
Plant Signaling Mediates Interactions Between Fall and Southern Armyworms (Lepidoptera: Noctuidae) and Their Shared Parasitoid Cotesia icipe (Hymenoptera: Braconidae)

 ,
, 
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Plants Material
2.2. Laboratory Insect Rearing
2.3. Olfactometer Assays
2.4. Plant Odors Collection
2.5. Chemical Analyses
- n = number of carbons of the shorter alkane
- N = number of carbons of the longer alkane
- tr(n) = adjusted retention time of the shorter alkane
- tr(N) = adjusted retention time of the longer alkane
- tr(comp) = adjusted retention time of the identified compound
2.6. Chemicals
2.7. Identification of Plant Signals Involved in the Attraction of Spodoptera frugiperda, Spodoptera eridania, and Cotesia icipe
2.8. Data Analyses
3. Results
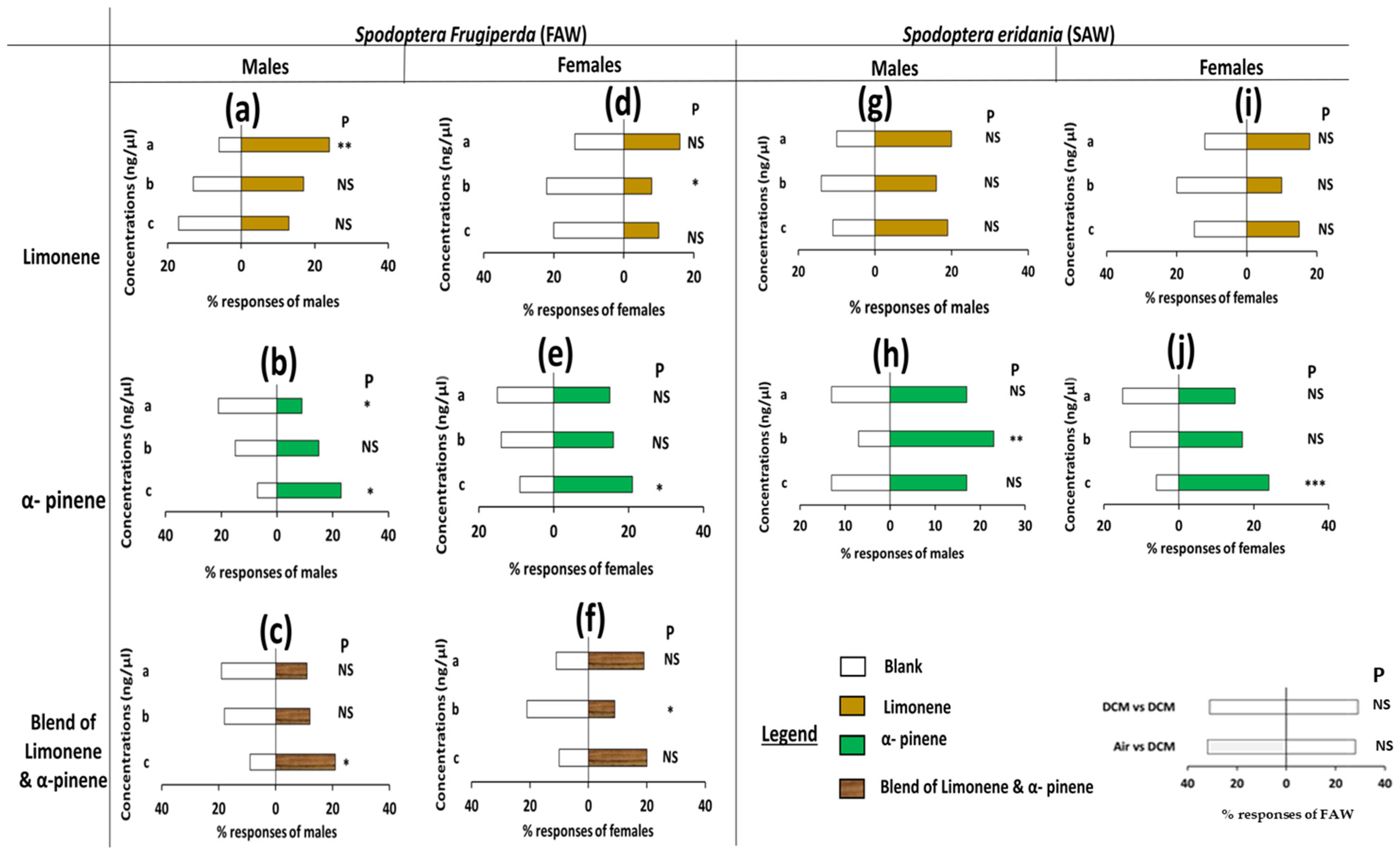
3.1. Olfactometer Assays
3.2. Chemical Analysis
3.3. Identification of Plant Signals Involved in the Attraction of FAW (Spodoptera frugiperda), SAW (Spodoptera eridania), and Cotesia icipe

4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- Guimapi, R.A.; Niassy, S.; Mudereri, B.T.; Abdel-Rahman, E.M.; Tepa-Yotto, G.T.; Subramanian, S.; Mohamed, S.A.; Thunes, K.H.; Kimathi, E.; Agboka, K.M.; et al. Harnessing data science to improve integrated management of invasive pest species across Africa: An application to Fall armyworm (Spodoptera frugiperda) (JE Smith) (Lepidoptera: Noctuidae). Glob. Ecol. Conserv. 2022, 35, e02056. [Google Scholar] [CrossRef]
- Tepa-Yotto, G.T.; Douro-Kpindou, O.K.; Koussihouédé, P.S.B.; Adjaoké, A.M.; Winsou, J.K.; Tognigban, G.; Tamò, M. Control potential of pultiple Nucleopolyhedrovirus (SfMNPV) isolated from fall armyworm in Nigeria (West Africa). Insects 2024, 15, 225. [Google Scholar] [CrossRef]
- Sisay, B.; Sevgan, S.; Weldon, C.W.; Krüger, K.; Torto, B.; Tamiru, A. Responses of the fall armyworm (Spodoptera frugiperda) to different host plants: Implications for its management strategy. Pest Manag. Sci. 2023, 79, 845–856. [Google Scholar] [CrossRef]
- Goergen, G.E. Southern armyworm, a new alien invasive pest identified in West and Central Africa. Crop Prot. 2018, 112, 371–373. [Google Scholar] [CrossRef]
- Resende, L.S.; Figueiredo, K.G.; Souza, B.H.D.; Carvalho, V.C.; Carvalho, G.A.; Presoto, J.C.; Nicolai, M.; Christoffoleti, P.J. Spodoptera eridania (Lepidoptera: Noctuidae): First report on Amaranthus hybridus (Amaranthaceae) in Brazil. Adv. Weed Sci. 2022, 40, e0202200024. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa–Gomez, D.R.; Roque–Specht, V.F.; de Barros, N.M. Immature stages of Spodoptera eridania (Lepidoptera: Noctuidae): Developmental parameters and host plants. J. Insect Sci. 2014, 14, 238. [Google Scholar] [CrossRef]
- Santos, K.B.D.; Meneguim, A.M.; Neves, P.M. Biologia de Spodoptera eridania (Cramer) (Lepidoptera: Noctuidae) em diferentes hospedeiros. Neotrop. Entomol. 2005, 34, 903–910. [Google Scholar] [CrossRef]
- Bortolotto, O.C.; Silva, G.V.; de Freitas Bueno, A.; Pomari, A.F.; Martinelli, S.; Head, G.P.; Carvalho, R.A.; Barbosa, G.C. Development and reproduction of Spodoptera eridania (Lepidoptera: Noctuidae) and its egg parasitoid Telenomus remus (Hymenoptera: Platygastridae) on the genetically modified soybean (Bt) MON 87701× MON 89788. Bull. Entomol. Res. 2014, 104, 724–730. [Google Scholar] [CrossRef]
- Weinberg, J.; Ota, N.; Goergen, G.; Fagbohoun, J.R.; Tepa-Yotto, G.; Kriticos, D.J. Spodoptera eridania: Current and emerging crop threats from another invasive, pesticide-resistant moth. Entomol. Gen. 2022, 42, 701–712. [Google Scholar] [CrossRef]
- Tepa-Yotto, G.T.; Gouwakinnou, G.N.; Fagbohoun, J.R.; Tamò, M.; Sæthre, M.G. Horizon scanning to assess the bioclimatic potential for the alien species Spodoptera eridania and its parasitoids after pest detection in West and Central Africa. Pest Manag. Sci. 2021, 77, 4437–4446. [Google Scholar] [CrossRef] [PubMed]
- Tepa-Yotto, G.T.; Tonnang, H.E.; Yeboah, S.; Osae, M.Y.; Gariba, A.A.; Dalaa, M.; Adomaa, F.O.; Damba, O.T.; Kyere, R.; Moutouama, F.T.; et al. Implementation Outline of Climate-Smart One Health: A System-Thinking Approach. Sustainability 2024, 16, 6652. [Google Scholar] [CrossRef]
- Zhang, D.D.; Xiao, Y.T.; Xu, P.J.; Yang, X.M.; Wu, Q.L.; Wu, K.M. Insecticide resistance monitoring for the invasive populations of fall armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 783–791. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.; Ullah, F.; Zhang, Z.; Zhang, J.; Huang, J.; Chen, L.; Siddiqui, J.A.; Ren, X.; Ren, X.; et al. Characterization of indoxacarb resistance in the fall armyworm: Selection, inheritance, cross-resistance, possible biochemical mechanisms, and fitness costs. Biology 2022, 11, 1718. [Google Scholar] [CrossRef]
- Peter, E.; Tamiru, A.; Sevgan, S.; Dubois, T.; Kelemu, S.; Kruger, K.; Torto, B.; Yusuf, A. Companion crops alter olfactory responses of the fall armyworm (Spodoptera frugiperda) and its larval endoparasitoid (Cotesia icipe). Chem. Biol. Technol. Agric. 2023, 10, 61. [Google Scholar] [CrossRef]
- Gargi, C.; Kennedy, J.S.; Jayabal, T.D. Olfactory response of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) to the volatiles of healthy and herbivore-damaged maize plants and their profiling. Pharm. Innov. J. 2021, 10, 1061–1067. [Google Scholar]
- Mutyambai, D.M. Exploiting early herbivory-induced defense traits in Zea species for the management of Chilo partellus in East Africa (Doctoral dissertation). Ph.D. Thesis, North-West University, Potchefstroom, South Africa, 2014. Available online: https://repository.nwu.ac.za/handle/10394/15461?show=full (accessed on 18 February 2025).
- Winsou, J.K.; Tepa-Yotto, G.T.; Thunes, K.H.; Meadow, R.; Tamò, M.; Sæthre, M.G. Seasonal variations of Spodoptera frugiperda host plant diversity and parasitoid complex in southern and central Benin. Insects 2022, 13, 491. [Google Scholar] [CrossRef]
- Fiaboe, K.K.; Fernández-Triana, J.; Nyamu, F.W.; Agbodzavu, K.M. Cotesia icipe sp. n., a new Microgastrinae wasp (Hymenoptera, Braconidae) of importance in the biological control of Lepidopteran pests in Africa. J. Hymenopt. Res. 2017, 61, 49–64. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Borkakati, R.N.; Venkatesh, M.R.; Saikia, D.K.; Sasanka, S.B. A brief review on food recognition by insects: Use of sensory and behavioural mechanisms. J. Entomol. Zool. Stud. 2019, 7, 574–579. [Google Scholar]
- Hartlieb, E.; Rembold, H. Behavioral response of female Helicoverpa (Heliothis) armigera HB. (Lepidoptera: Noctuidae) moths to synthetic pigeonpea (Cajanus cajan L.) kairomone. J. Chem. Ecol. 1996, 22, 821–837. [Google Scholar] [CrossRef]
- Signoretti, A.G.C.; Peñaflor, M.F.G.V.; Bento, J.M.S. Fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), female moths respond to herbivore-induced corn volatiles. Neotrop. Entomol. 2012, 41, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, A.S.; Cordeiro, E.M.; Omoto, C. Agricultural insect hybridization and implications for pest management. Pest Manag. Sci. 2019, 75, 2857–2864. [Google Scholar] [CrossRef]
- Büchel, K.; Malskies, S.; Mayer, M.; Fenning, T.M.; Gershenzon, J.; Hilker, M.; Meiners, T. How plants give early herbivore alert: Volatile terpenoids attract parasitoids to egg-infested elms. Basic Appl. Ecol. 2011, 12, 403–412. [Google Scholar] [CrossRef]
- Degenhardt, J. Indirect defense responses to herbivory in grasses. Plant Physiol. 2009, 149, 96–102. [Google Scholar] [CrossRef]
- Agbodzavu, M.K.; Lagat, Z.O.; Gikungu, M.; Rwomushana, I.; Ekesi, S.; Fiaboe, K.K.M. Performance of the newly identified endoparasitoid Cotesia icipe Fernandez-Triana & Fiaboe on Spodoptera littoralis (Boisduval). J. Appl. Entomol. 2018, 142, 646–653. [Google Scholar] [CrossRef]
- Ortiz-Carreon, F.R.; Rojas, J.C.; Cisneros, J.; Malo, E.A. Herbivore-induced volatiles from maize plants attract Chelonus insularis, an egg-larval parasitoid of the fall armyworm. J. Chem. Ecol. 2019, 45, 326–337. [Google Scholar] [CrossRef]
- Sokame, B.M.; Ntiri, E.S.; Ahuya, P.; Torto, B.; Le Ru, B.P.; Kilalo, D.C.; Juma, G.; Calatayud, P.A. Caterpillar-induced plant volatiles attract conspecific and heterospecific adults for oviposition within a community of lepidopteran stemborers on maize plant. Chemoecology 2019, 29, 89–101. [Google Scholar] [CrossRef]
- Mann, L.; Laplanche, D.; Turlings, T.C.; Desurmont, G.A. A comparative study of plant volatiles induced by insect and gastropod herbivory. Sci. Rep. 2021, 11, 23698. [Google Scholar] [CrossRef] [PubMed]
- Shen YouLian, S.Y.; Gao Yang, G.Y.; Du YongJun, D.Y. The synergism of plant volatile compounds and sex pheromones of the tobacco cutworm moth. Spodoptera litura (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2009, 52, 1290–1297. [Google Scholar]
- Carroll, M.J.; Schmelz, E.A.; Meagher, R.L.; Teal, P.E. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings. J. Chem. Ecol. 2006, 32, 1911–1924. [Google Scholar] [CrossRef]
- Deng, J.Y.; Wei, H.Y.; Huang, Y.P.; Du, J.W. Enhancement of attraction to sex pheromones of Spodoptera exigua by volatile compounds produced by host plants. J. Chem. Ecol. 2004, 30, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Dickens, J.C. Sensitivity and responsiveness of neurons associated with male-specific Trichoid sensilla in the beet armyworm, Spodoptera exigua, to volatile emissions of sympatric Spodoptera spp. and plant odors. In Olfaction and Taste XI, Proceedings of the 11th International Symposium on Olfaction and Taste and of the 27th Japanese Symposium on Taste and Smell. Joint Meeting, Sapporo, Japan, 12–16 July 1993; Kurihara, K., Suzuki, N., Ogawa, H., Eds.; Springer: Tokyo, Japan, 1994; pp. 847–848. [Google Scholar] [CrossRef]
- Lewis, W.J.; Nordlund, D.A. Semiochemicals influencing fall armyworm parasitoid behavior: Implications for behavioral manipulation. Florida Entomol. 1984, 67, 343–349. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations (ng/μL) | Limonene | α-Pinene | (Z)-Beta-Farnesene | Methyl Dodecanoate | Isopentyl Acetate |
|---|---|---|---|---|---|
| a | 911 | 415 | 1187 | 59 | 61 |
| b | 1823 | 831 | 2374 | 118 | 122 |
| c | 3646 | 1662 | 4748 | 236 | 244 |
| (A) | ||||||
| Chemical Profiles of Healthy Plant Species | ||||||
| RT | Library/ID | Qual | Index cal. | Index lit. | Maize | Amaranth |
| 8.74 | o-Xylene | 97 | 847.09 | 855 | + | + |
| 9.28 | p-Xylene | 95 | 886.67 | 888 | + | + |
| 9.46 | n-Nonane | 81 | 899.87 | 900 | + | - |
| 10.19 | alpha-Pinene | 97 | 936.00 | 931 | + | + |
| 9.84 | Anisole | 64 | 918.81 | 918 | - | + |
| 10.01 | Cumene | 90 | 927.23 | 928 | - | + |
| 10.78 | Mesitylene | 90 | 965.44 | 972 | + | + |
| 11.15 | Phenol | 58 | 983.66 | 985 | - | + |
| 12.06 | Limonene | 97 | 1032.07 | 1027 | + | + |
| 14.97 | n-Decanal | 87 | 1209.34 | 1209 | + | + |
| 15.35 | Benzothiazole | 94 | 1235.65 | 1243 | + | - |
| 16.04 | Tridecane | 74 | 1283.45 | 1300 | - | + |
| 16.88 | Benzene, 3-cyclohexen-1-yl- | 60 | 1344.71 | 1345 | + | + |
| 17.42 | Longicyclene | 70 | 1384.59 | 1374 | + | - |
| 19.40 | trans-Calamenene | 58 | 1538.12 | 1534 | + | - |
| 20.23 | 1-Dodecanol, 2-hexyl- | 60 | 1609.32 | 1611 | - | + |
| 21.43 | Eicosane | 58 | 1712.50 | - | - | + |
| (B) | ||||||
| Chemical profile of attacked maize plant | ||||||
| RT | Library/ID | Qual | Index cal. | Index lit. | FAW | SAW |
| 6.06 | Phenyl ethyl alcohol | 53 | + | - | ||
| 8.55 | Ethylbenzene | 91 | 833.06 | 846 | - | + |
| 8.73 | o-Xylene | 97 | 847.09 | 855 | + | + |
| 9.28 | p-Xylene | 95 | 886.67 | 888 | - | + |
| 9.00 | Isopentyl acetate | 53 | 866.18 | 875 | + | - |
| 9.83 | Anisole | 96 | 918.32 | 918 | + | - |
| 10.19 | alpha-Pinene | 97 | 936.14 | 936 | + | - |
| 11.60 | n-Octanal | 59 | 1006.66 | 1008 | - | + |
| 12.05 | Limonene | 64 | 1031.46 | 1027 | + | - |
| 12.40 | 1,3,6-Octatriene, 3,7-dimethyl-, (Z)- | 95 | 1050.83 | 1038 | + | - |
| 13.57 | 4,8-Dimethyl-1,3-(Z),7-nonatriene | 74 | 1118.18 | 1113 | + | - |
| 14.11 | 2-Ethyl hexyl acetate | 53 | 1153.25 | 1159 | + | - |
| 14.97 | n-Decanal | 72 | 1209.66 | 1209 | + | + |
| 16.04 | Tridecane | 74 | 1283.60 | 1300 | - | + |
| 17.67 | Tetradecane | 91 | 1403.03 | 1400 | + | |
| 17.81 | Dodecanal | 91 | 1413.97 | 1409 | - | + |
| 18.44 | (Z)-beta-Farnesene | 83 | 1461.36 | 1455 | + | - |
| 18.62 | n-Tridecanol | 83 | 1475.15 | 1510 | - | + |
| 19.25 | Methyl dodecanoate | 95 | 1525.66 | 1528 | - | + |
| (C) | ||||||
| Chemical profile of attacked amaranth plant | ||||||
| RT | Library/ID | Qual | Index cal | Index lit. | FAW | SAW |
| 9.29 | P-Xylene | 95 | 887.49 | 888 | + | + |
| 11.19 | alpha-Methyl styrene | 91 | 985.42 | 986 | + | + |
| 11.54 | (S)-3-Ethyl-4-methyl pentanol | 59 | 1003.56 | 1020 | + | - |
| 12.07 | Limonene | 93 | 1032.04 | 1027.00 | - | + |
| 13.32 | n-Undecane | 56 | 1101.73 | 1100 | + | - |
| 14.91 | 1,3,5,7-Tetramethyl-adamantane | 70 | 1205.48 | 1214 | + | - |
| 14.95 | n-Decanal | 87 | 1208.57 | 1209.00 | - | + |
| 15.35 | Benzothiazole | 96 | 1235.64 | 1228 | + | - |
| 15.63 | Caprolactam | 94 | 1254.98 | 1244 | + | + |
| 16.32 | Tridecane | 95 | 1303.16 | 1300 | + | - |
| 16.88 | Benzene, 3-cyclohexen-1-yl- | 97 | 1345.53 | 1345.00 | - | + |
| 19.25 | Methyl dodecanoate | 91 | 1525.65 | 1528 | + | - |
| 17.80 | Dodecanal | 87 | 1413.13 | 1409.00 | - | + |
| 18.63 | n-Pentadecanol | 87 | 1476.02 | - | - | + |
| 19.26 | Dodecanoic acid, methyl ester | 94 | 1526.62 | 1528.00 | - | + |
| 19.46 | Pentadecane | 74 | 1543.86 | - | + | - |
| 20.14 | Heptadecane | 91 | 1601.38 | - | + | - |
| 21.43 | Eicosane | 80 | 1712.50 | - | - | + |
| 25.86 | Octadecanoic acid | 91 | 2160.64 | 2172.00 | - | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tepa-Yotto, G.T.; Kpongbe, H.; Winsou, J.K.; Agossadou, A.H.; Tamò, M. Plant Signaling Mediates Interactions Between Fall and Southern Armyworms (Lepidoptera: Noctuidae) and Their Shared Parasitoid Cotesia icipe (Hymenoptera: Braconidae). Insects 2025, 16, 580. https://doi.org/10.3390/insects16060580
Tepa-Yotto GT, Kpongbe H, Winsou JK, Agossadou AH, Tamò M. Plant Signaling Mediates Interactions Between Fall and Southern Armyworms (Lepidoptera: Noctuidae) and Their Shared Parasitoid Cotesia icipe (Hymenoptera: Braconidae). Insects. 2025; 16(6):580. https://doi.org/10.3390/insects16060580
Chicago/Turabian StyleTepa-Yotto, Ghislain T., Hilaire Kpongbe, Jeannette K. Winsou, Anette H. Agossadou, and Manuele Tamò. 2025. "Plant Signaling Mediates Interactions Between Fall and Southern Armyworms (Lepidoptera: Noctuidae) and Their Shared Parasitoid Cotesia icipe (Hymenoptera: Braconidae)" Insects 16, no. 6: 580. https://doi.org/10.3390/insects16060580
APA StyleTepa-Yotto, G. T., Kpongbe, H., Winsou, J. K., Agossadou, A. H., & Tamò, M. (2025). Plant Signaling Mediates Interactions Between Fall and Southern Armyworms (Lepidoptera: Noctuidae) and Their Shared Parasitoid Cotesia icipe (Hymenoptera: Braconidae). Insects, 16(6), 580. https://doi.org/10.3390/insects16060580