
Multigeneration Sublethal Chlorantraniliprole Treatment Disrupts Nutritional Metabolism and Inhibits Growth, Development, and Reproduction of Phthorimaea absoluta

 , , , ,
, , , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Chemicals
2.2. Bioassays
2.3. Selection Experiments
2.4. Life Table Data, Study, and Analysis
2.5. Determination of Nutrient Reserves
2.6. Transcriptome Sequencing
2.7. Validation of Transcriptomic Data with Quantitative Real-Time PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results
3.1. Selection of P. absoluta Resistance to Chlorantraniliprole
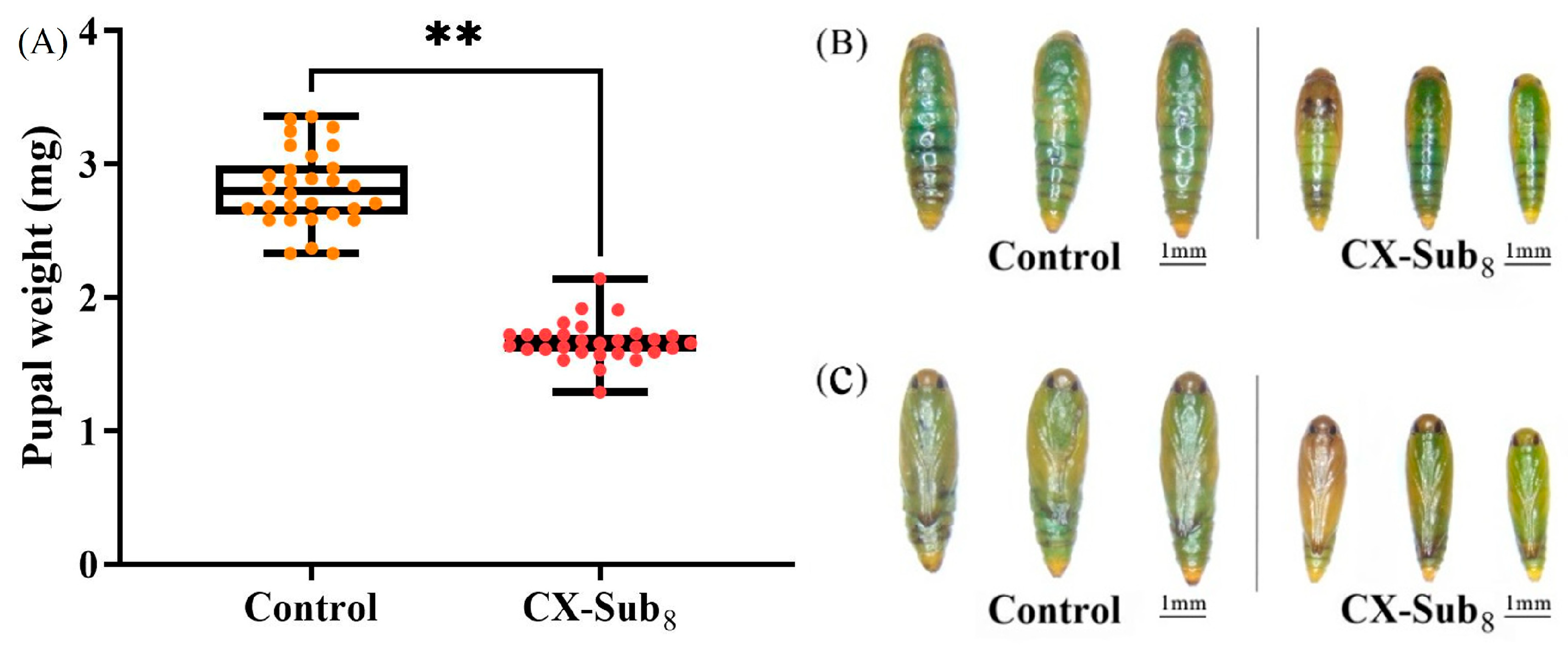
3.2. Inhibitory Effect of Chlorantraniliprole on the Growth, Development, and Reproduction of P. absoluta
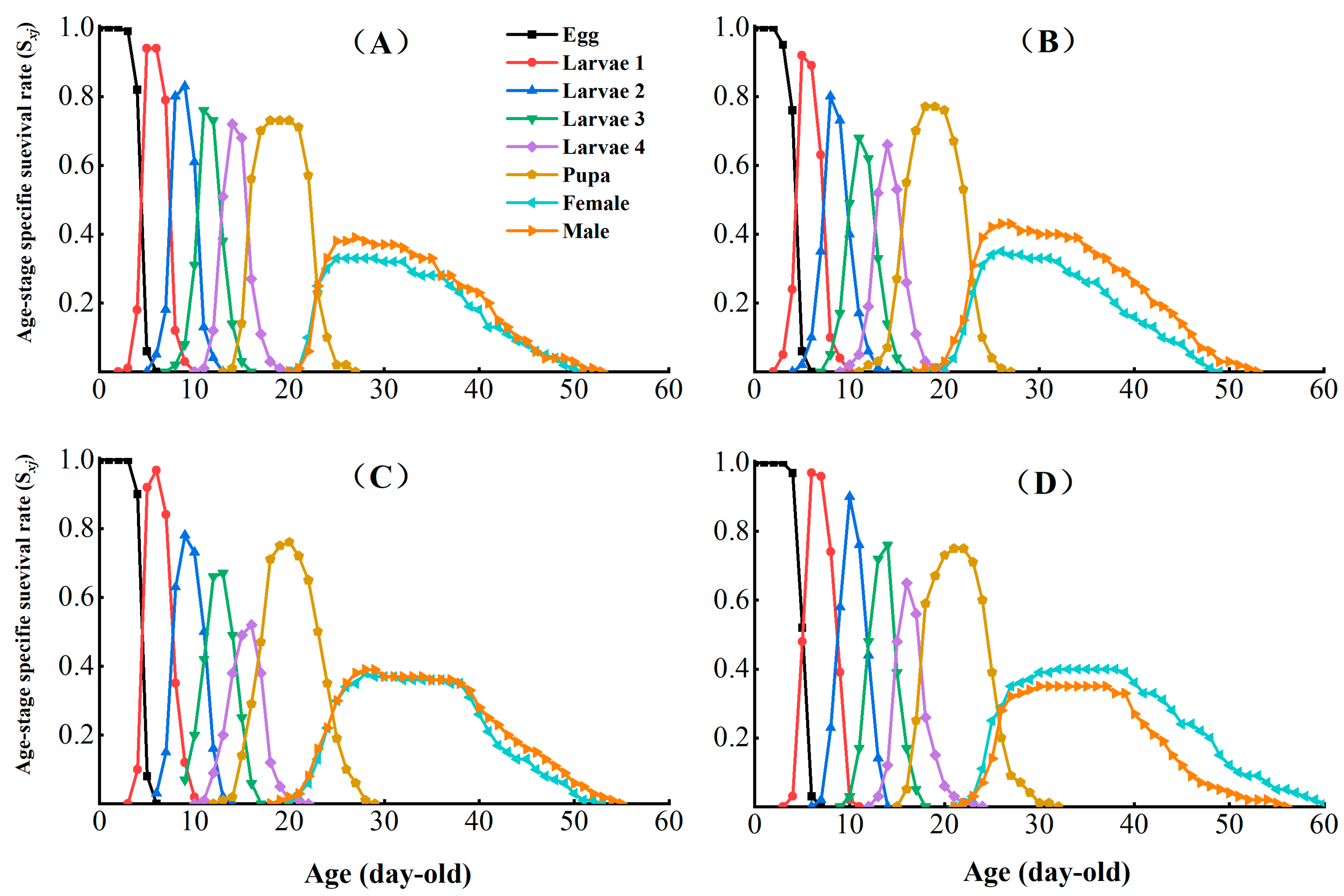
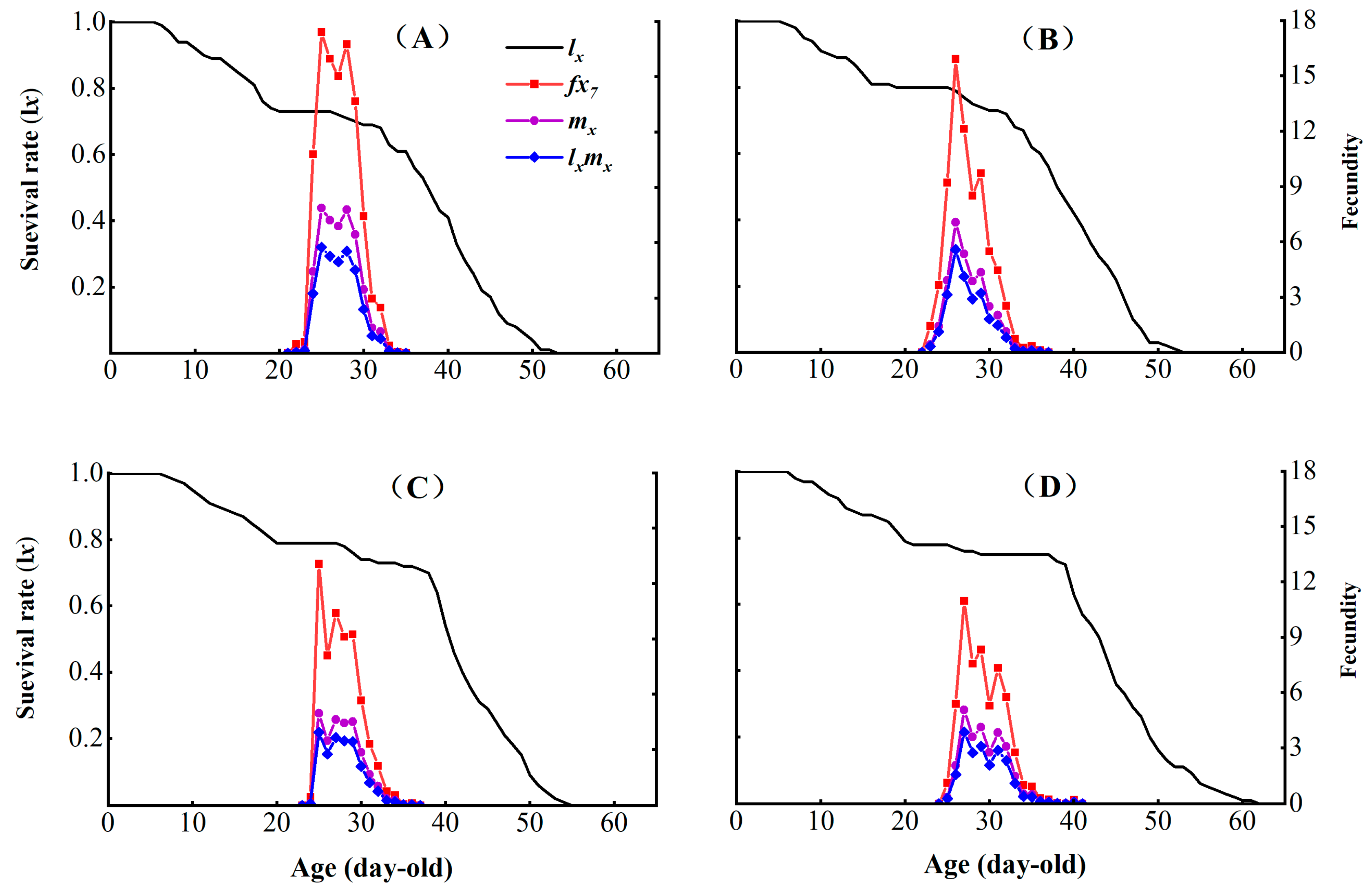
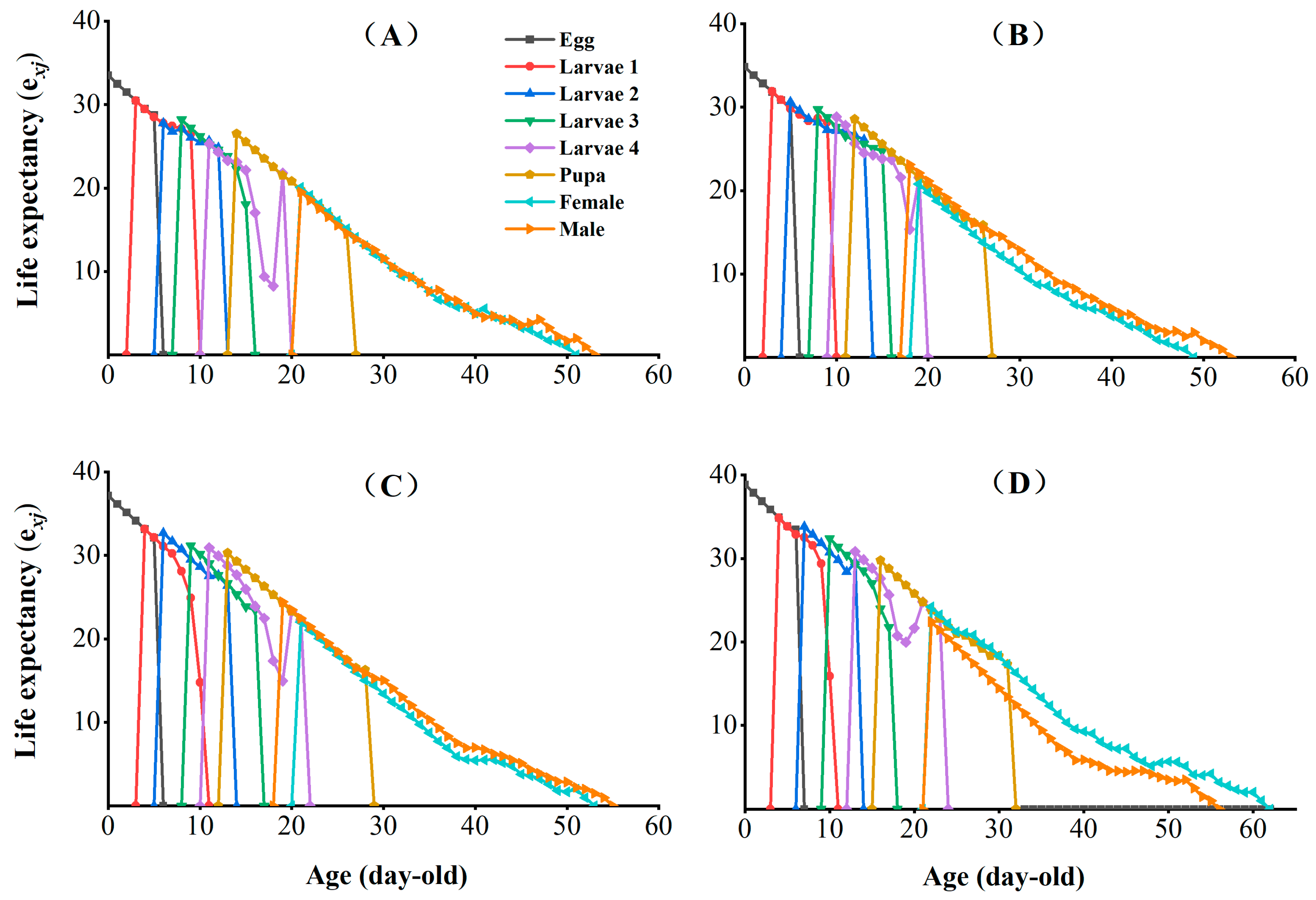
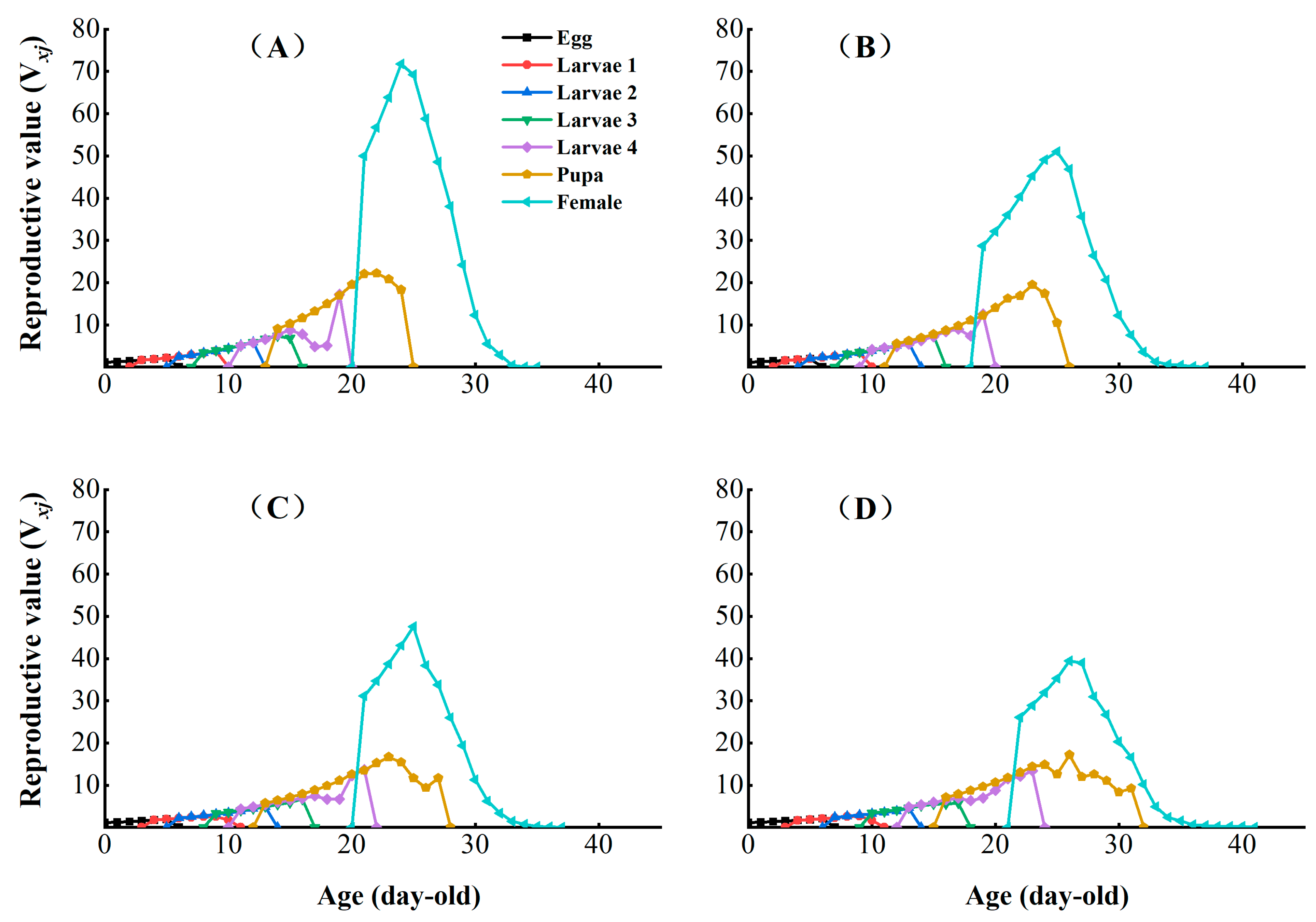
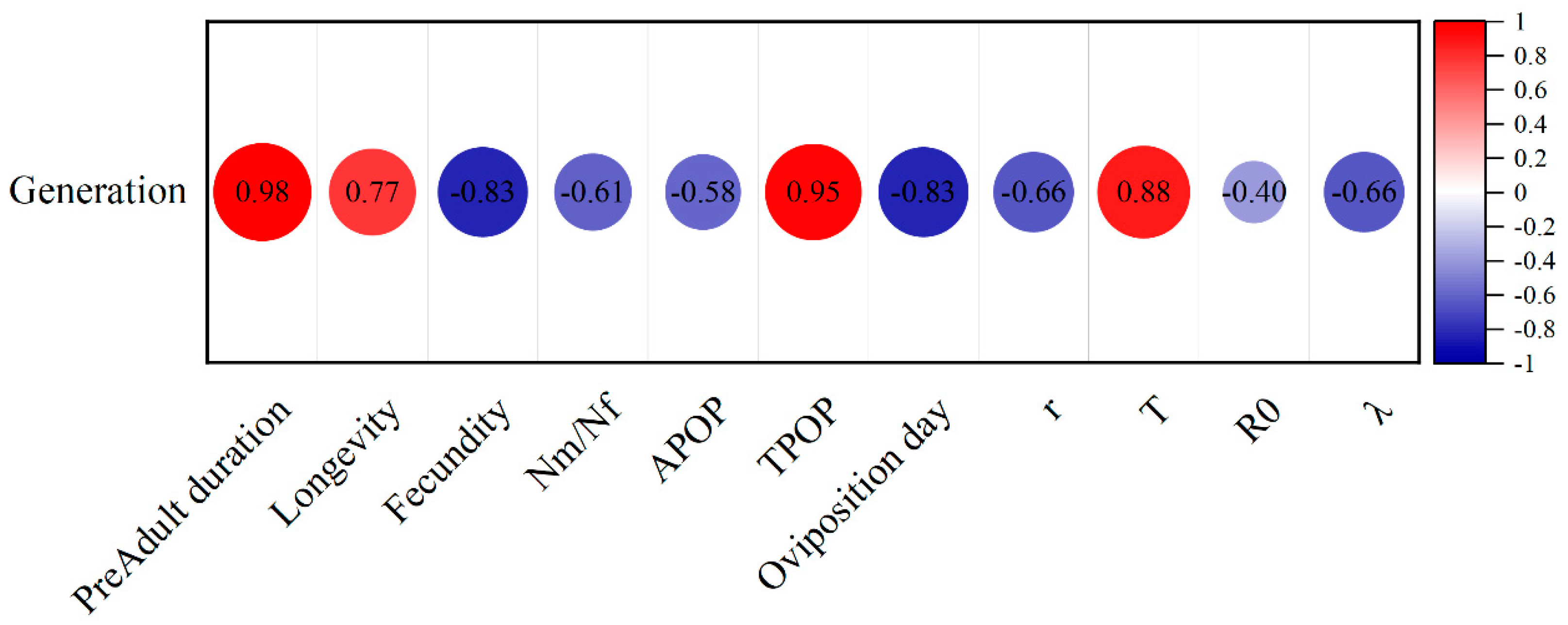
3.3. Effects of Chlorantraniliprole on the Life Table Parameters of the F2, F4, and F8 Generations of P. absoluta
3.4. Effects of Chlorantraniliprole on the Population Parameters of the F2, F4, and F8 Generations of P. absoluta
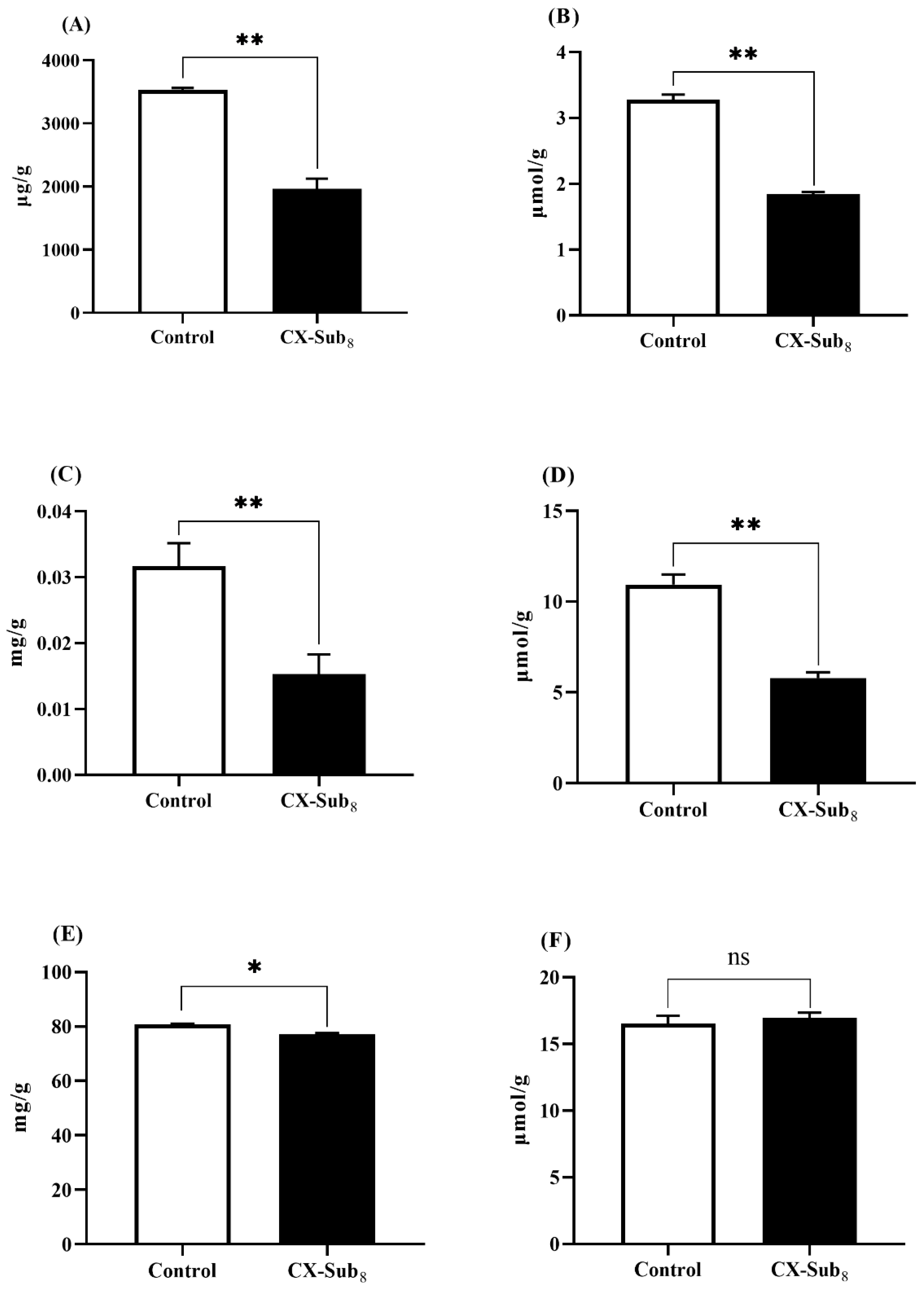
3.5. Chlorantraniliprole Reduces Nutrient Reserves in the Second-Instar Larvae of P. absoluta
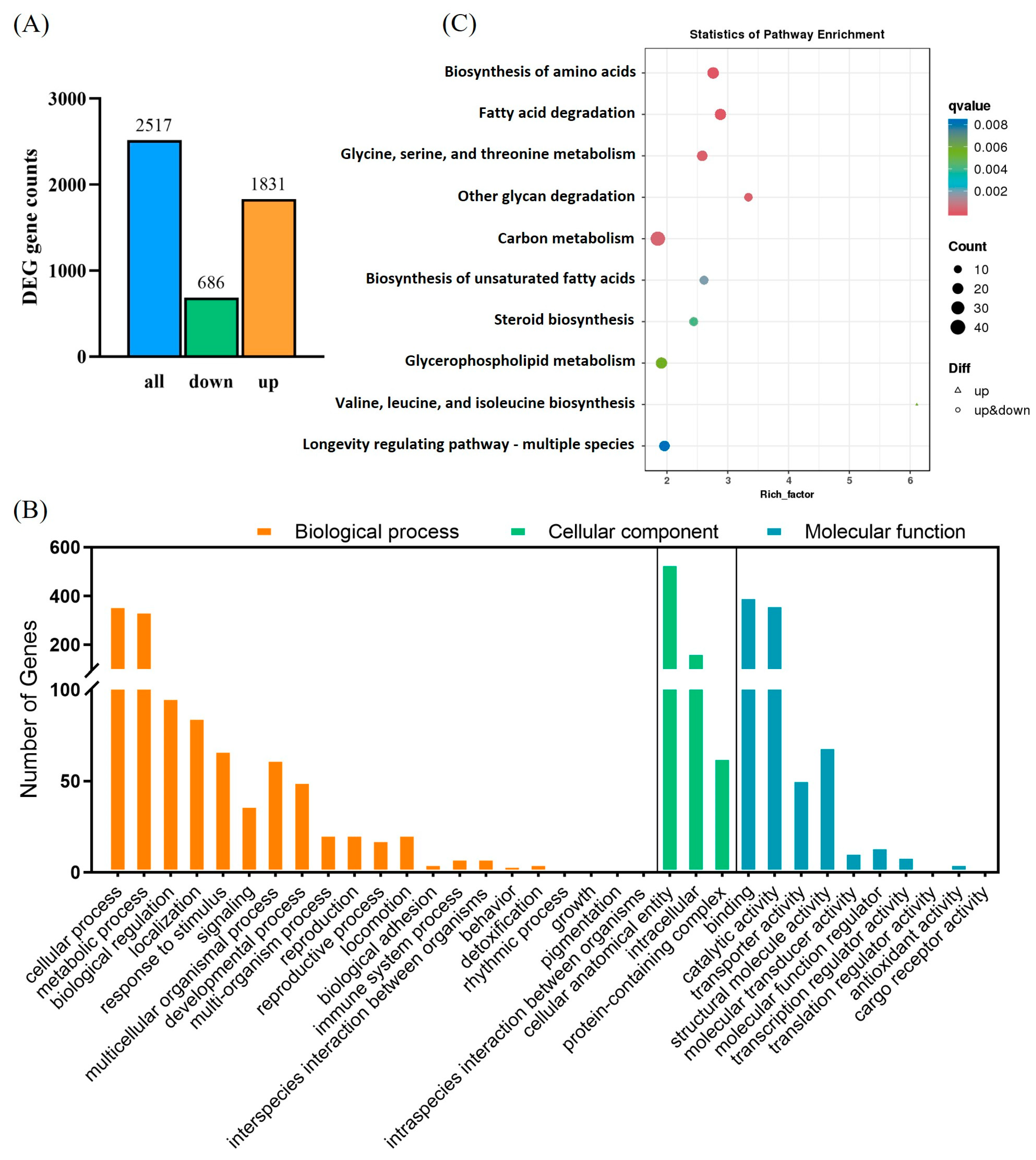
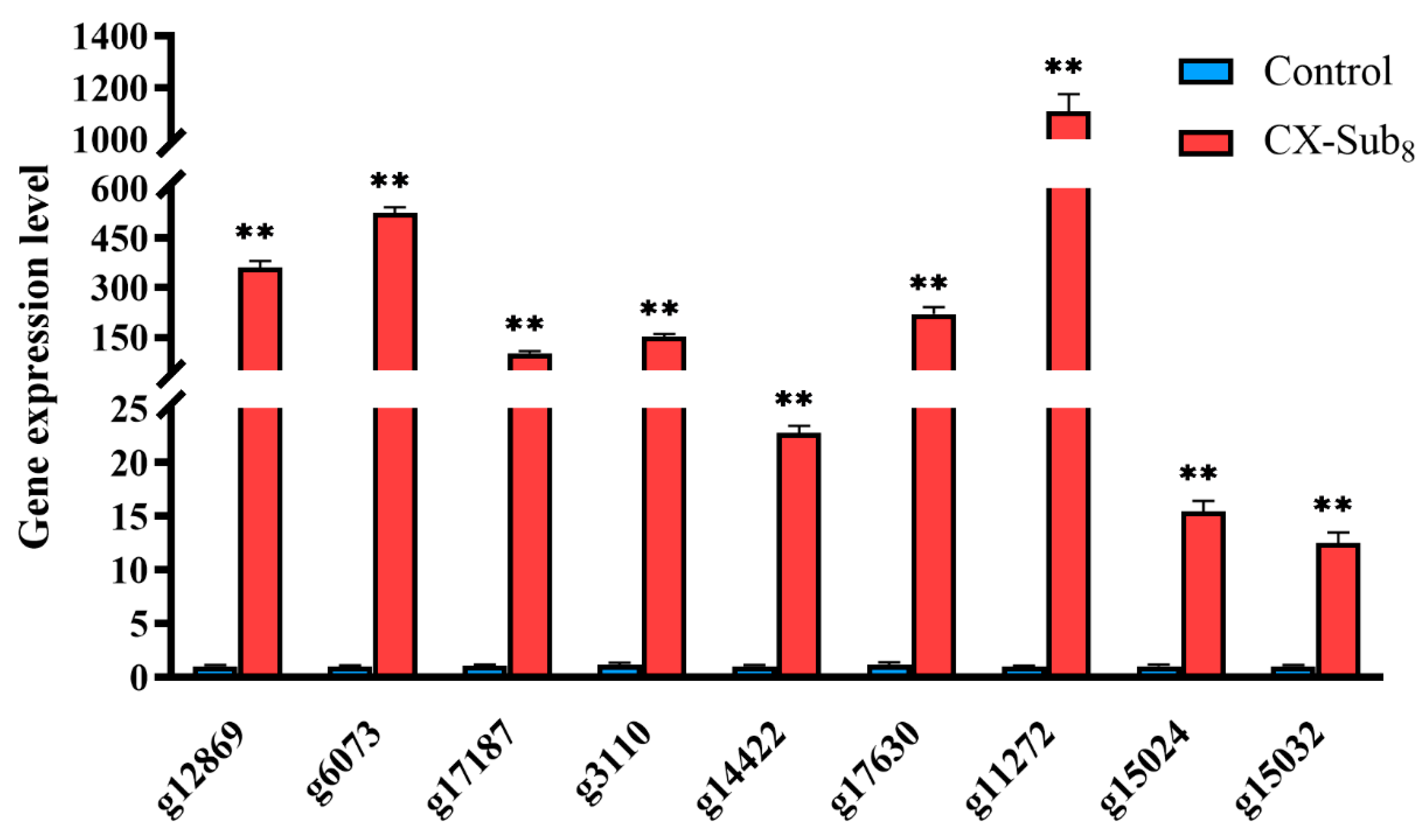
3.6. Disruption of Nutritional Metabolism in P. absoluta by Chlorantraniliprole Revealed Through Transcriptome Sequencing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SE | Standard error |
| ri | Intrinsic rate of increase |
| λ | Finite rate of increase |
| R0 | Net reproductive rate |
| T | Mean generation time |
| GRR | Gross reproductive rate |
| Rf | Relative fitness |
| CX-Sub8 | Chlorantraniliprole-resistant strain continuously selected for eight generations |
| CX-Sub-n | Where n = 1 to n = 8 and represents the serial numbers of generations |
| CX-S | Unselected strains |
| CX-S8 | The eighth generation of the unselected strain, the control strain |
| DEG | Differentially expressed gene |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| APOP | Adult preoviposition period |
| TPOP | Total preoviposition period |
| Nm/Nf | Number of males/Number of females |
References
- Chang, P.E.C.; Metz, M.A. Classification of Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae: Gelechiinae: Gnorimoschemini) based on cladistic analysis of morphology. Proc. Entomol. Soc. Wash. 2021, 123, 41–54. [Google Scholar]
- Desneux, N.; Luna, M.G.; Guillemaud, T.; Urbaneja, A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: The new threat to tomato world production. J. Pest Sci. 2011, 84, 403–408. [Google Scholar] [CrossRef]
- Guillemaud, T.; Blin, A.; Le Goff, I.; Desneux, N.; Reyes, M.; Tabone, E.; Tsagkarakou, A.; Niño, L.; Lombaert, E. The tomato borer, Tuta absoluta, invading the Mediterranean Basin, originates from a single introduction from Central Chile. Sci. Rep. 2015, 5, 8371. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.R.; Biondi, A.; Adiga, A.; Guedes, R.N.; Desneux, N. From the western Palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. J. Pest Sci. 2017, 90, 787–796. [Google Scholar] [CrossRef]
- Zhang, G.F.; Xian, X.Q.; Zhang, Y.B.; Liu, W.X.; Liu, H.; Feng, X.D.; Ma, D.Y.; Wang, Y.S.; Gao, Y.H.; Zhang, R.; et al. Outbreak of the South American tomato leafminer Tuta absoluta on the Chinese mainland: Geographic and potential host range expansion. Pest Manag. Sci. 2021, 77, 5475–5488. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Zhang, Y.; Sun, Z.; Li, Y.; Ding, J.; Zhang, G.; Du, E.; Zi, X.; Tian, C.; et al. Role of Enterococcus mundtii in gut of the tomato leaf miner (Tuta absoluta) to detoxification of Chlorantraniliprole. Pestic. Biochem. Physiol. 2024, 204, 106060. [Google Scholar] [CrossRef]
- Garzia, T.G.; Siscaro, G.; Biondi, A.; Zappala, L. Tuta absoluta, a South American pest of tomato now in the EPPO region: Biology, distribution and damage. Bulletin OEPP. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Siqueira, H.A.A. The tomato borer Tuta absoluta: Insecticide resistance and control failure. CAB Rev. 2012, 7, 1–7. [Google Scholar] [CrossRef]
- Barrientos, R.Z.; Apablaza, H.J.; Norero, S.A.; Estay, P.P. Temperatura base y constante térmica de desarrollo de la polilla del tomate, Tuta absoluta (Lepidoptera: Gelechiidae). Cienc. Investig. Agrar. 1998, 25, 133–137. [Google Scholar] [CrossRef]
- Silva, G.A.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Rosado, J.F.; Guedes, R.N.C. Control failure likelihood and spatial dependence of insecticide resistance in the tomato pinworm, Tuta absoluta. Pest Manag. Sci. 2011, 67, 913–920. [Google Scholar] [CrossRef]
- Gontijo, P.C.; Picanço, M.C.; Pereira, E.J.G.; Martins, J.C.; Chediak, M.; Guedes, R.N.C. Spatial and temporal variation in the control failure likelihood of the tomato leaf miner, Tuta absoluta. Ann. Appl. Biol. 2013, 162, 50–59. [Google Scholar] [CrossRef]
- Grant, C.; Jacobson, R.; Ilias, A.; Berger, M.; Vasakis, E.; Bielza, P.; Zimmer, C.T.; Williamson, M.S.; Ffrench-Constant, R.H.; Vontas, J.; et al. The evolution of multiple-insecticide resistance in UK populations of tomato leafminer, Tuta absoluta. Pest Manag. Sci. 2019, 75, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide resistance in the tomato pinworm Tuta absoluta: Patterns, spread, mechanisms, management and outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Ma, X.; Qu, C.; Yao, J.; Xia, J.; Luo, C.; Guedes, R.N.C.; Wang, R. Resistance monitoring of diamide insecticides and characterization of field-evolved chlorantraniliprole resistance among Chinese populations of the tomato pinworm Phthorimaea (=Tuta) absoluta (Lepidoptera: Gelechiidae). Pestic. Biochem. Physiol. 2024, 205, 106140. [Google Scholar] [CrossRef] [PubMed]
- Sattelle, D.B.; Cordova, D.; Cheek, T.R. Insect ryanodine receptors: Molecular targets for novel pest control chemicals. Invertebr. Neurosci. 2008, 8, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Roditakis, E.; Steinbach, D.; Moritz, G.; Vasakis, E.; Stavrakaki, M.; Ilias, A.; García-Vidal, L.; Martínez-Aguirre, M.D.R.; Bielza, P.; Morou, E.; et al. Ryanodine receptor point mutations confer diamide insecticide resistance in tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae). Insect Biochem. Mol. Biol. 2017, 80, 11–20. [Google Scholar] [CrossRef]
- Siqueira, H.A.A.; Guedes, R.N.C.; Picanço, M.C. Insecticide resistance in populations of Tuta absoluta (Lepidoptera: Gelechiidae). Agric. For. Entomol. 2000, 2, 147–153. [Google Scholar] [CrossRef]
- Roditakis, E.; Skarmoutsou, C.; Staurakaki, M.; Martínez-Aguirre, R.; García-Vidal, L.; Bielza, P.; Haddi, K.; Rapisarda, C.; Rison, J.L.; Bassi, A.; et al. Determination of baseline susceptibility of European populations of Tuta absoluta (Meyrick) to indoxacarb and chlorantraniliprole using a novel dip bioassay method. Pest Manag. Sci. 2013, 69, 217–227. [Google Scholar] [CrossRef]
- Silva, W.M.; Berger, M.; Bass, C.; Balbino, V.Q.; Amaral, M.H.P.; Campos, M.R.; Siqueira, H.A.A. Status of pyrethroid resistance and mechanisms in Brazilian populations of Tuta absoluta. Pestic. Biochem. Physiol. 2015, 122, 8–14. [Google Scholar] [CrossRef]
- Nauen, R.; Bass, C.; Feyereisen, R.; Vontas, J. The role of cytochrome P450s in insect toxicology and resistance. Annu. Rev. Entomol. 2022, 67, 105–124. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Li, X.Y.; Wan, Y.; Yuan, G.; Hussain, S.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Wu, Q. Fitness trade-off associated with Spinosad resistance in Frankliniella occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 2017, 110, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Sanada-Morimura, S.; Matsukura, K.; Van Chien, H.; Cuong, L.Q.; Loc, P.M.; Estoy, G.F.; Matsumura, M. Energy reserve compensating for trade-off between metabolic resistance and life history traits in the brown planthopper (Hemiptera: Delphacidae). J. Econ. Entomol. 2020, 113, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Hardstone, M.C.; Huang, X.; Harrington, L.C.; Harrington, L.C.; Scott, J.G. Differences in development, glycogen, and lipid content associated with cytochrome P450-mediated permethrin resistance in Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2010, 2, 188. [Google Scholar] [CrossRef]
- Nath, A.; Gadratagi, B.G.; Maurya, R.P.; Ullah, F.; Patil, N.B.; Adak, T.; Govindharaj, G.P.P.; Ray, A.; Mahendiran, A.; Desneux, N.; et al. Sublethal phosphine fumigation induces transgenerational hormesis in a factitious host, Corcyra cephalonica. Pest Manag. Sci. 2023, 79, 3548–3558. [Google Scholar] [CrossRef]
- Hafeez, M.; Li, X.; Yousaf, H.K.; Khan, M.M.; Imran, M.; Zhang, Z.; Huang, Y.; Zhang, J.; Shah, S.; Wang, L.; et al. Sublethal effects of bistrifluron on key biological traits, macronutrients contents and vitellogenin (SeVg) expression in Spodoptera exigua (Hübner). Pestic Biochem. Physiol. 2021, 174, 104802. [Google Scholar] [CrossRef]
- Nath, B.S.; Suresh, A.; Varma, B.M.; Kumar, R.P. Changes in protein metabolism in hemolymph and fat body of the silkworm, Bombyx mori (Lepidoptera: Bombycidae) in response to organophosphorus insecticides toxicity. Ecotoxicol. Environ. Saf. 1997, 36, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A. Biochemical response in phosphine resistant and susceptible adult beetles of Trogoderma granarium to the sublethal dose. J. Entomol. Zool. Stud. 2016, 4, 582–587. [Google Scholar]
- Kulma, K.; Saddler, A.; Koella, J.C. Effects of Age and Larval Nutrition on Phenotypic Expression of Insecticide-Resistance in Anopheles Mosquitoes. PLoS ONE 2013, 8, e0058322. [Google Scholar] [CrossRef]
- Freeman, J.C.; Smith, L.B.; Silva, J.J.; Fan, Y.; Sun, H.; Scott, J.G. Fitness studies of insecticide resistant strains: Lessons learned and future directions. Pest Manag. Sci. 2021, 77, 3847–3856. [Google Scholar] [CrossRef]
- Ribeiro, L.M.; Wanderley-Teixeira, V.; Ferreira, H.N.; Teixeira, A.A.; Siqueira, H.A. Fitness costs associated with field-evolved resistance to chlorantraniliprole in Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 2014, 104, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, M.; Liu, S.; Jan, S.; Ali, B.; Shahid, M.; Fernández-Grandon, G.M.; Nawaz, M.; Ahmad, A.; Wang, M. Gossypol-induced fitness gain and increased resistance to deltamethrin in beet armyworm, Spodoptera exigua (Hübner). Pest Manag. Sci. 2019, 75, 683–693. [Google Scholar] [CrossRef]
- Shah, R.M.; Shad, S.A. House fly resistance to chlorantraniliprole: Cross resistance patterns, stability and associated fitness costs. Pest Manag. Sci. 2020, 76, 1866–1873. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Yang, X.B.; Yang, H.; Long, G.Y.; Jin, D.C. Effects of sublethal concentrations of insecticides on the fecundity of Sogatella furcifera (Hemiptera: Delphacidae) via the regulation of vitellogenin and its receptor. J. Insect Sci. 2020, 20, 14. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gong, Y.; Cheng, S.; Desneux, N.; Gao, X.; Xiu, X.; Wang, F.; Hou, M. Transgenerational hormesis effects of nitenpyram on fitness and insecticide tolerance/resistance of Nilaparvata lugens. J. Pest Sci. 2023, 96, 161–180. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Kavousi, A.; Gharekhani, G.; Atlihan, R.; Salih Özgökçe, M.S.; Güncan, A.; Gökçe, A.; Smith, C.L.; Benelli, G.; Guedes, R.N.C.; et al. Advances in theory, data analysis, and application of the age-stage, two-sex life table for demographic research, biological control, and pest management. Entomol. Gen. 2023, 43, 705–732. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Chi, H. Demographic comparison of sweetpotato weevil reared on a major host, Ipomoea batatas, and an alternative host, I. triloba. Sci. Rep. 2015, 5, 11871. [Google Scholar] [CrossRef]
- Abbas, N.; Shad, S.A.; Razaq, M. Fitness cost, cross resistance and realized heritability of resistance to Imidacloprid in Spodoptera litura (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2012, 103, 181–188. [Google Scholar] [CrossRef]
- Nie, Z.; Zuo, L.; Tang, S.; Fang, C.; Ma, Y.; Li, X.; Zhang, J.; Su, J. Effects of Prey Switching at Different Stages on Life Parameters of Neoseiulus bicaudus. Agriculture 2024, 14, 728. [Google Scholar] [CrossRef]
- Biocloud. net. “Transcriptome Analysis Platform for Eukaryotes with Reference Genomes.” n.d. Available online: https://www.biocloud.net/ (accessed on 20 January 2025).
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, Y.; Xu, K.; Wang, Y.; Yang, W. Selection and validation of reference genes for gene expression analysis in Tuta absoluta Meyrick (Lepidoptera: Gelechiidae). Insects 2021, 12, 589. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0; IBM Corp.: Chicago, IL, USA, 2013.
- Silva, J.E.; Ribeiro, L.; Vinasco, N.; Guedes, R.N.C.; Siqueria, H.A.A. Field-evolved resistance to chlorantraniliprole in the tomato pinworm Tuta absoluta: Inheritance, cross-resistance profile, and metabolism. J. Pest Sci. 2019, 92, 1421–1431. [Google Scholar] [CrossRef]
- Jallow, M.F.A.; Dahab, A.A.; Albaho, M.S.; Devi, V.Y.; Awadh, D.G.; Thomas, B.M. Baseline susceptibility and assessment of resistance risk to flubendiamide and chlorantraniliprole in Tuta absoluta (Lepidoptera: Gelechiidae) populations from Kuwait. Appl. Entomol. Zool. 2019, 54, 91–99. [Google Scholar] [CrossRef]
- Gong, W.; Yan, H.H.; Gao, L.; Guo, Y.Y.; Xue, C.B. Chlorantraniliprole resistance in the diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2014, 107, 806–814. [Google Scholar] [CrossRef]
- Troczka, B.J.; Williams, A.J.; Williamson, M.S.; Field, L.M.; Lüemmen, P.; Davies, T.G.E. Stable expression and functional characterisation of the diamondback moth ryanodine receptor G4946E variant conferring resistance to diamide insecticides. Sci. Rep. 2015, 5, 14680. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Bass, C. Does resistance really carry a fitness cost? Curr. Opin. Insect Sci. 2017, 21, 39–46. [Google Scholar] [CrossRef]
- Liu, S.; Yao, X.; Xiang, X.; Yang, Q.; Wang, X.; Xin, T.; Yu, S. Fitness costs associated with chlorantraniliprole resistance in Spodoptera exigua (Lepidoptera: Noctuidae). Pest Manag. Sci. 2021, 77, 1739–1747. [Google Scholar] [CrossRef]
- Yin, X.; Wu, Q.; Li, X.; Zhang, Y.; Xu, B. Demographic Changes in Multigeneration Plutella xylostella (Lepidoptera: Plutellidae) After Exposure to Sublethal Concentrations of Spinosad. J. Econ. Entomol. 2009, 1, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Teixeira, L.A.; Andaloro, J.T. Diamide insecticides: Global efforts to address insect resistance stewardship challenges. Pestic. Biochem. Physiol. 2013, 106, 76–78. [Google Scholar] [CrossRef]
- Erb, S.L.; Bourchier, R.S.; Van Frankenhuyzen, K.; Smith, S.M. Sublethal effects of Bacillus thuringiensis Berliner subsp. kurstaki on Lymantria dispar (Lepidoptera: Lymantriidae) and the Tachinid Parasitoid Compsilura concinnata (Diptera: Tachinidae). Environ. Entomol. 2001, 30, 1174–1181. [Google Scholar] [CrossRef]
- Hannig, G.T.; Ziegler, M.; Marçon, P.G. Feeding cessation effects of chlorantraniliprole, a new anthranilic diamide insecticide, in comparison with several insecticides in distinct chemical classes and mode-of-action groups. Pest Manag. Sci. 2009, 65, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Qu, C.; Yao, J.; Zhan, Q.; Zhang, D.; Li, Y.; Huang, J.; Wang, R. Risk assessment, cross-resistance, biochemical mechanism and fitness cost of tetraniliprole resistance in the tomato pinworm Tuta absoluta. Crop Prot. 2024, 183, 106756. [Google Scholar] [CrossRef]
- Zhang, R.; Dong, J.; Chen, J.; Ji, Q.; Cui, J. The sublethal effects of chlorantraniliprole on Helicoverpa armigera (Lepidoptera: Noctuidae). J. Integr. Agric. 2013, 12, 457–466. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, Z.; Wang, W.; Pan, Y. Effects of sublethal fumigation with phosphine on the reproductive capacity of Liposcelis entomophila (End.) (Psocoptera: Liposcelididae). Int. J. Pest Manag. 2020, 66, 75–81. [Google Scholar] [CrossRef]
- Bozhgani, N.S.; Ghobadi, H.; Riahi, E. Sublethal effects of chlorfenapyr on the life table parameters of two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Syst. Appl. Acarol. 2018, 23, 1342–1351. [Google Scholar] [CrossRef]
- Zalizniak, L.; Nugegoda, D. Effect of sublethal concentrations of chlorpyrifos on three successive generations of Daphnia carinata. Ecotoxicol. Environ. Saf. 2006, 64, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.K.; Collins, P.J. Influence of concentration, temperature and humidity on the toxicity of phosphine to the strongly phosphine-resistant psocid Liposcelis bostrychophila Badonnel (Psocoptera: Liposcelididae). Pest Manag. Sci. 2010, 64, 9. [Google Scholar] [CrossRef]
- Ridley, A.W.; Schlipalius, D.I.; Daglish, G.J. Reproduction of phosphine resistant Rhyzopertha dominica (F.) following sublethal exposure to phosphine. J. Stored Prod. Res. 2012, 48, 106–110. [Google Scholar] [CrossRef]
- Knight, A.L.; Flexner, L. Disruption of mating in codling moth (Lepidoptera: Tortricidae) by chlorantranilipole, an anthranilic diamide insecticide. Pest Manag. Sci. 2007, 63, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, P.C.; Sánchez, N.E. Effect of two solanaceous plants on developmental and population parameters of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Neotrop. Entomol. 2006, 35, 671–676. [Google Scholar] [CrossRef]
- Han, W.; Zhang, S.; Shen, F.; Liu, M.; Ren, C.; Gao, X. Residual toxicity and sublethal effects of chlorantraniliprole on Plutella xylostella (Lepidoptera: Plutellidae). Pest Manag. Sci. 2012, 68, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gong, P.; Li, M.; Qiu, X.; Wang, K. Sublethal effects of Spinosad on survival, growth and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Manag. Sci. 2009, 65, 223–227. [Google Scholar] [CrossRef]
- Dong, J.; Wang, K.; Li, Y.; Wang, S. Lethal and sublethal effects of cyantraniliprole on Helicoverpa assulta (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2017, 136, 58–63. [Google Scholar] [CrossRef]
- Bessin, R.T.; Reagan, T.E. Fecundity of Sugarcane Borer (Lepidoptera: Pyralidae), as affected by larval development on gramineous host plants. Environ. Entomol. 1990, 19, 635–639. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hasegawa, E.; Yamamoto, Y.; Kawatsu, K.; Vargo, E.L.; Yoshimura, J.; Matsuura, K. Sex ratio biases in termites provide evidence for kin selection. Nat. Commun. 2013, 4, 2048. [Google Scholar] [CrossRef]
- Haddi, K.; Mendes, M.V.; Barcellos, M.S.; Lino-Neto, J.; Freitas, H.L.; Guedes, R.N.; Oliveira, E.E. Sexual Success After Stress? Imidacloprid-Induced Hormesis in Males of the Neotropical Stink Bug Euschistus heros. PLoS ONE 2016, 11, e0156616. [Google Scholar] [CrossRef]
- Nozad-Bonab, Z.; Hejazi, M.J.; Iranipour, S.; Arzanlou, M. Lethal and sublethal effects of some chemical and biological insecticides on Tuta absoluta (Lepidoptera: Gelechiidae) eggs and neonates. J. Econ. Entomol. 2017, 110, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lü, L.; He, Y. Effects of insecticides on sex pheromone communication and mating behavior in Trichogramma chilonis. J. Pest Sci. 2018, 91, 65–78. [Google Scholar] [CrossRef]
- Andreazza, F.; Haddi, K.; Nörnberg, S.D.; Guedes, R.N.; Nava, D.E.; Oliveira, E.E. Sex-dependent locomotion and physiological responses shape the insecticidal susceptibility of parasitoid wasps. Environ. Pollut. 2020, 264, 114605. [Google Scholar] [CrossRef]
- Margus, A.; Piiroinen, S.; Lehmann, P.; Tikka, S.; Karvanen, J.; Lindström, L. Sublethal pyrethroid insecticide exposure carries positive fitness effects over generations in a pest insect. Sci. Rep. 2019, 9, 11320. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, C.; Wang, Q.; Wei, Y.; Liu, F.; Xu, S.; Zhang, Z.; Mu, W. Effects of the microbial secondary metabolite benzothiazole on the nutritional physiology and enzyme activities of Bradysia odoriphaga (Diptera: Sciaridae). Pestic. Biochem. Physiol. 2016, 129, 49–55. [Google Scholar] [CrossRef]
- Piri, F.; Sahragard, A.; Ghadamyari, M. Sublethal effects of Spinosad on some biochemical and biological parameters of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Plant Prot. Sci. 2014, 50, 135–144. [Google Scholar] [CrossRef]
- Arambourou, H.; Gismondi, E.; Branchu, P.; Beisel, J.N. Biochemical and morphological responses in Chironomus riparius (Diptera, Chironomidae) larvae exposed to lead-spiked sediment. Environ. Toxicol. Chem. 2013, 32, 2558–2564. [Google Scholar] [CrossRef]
- Etebari, K.; Mirhoseini, S.Z.; Matindoost, L. A study on interaspecific biodiversity of eight groups of silkworm (Bombyx mori) by biochemical markers. Insect Sci. 2005, 12, 87–94. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank ID | NR: Description | E Value | Per. Ident | Direction | Primer Sequences (5′-3′) |
|---|---|---|---|---|---|---|
| g12869 | KAJ2938428.1 | fatty acid desaturase domain-containing protein [Phthorimaea operculella] | 0.0 | 95.43% | F | ATCGGCTGGCTCATGATGAA |
| R | TCGCACACTTGAGGTCTTCT | |||||
| g6073 | KAJ2949132.1 | fatty acid desaturase domain-containing protein [Phthorimaea operculella] | 0.0 | 95.03% | F | CTCACAATGCAGAACGAGGG |
| R | CCAGGTCTCGTTCCAGAAGT | |||||
| g17187 | KAJ2947408.1 | fatty acid desaturase domain-containing protein [Phthorimaea operculella] | 0.0 | 90.82% | F | CAACAGTGCAGCCCATCTTT |
| R | GCAGACAGATTGAACCGGTC | |||||
| g3110 | KAJ2954803.1 | cuticle protein 19.8-like [Melitaea cinxia] | 5 × 10−63 | 56.81% | F | ACGTAGCGGACTCACTTACC |
| R | ATCCTGGCTGACACTACTGC | |||||
| g14422 | KAJ2939703.1 | insect cuticle domain-containing protein [Phthorimaea operculella] | 1 × 10−75 | 76.63% | F | CTCCCACCCCAAATACGAGT |
| R | TGTTGGGGATGTTTGTGTGC | |||||
| g17630 | KAJ2938281.1 | insect cuticle domain-containing protein [Phthorimaea operculella] | 2 × 10−118 | 93.57% | F | CCTGGAGGCTGGTCATACAA |
| R | AATGCGCGTACTTCTGTGTC | |||||
| g11272 | KAJ2949947.1 | GMC oxidoreductase domain-containing protein [Phthorimaea operculella] | 0.0 | 88.66% | F | TGTTTCCGATTTGCCAGTGG |
| R | CATGTAATCGGGGTGCGATG | |||||
| g15024 | KAJ2942836.1 | GMC oxidoreductase domain-containing protein [Phthorimaea operculella] | 0.0 | 93.35% | F | ACGCAACCTAGACATCTCGT |
| R | GTTGAGGACTGTTGATCGCC | |||||
| g15032 | KAJ2942843.1 | GMC oxidoreductase domain-containing protein [Phthorimaea operculella] | 0.0 | 98.10% | F | CTATGTTGGTGGCTGCGATC |
| R | GTTTCATCCCCACCAGCTTC | |||||
| EF1α | MZ054826 | elongation factor 1 α | / | / | F | CCTGGGCACAGAGATTTCAT |
| R | GATCAGCTGCTTGACACCAA | |||||
| RPL28 | MZ054829 | ribosomal protein L28 | / | / | F | TCAGACGTGCTGAACACACA |
| R | GCCAGTCTTGGACAACCATT |
| Generation Number | Population | Slope ± SE | LC25 (95% CL) (mg·L−1) a | LC50 (95% CL) (mg·L−1) | χ2 | df | RR b (with SS) | RR (with CX-Sub1) |
|---|---|---|---|---|---|---|---|---|
| / | SS | 0.465 ± 0.102 | 0.006 (0.000–0.035) | 0.170 (0.026–0.477) | 1.331 | 4 | / | / |
| 1st | CX-Sub1 | 1.213 ± 0.175 | 1.873 (0.709–3.514) | 6.741 (3.609–11.180) | 6.383 | 4 | 39.65 | / |
| 2nd | CX-Sub2 | 1.332 ± 0.178 | 2.160 (1.012–3.627) | 6.933 (4.217–10.649) | 1.482 | 4 | 40.78 | 1.03 |
| 3rd | CX-Sub3 | 0.558 ± 0.110 | 4.734 (1.708–9.306) | 36.504 (19.817–73.235) | 3.419 | 4 | 214.73 | 5.42 |
| 4th | CX-Sub4 | 0.983 ± 0.134 | 6.762 (3.206–11.597) | 32.816 (19.913–56.055) | 2.223 | 4 | 193.04 | 4.87 |
| 5th | CX-Sub5 | 0.923 ± 0.129 | 5.906 (2.639–10.462) | 31.722 (18.793–55.760) | 1.553 | 4 | 186.60 | 4.71 |
| 6th | CX-Sub6 | 0.791 ± 0.121 | 4.871 (1.828–9.392) | 34.708 (19.188–67.062) | 1.425 | 4 | 204.16 | 5.15 |
| 7th | CX-Sub7 | 0.844 ± 0.124 | 5.699 (2.346–10.536) | 35.878 (20.458–66.788) | 0.819 | 4 | 211.05 | 5.32 |
| 8th | CX-Sub8 | 0.947 ± 0.132 | 7.426 (3.455–12.895) | 38.313 (22.937–67.422) | 2.144 | 4 | 225.37 | 5.68 |
| Stage | Control (Mean ± SE) | CX-Sub2 (Mean ± SE) | CX-Sub4 (Mean ± SE) | CX-Sub8 (Mean ± SE) |
|---|---|---|---|---|
| Egg (days) | 4.87 ± 0.05 bc | 4.77 ± 0.06 c | 4.98 ± 0.04 b | 5.52 ± 0.06 a |
| Larvae 1 (days) | 3.02 ± 0.07 c | 2.88 ± 0.07 c | 3.28 ± 0.08 b | 3.59 ± 0.06 a |
| Larvae 2 (days) | 2.81 ± 0.07 b | 2.76 ± 0.09 b | 3.22 ± 0.08 a | 3.28 ± 0.06 a |
| Larvae 3 (days) | 2.72 ± 0.07 c | 2.80 ± 0.07 b | 3.10 ± 0.08 a | 3.14 ± 0.08 a |
| Larvae 4 (days) | 2.77 ± 0.07 a | 2.63 ± 0.08 a | 2.52 ± 0.07 b | 2.71 ± 0.09 a |
| Pupae (days) | 7.14 ± 0.09 bc | 6.94 ± 0.08 c | 7.25 ± 0.12 ab | 7.58 ± 0.11 a |
| Preadult (days) | 23.26 ± 0.13 c | 22.93 ± 0.17 c | 24.24 ± 0.23 b | 25.71 ± 0.21 a |
| Adult (days) | 17.53 ± 0.66 b | 17.54 ± 0.73 b | 19.03 ± 0.61 ab | 20.06 ± 0.72 a |
| Female adult longevity (days) | 18.09 ± 0.92 b | 16.80 ± 0.99 b | 18.79 ± 0.78 ab | 21.12 ± 1.13 a |
| Male adult longevity (days) | 17.07 ± 0.93 a | 18.27 ± 1.00 a | 19.24 ± 0.93 a | 18.77 ± 0.74 a |
| APOP (days) | 2.12 ± 0.14 b | 3.03 ± 0.14 a | 3.11 ± 0.15 a | 2.68 ± 0.19 a |
| TPOP (days) | 25.12 ± 0.25 d | 26.00 ± 0.30 c | 27.34 ± 0.31 b | 28.45 ± 0.32 a |
| Oviposition (days) | 5.79 ± 0.17 a | 5.23 ± 0.25 a | 4.24 ± 0.19 b | 4.13 ± 0.21 b |
| Fecundity (eggs/♀) | 102.18 ± 4.68 a | 71.06 ± 7.00 b | 57.63 ± 4.81 bc | 48.56 ± 6.30 c |
| Life Table Parameters | Control (Mean ± SE) | CX-Sub2 (Mean ± SE) | CX-Sub4 (Mean ± SE) | CX-Sub8 (Mean ± SE) |
|---|---|---|---|---|
| GRR (offspring individual−1) | 47.14 ± 6.40 a | 32.56 ± 5.10 ab | 28.37 ± 3.99 b | 27.68 ± 4.50 b |
| R0 (offspring individual−1) | 33.72 ± 5.05 a | 24.87 ± 4.17 ab | 21.90 ± 3.32 b | 20.88 ± 3.60 b |
| ri (day−1) | 0.13 ± 0.01 a | 0.11 ± 0.01 ab | 0.11 ± 0.01 b | 0.10 ± 0.01 b |
| T (days) | 27.82 ± 0.23 b | 28.17 ± 0.29 b | 28.58 ± 0.32 b | 30.17 ± 0.42 a |
| λ(day−1) | 1.14 ± 0.01 a | 1.12 ± 0.01 ab | 1.11 ± 0.01 b | 1.10 ± 0.01 b |
| Rf | 1.00 | 0.74 | 0.65 | 0.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Jia, Z.; Fu, K.; Ding, X.; Jiang, W.; Wang, X.; Ahmat, T.; Wu, J.; Wen, Y.; Ye, X.; et al. Multigeneration Sublethal Chlorantraniliprole Treatment Disrupts Nutritional Metabolism and Inhibits Growth, Development, and Reproduction of Phthorimaea absoluta. Insects 2025, 16, 524. https://doi.org/10.3390/insects16050524
Li L, Jia Z, Fu K, Ding X, Jiang W, Wang X, Ahmat T, Wu J, Wen Y, Ye X, et al. Multigeneration Sublethal Chlorantraniliprole Treatment Disrupts Nutritional Metabolism and Inhibits Growth, Development, and Reproduction of Phthorimaea absoluta. Insects. 2025; 16(5):524. https://doi.org/10.3390/insects16050524
Chicago/Turabian StyleLi, Lun, Zunzun Jia, Kaiyun Fu, Xinhua Ding, Weihua Jiang, Xiaowu Wang, Tursun. Ahmat, Jiahe Wu, Yutong Wen, Xiaoqin Ye, and et al. 2025. "Multigeneration Sublethal Chlorantraniliprole Treatment Disrupts Nutritional Metabolism and Inhibits Growth, Development, and Reproduction of Phthorimaea absoluta" Insects 16, no. 5: 524. https://doi.org/10.3390/insects16050524
APA StyleLi, L., Jia, Z., Fu, K., Ding, X., Jiang, W., Wang, X., Ahmat, T., Wu, J., Wen, Y., Ye, X., Guo, W., & Hu, H. (2025). Multigeneration Sublethal Chlorantraniliprole Treatment Disrupts Nutritional Metabolism and Inhibits Growth, Development, and Reproduction of Phthorimaea absoluta. Insects, 16(5), 524. https://doi.org/10.3390/insects16050524