J. Environ. Manag. 2023, 326, 116679. [Google Scholar] [CrossRef]
- Bowler, K.; Terblanche, J.S. Insect thermal tolerance: What is the role of ontogeny, ageing and senescence? Biol. Rev. 2008, 83, 339–355. [Google Scholar] [CrossRef]
- Cameron, S.A.; Sadd, B.M. Global Trends in Bumble Bee Health. Annu. Rev. Entomol. 2020, 65, 209–232. [Google Scholar] [CrossRef]
- Traynor, K.S.; Tosi, S.; Rennich, K.; Steinhauer, N.; Forsgren, E.; Rose, R.; Kunkel, G.; Madella, S.; Lopez, D.; Eversole, H.; et al. Pesticides in honey bee colonies: Establishing a baseline for real world exposure over seven years in the USA. Environ. Pollut. 2021, 279, 116566. [Google Scholar] [CrossRef]
- Xiao, J.; He, Q.; Liu, Q.; Wang, Z.; Yin, F.; Chai, Y.; Yang, Q.; Jiang, X.; Liao, M.; Yu, L.; et al. Analysis of honey bee exposure to multiple pesticide residues in the hive environment. Sci. Total Environ. 2022, 805, 150292. [Google Scholar] [CrossRef] [PubMed]
- Sabo, R.; Staroň, M.; Sabová, L.; Majchrák, T.; Bischoff, G.; Pistorius, J.; Janke, M.; Alkassab, A.T. Honey bees for pesticide monitoring in the landscape: Which bee matrices should be used? Chemosphere 2024, 364, 143130. [Google Scholar] [CrossRef]
- Jiang, H.; Li, X.; Li, Y.; Liu, X.; Zhang, S.; Li, H.; Zhang, M.; Wang, L.; Yu, M.; Qiao, Z. Molecular and functional characterization of ribosome protein S24 in ovarian development of Macrobrachium nipponense. Int. J. Biol. Macromol. 2024, 254, 127934. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.S.; Varma, M.; Manoharan, C.; Rao, G.P.; Vijayan, K.; Mishra, R.K.; Gopalapillai, R.; Sinha, D.K. Molecular characterization and functional analysis of the vitellogenin receptor from eri silkworm, Samia ricini. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 242, 110417. [Google Scholar] [CrossRef] [PubMed]
- Phokasem, P.; Disayathanoowat, T.; Chantaphanwattana, T.; Sinpoo, C.; Chen, Y.P.; Evans, J.D.; Lee, J.H.; Krongdang, S. Comparative toxicity of oral exposure to paraquat: Survival rates and gene expression in two honey bees species; Apis mellifera and Apis cerana. Environ. Pollut. 2024, 362, 125026. [Google Scholar] [CrossRef]
- Ali, A.; Rashid, M.A.; Huang, Q.Y.; Lei, C.L. Effect of UV-A radiation as an environmental stress on the development, longevity, and reproduction of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). Environ. Sci. Pollut. Res. Int. 2016, 23, 17002–17007. [Google Scholar] [CrossRef]
- Lian, Y.; Peng, S.; Jia, J.; Li, J.; Wang, A.; Yang, S.; Zheng, R.; Yang, X.; Zhou, S. Function of Vitellogenin receptor gene in reproductive regulation of Zeugodacus cucurbitae (Coquillett) after short-term high-temperature treatment. Front. Physiol. 2022, 13, 995004. [Google Scholar] [CrossRef]
- Wongtrakul, J.; Pongjaroenkit, S.; Leelapat, P.; Nachaiwieng, W.; Prapanthadara, L.A.; Ketterman, A.J. Expression and characterization of three new glutathione transferases, an epsilon (AcGSTE2-2), omega (AcGSTO1-1), and theta (AcGSTT1-1) from Anopheles cracens (Diptera: Culicidae), a major Thai malaria vector. J. Med. Entomol. 2010, 47, 162–171. [Google Scholar] [CrossRef]
- Layalle, S.; They, L.; Ourghani, S.; Raoul, C.; Soustelle, L. Amyotrophic Lateral Sclerosis Genes in Drosophila melanogaster. Int. J. Mol. Sci. 2021, 22, 904. [Google Scholar] [CrossRef]
- Lei, L.; Gao, Z.; Zhao, Q.; Wang, C.; Wang, Y.; Wang, H.; Chi, X.; Xu, B. Identification of the cytochrome P450 gene AccCYP6A13 in Apis cerana cerana and its response to environmental stress. Pestic. Biochem. Physiol. 2024, 202, 105890. [Google Scholar] [CrossRef]
- Xiang, K.; Wu, H.; Liu, Y.; Wang, S.; Li, X.; Yang, B.; Zhang, Y.; Ma, L.; Lu, G.; He, L.; et al. MOF-derived bimetallic nanozyme to catalyze ROS scavenging for protection of myocardial injury. Theranostics 2023, 13, 2721–2733. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Feng, K.; Zhong, W.; Tan, W.; Rengaowa, S.; Hu, W. Investigating the Hepatoprotective Properties of Mulberry Leaf Flavonoids against Oxidative Stress in HepG2 Cells. Molecules 2024, 29, 2597. [Google Scholar] [CrossRef] [PubMed]
- Mo, Q.; Fu, H.; Zhao, D.; Zhang, J.; Wang, C.; Wang, D.; Li, M. Protective Effects of Mogroside V on Oxidative Stress Induced by H2O2 in Skin Fibroblasts. Drug Des. Devel. Ther. 2021, 15, 4901–4909. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsu, S.P.; Wu, M.S.; Hsu, S.M.; Chien, C.T. Effects of vitamin C infusion and vitamin E-coated membrane on hemodialysis-induced oxidative stress. Kidney Int. 2006, 69, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Alkan, Ç.; Çelik, C.; Büyükgüzel, K.; Büyükgüzel, E. Oxidative and genotoxic effect of piperazine on Galleria mellonella (Lepidoptera: Pyralidae) hemolymph. Environ. Entomol. 2023, 52, 399–407. [Google Scholar] [CrossRef]
- Kim, S.H.; Kang, K.A.; Zhang, R.; Piao, M.J.; Ko, D.O.; Wang, Z.H.; Chae, S.W.; Kang, S.S.; Lee, K.H.; Kang, H.K.; et al. Protective effect of esculetin against oxidative stress-induced cell damage via scavenging reactive oxygen species. Acta Pharmacol. Sin. 2008, 29, 1319–1326. [Google Scholar] [CrossRef]
- Kang, K.A.; Piao, M.J.; Kim, K.C.; Cha, J.W.; Zheng, J.; Yao, C.W.; Chae, S.; Hyun, J.W. Fisetin attenuates hydrogen peroxide-induced cell damage by scavenging reactive oxygen species and activating protective functions of cellular glutathione system. Vitro Cell. Dev. Biol. Anim. 2014, 50, 66–74. [Google Scholar] [CrossRef]
- Guo, Q.; Zu, M.; Liu, D.; Yan, Y.; Yang, W.; Xu, K. Roles of Vitellogenin and Its Receptor Genes in Female Reproduction of the Cigarette Beetle, Lasioderma serricorne. Insects 2025, 16, 175. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, L.; Song, H.; Liu, Z.; Zhang, G.; Wang, Y.; Xu, B. Functional Elucidation of Vitellogenin receptor Activity in Apis mellifera in Response to Abiotic Stress. Insects 2025, 16, 650. https://doi.org/10.3390/insects16070650
Lei L, Song H, Liu Z, Zhang G, Wang Y, Xu B. Functional Elucidation of Vitellogenin receptor Activity in Apis mellifera in Response to Abiotic Stress. Insects. 2025; 16(7):650. https://doi.org/10.3390/insects16070650
Chicago/Turabian StyleLei, Li, Hongyu Song, Zhenguo Liu, Ge Zhang, Ying Wang, and Baohua Xu. 2025. "Functional Elucidation of Vitellogenin receptor Activity in Apis mellifera in Response to Abiotic Stress" Insects 16, no. 7: 650. https://doi.org/10.3390/insects16070650
APA StyleLei, L., Song, H., Liu, Z., Zhang, G., Wang, Y., & Xu, B. (2025). Functional Elucidation of Vitellogenin receptor Activity in Apis mellifera in Response to Abiotic Stress. Insects, 16(7), 650. https://doi.org/10.3390/insects16070650