
Assessing Wine Grape Cultivar Susceptibility to Spotted Wing Drosophila and Melanogaster-Type Drosophila in Hungarian Vineyards: Effects of Berry Integrity and Insights into Larval Interactions


 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location and Cultivars
2.2. Bottle Trap Survey
2.3. Intact Berry Treatment
2.4. Hurt Berry Treatment
2.5. Statistical Analysis
3. Results
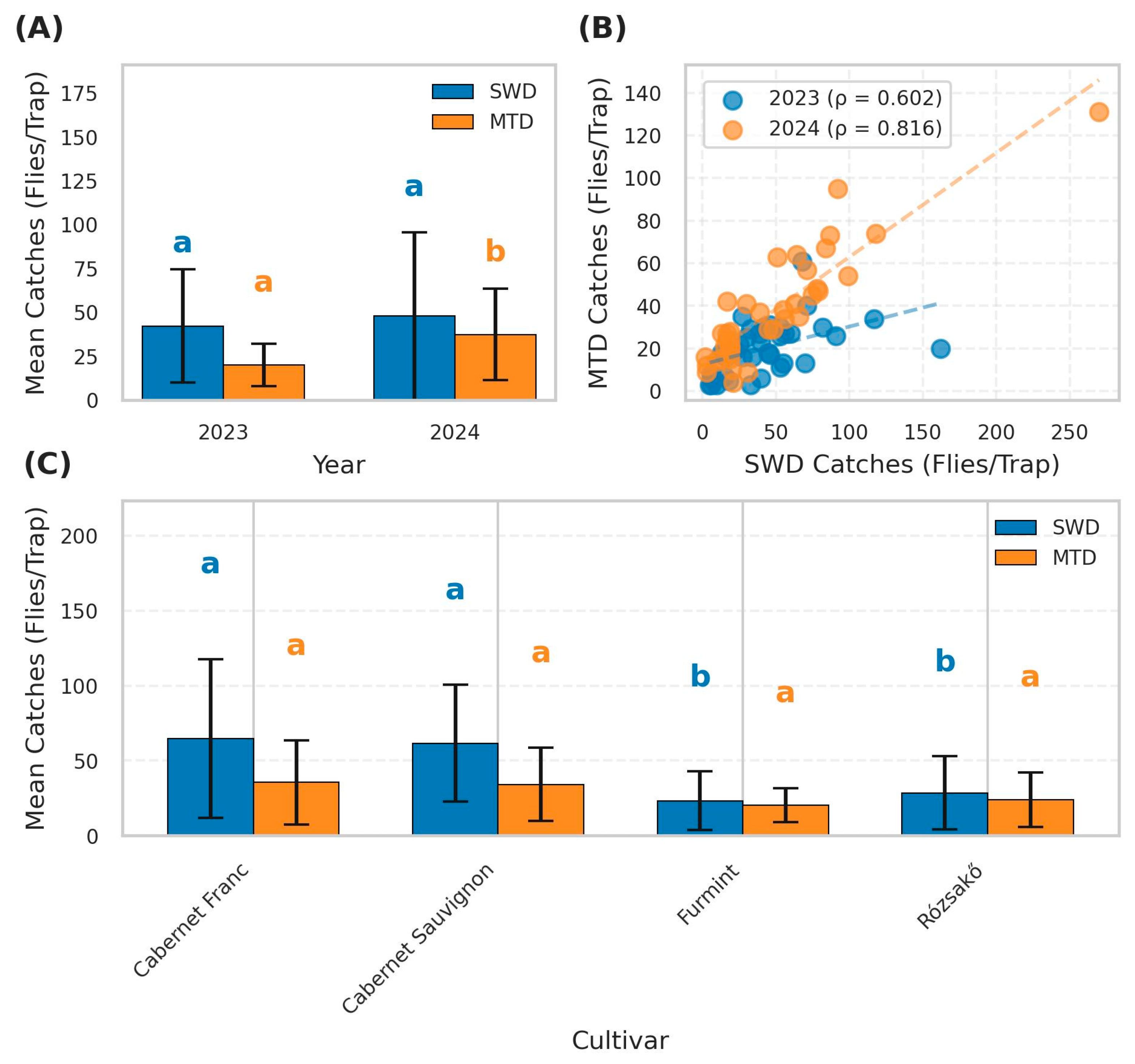
3.1. Bottle Trap Catches
3.2. Fly Emergence from Unhurt Single Berries
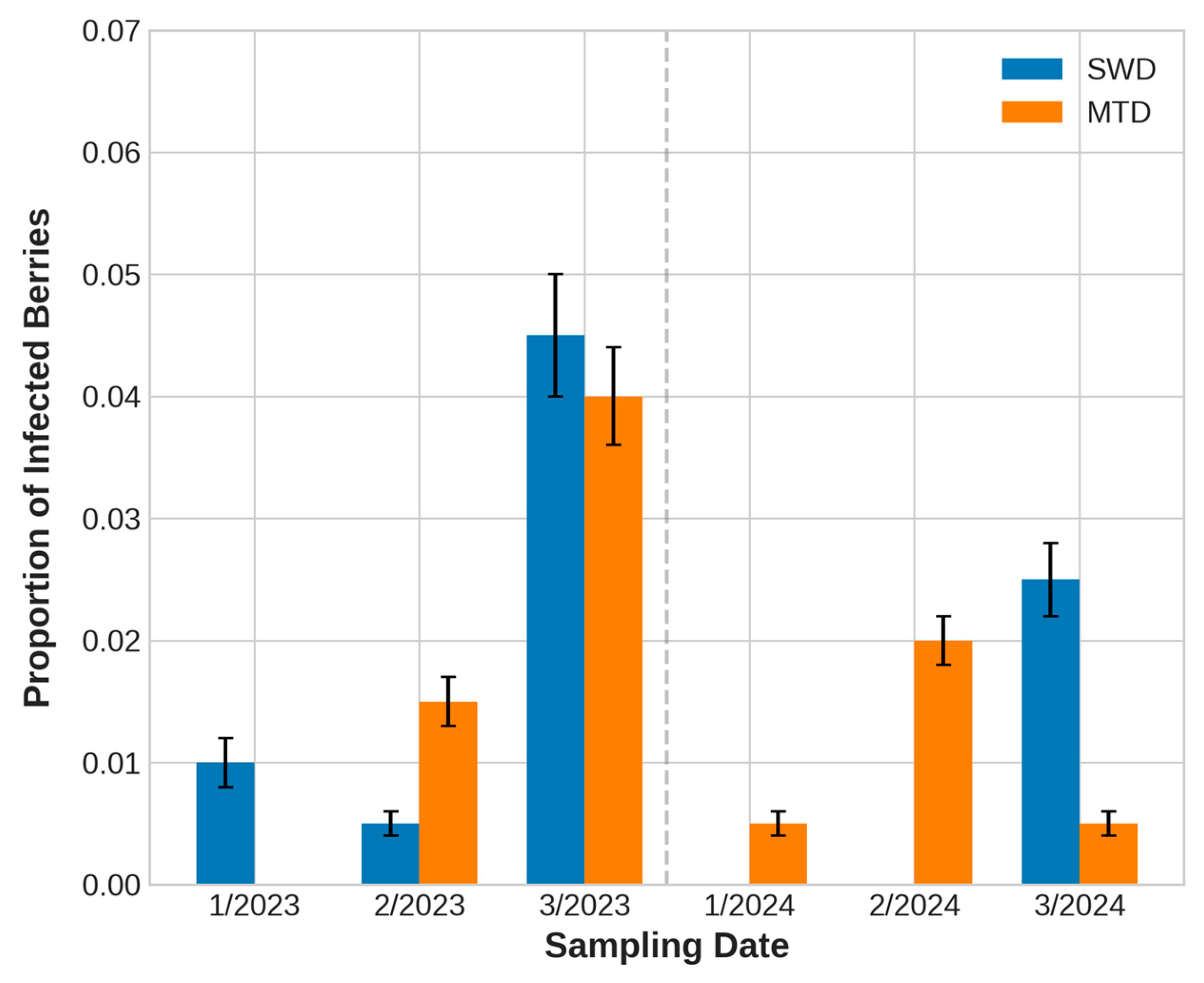
3.2.1. Proportion of Infested Berries by Sampling Dates
3.2.2. Varietal Differences in Infestation Rates
3.3. Fly Emergence from Unhurt Pooled Berries (Varietal Differences)
3.4. Comparison of Infestation of Intact and Hurt Single Berries
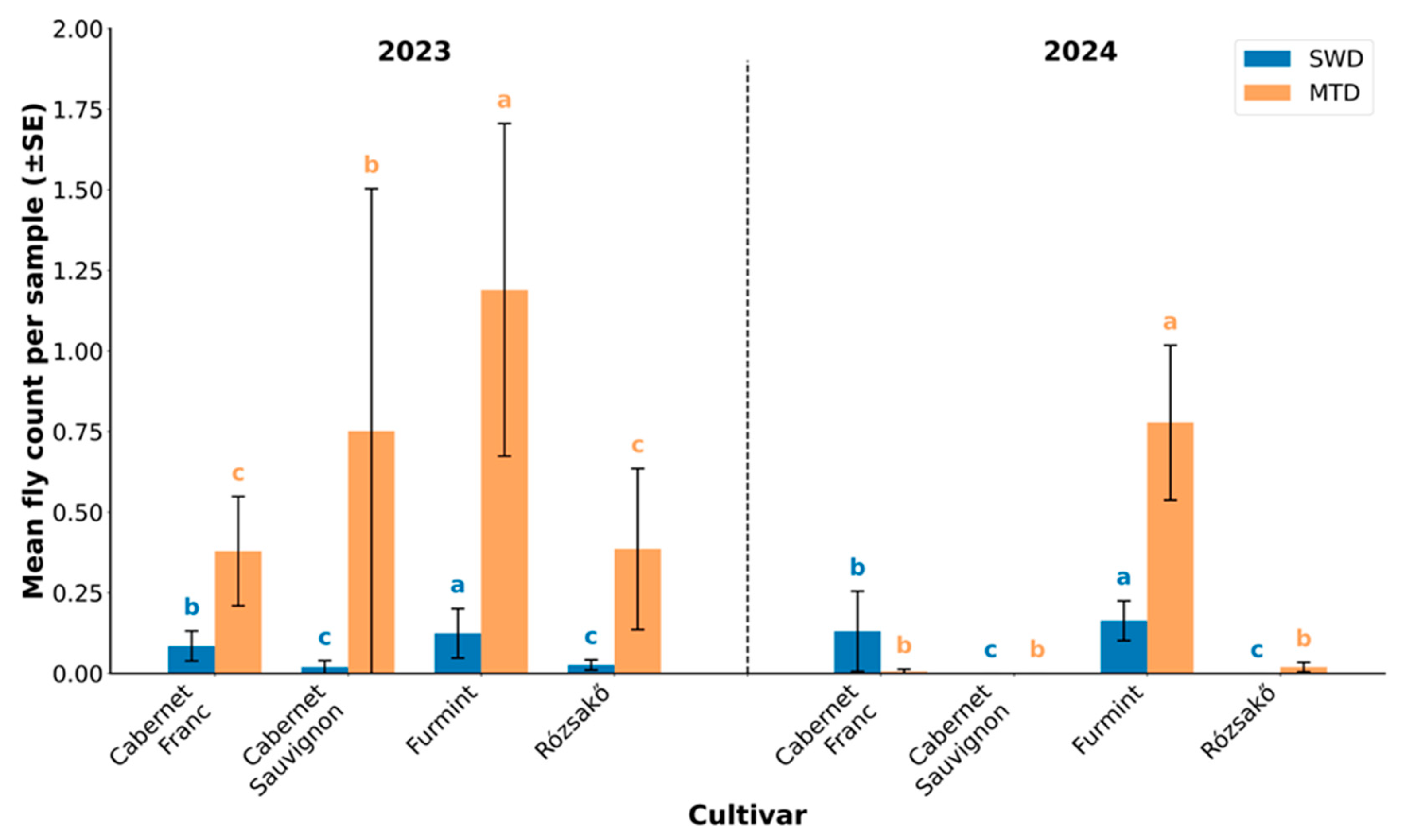
3.5. Differences in Drosophila Composition in Isolated and Pooled Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Matsumura, S. 6000 Illustrated Insects of Japan–Empire; The Toko-Shoin: Tokyo, Japan, 1931; p. 1689. [Google Scholar]
- Kaneshiro, K.Y. Drosophila (Sophophora) suzukii (Matsumura). Proc. Hawaii. Entomol. Soc. 1983, 24, 179. [Google Scholar]
- Hauser, M. A historic account of the invasion of Drosophila suzukii (Matsumura) in the continental United States, with remarks on identification. Pest Manag. Sci. 2011, 67, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Ometto, L.; Cestaro, A.; Ramasamy, S.; Grassi, A.; Revadi, S.; Siozios, S.; Moretto, M.; Fontana, P.; Varotto, C.; Pisani, D.; et al. Linking genomics and ecology to investigate the complex evolution of an invasive drosophilid pest. Genome Biol. Evol. 2013, 5, 745–757. [Google Scholar] [CrossRef]
- EPPO. EPPO Global Database—Drosophila suzukii. Available online: https://gd.eppo.int/taxon/DROSSU (accessed on 10 January 2025).
- Deutsch, F.; Kiss, B. Seasonal Abundance Changes of Spotted Wing Drosophila in Neighbouring Habitats in Hungary. In Proceedings of the 1st International Electronic Conference on Entomology, Virtual Event, 1–15 July 2021; MDPI: Basel, Switzerland, 2021. [Google Scholar]
- Pelton, E.; Gratton, C.; Guédot, C. Susceptibility of cold-hardy grapes to Drosophila suzukii (Diptera: Drosophilidae). J. Appl. Entomol. 2017, 141, 644–652. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the Asiatic fruit fly Drosophila suzukii: The key to its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef]
- Ioriatti, C.; Walton, V.M.; Dalton, D.T.; Anfora, G.; Grassi, A.; Maistri, S.; Mazzoni, V. Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. J. Econ. Entomol. 2015, 108, 1148–1155. [Google Scholar] [CrossRef]
- Entling, W.; Hoffmann, C. Single and combined effects of Drosophila suzukii and Drosophila melanogaster on sour rot development in viticulture. J. Appl. Entomol. 2019, 143, 592–601. [Google Scholar] [CrossRef]
- Rombaut, A.; Guilbaud, R.; Xuéreb, A.; Benoit, L.; Chapuis, M.P.; Gibert, P.; Fellous, S. Invasive Drosophila suzukii facilitates Drosophila melanogaster infestation and sour rot outbreaks in vineyards. R. Soc. Open Sci. 2017, 4, 170181. [Google Scholar] [CrossRef]
- Linder, C.; Martin, C.; Laboisse, S.; Chatelain, P.G.; Kehrli, P. Susceptibility of various grape cultivars to Drosophila suzukii and other vinegar flies. IOBC-WPRS Bull. 2014, 105, 219–224. [Google Scholar]
- Baser, N.; Broutou, O.; Verrastro, V.; Porcelli, F.; Ioriatti, C.; Anfora, G.; Mazzoni, V.; Stacconi, M.V.R. Susceptibility of table grape varieties grown in south-eastern Italy to Drosophila suzukii. J. Appl. Entomol. 2018, 142, 465–472. [Google Scholar] [CrossRef]
- Knapp, L.; Mazzi, D.; Finger, R. The economic impact of Drosophila suzukii: Perceived costs and revenue losses of Swiss fruit growers. Pest Manag. Sci. 2020, 76, 3434–3445. [Google Scholar]
- Deans, C.A.; Hutchison, W. The Protein Paradox: Elucidating the Complex Nutritional Ecology of the Invasive Berry Pest, Spotted-Wing Drosophila (Diptera: Drosophila suzukii). Front. Insect Sci. 2021, 1, 787169. [Google Scholar] [CrossRef] [PubMed]
- Nagy, G.M.; Varga, J.; Dénes, F. Drosophila suzukii (Matsumura) fertőzés terjedése és kártétele 2016-ban Magyarországon [Invasion and damage of Drosophila suzukii (Matsumura) in 2016 in Hungary]. In Studies in Honour of Károly Mészáros; Szabó, P., Ed.; University of Sopron: Sopron, Hungary, 2017; pp. 37–42. (In Hungarian) [Google Scholar]
- Kiss, B.; Lengyel, G.D.; Nagy, Z.; Kárpáti, Z. A pettyesszárnyú muslica (Drosophila suzukii) első magyarországi előfordulása [First record of spotted wing drosophila in Hungary]. Növényvédelem 2013, 49, 97–99. (In Hungarian) [Google Scholar]
- Soto-Yéber, L.; Soto-Ortiz, J.; Godoy, P.; Godoy-Herrera, R. The behavior of adult Drosophila in the wild. PLoS ONE 2018, 13, e0209917. [Google Scholar] [CrossRef]
- Robinson, J.; Harding, J.; Vouillamoz, J. Wine Grapes: A Complete Guide to 1368 Vine Varieties, Including Their Origins and Flavors, 1st ed.; HarperCollins: New York, NY, USA, 2012; p. 1280. [Google Scholar]
- Boulton, R.B.; Singleton, V.L.; Bisson, L.F.; Kunkee, R.E. Principles and Practices of Winemaking; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar] [CrossRef]
- Jackson, R.S. Wine Science: Principles and Applications, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology: The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons: Chichester, UK, 2006; Volume 1, p. 497. [Google Scholar]
- Kállay, M.; Hajdu, E. Furmint és a Tokaji Borok Kémiai Összetétele [Furmint and the Chemical Composition of Tokaji Wines]; Pécsi Tudományegyetem: Pécs, Hungary, 2007. (In Hungarian) [Google Scholar]
- Kozma, P.; Karácsony, Z.; Bisztray, G. Szőlőfajták Magyarországon [Grape Varieties in Hungary]; Mezőgazda Kiadó: Budapest, Hungary, 2005. (In Hungarian) [Google Scholar]
- Parsons, P.A. Phototactic responses along a gradient of light intensities for the sibling species Drosophila melanogaster and Drosophila simulans. Behav. Genet. 1975, 5, 17–25. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted wing Drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A.; et al. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. 2018, 91, 1213–1224. [Google Scholar] [CrossRef]
- Aguiar, A.A.R.M.; Neto, J.; Sousa, P.A.S.; Roque, V.; Chichorro, L. Off-Crop and Off-Season Monitoring, Key Elements to Be Integrated into an Effective Strategy for the Control of Drosophila suzukii (Diptera: Drosophilidae). Agronomy 2024, 14, 2714. [Google Scholar] [CrossRef]
- Arnold, G. A Drosophila suzukii és a szőlő kapcsolata [The Relationship Between Drosophila suzukii and Grapes]. Master’s Thesis, Szent István University, Budapest, Hungary, 2018. (In Hungarian). [Google Scholar]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Weißinger, L.; Samuel, N.; Breuer, M.; Müller, C. Effects of variety and grape berry condition of Vitis vinifera on preference behavior and performance of Drosophila suzukii. Insects 2019, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Marcellin-Gros, R.; Hévin, S.; Chevalley, C.; Boccard, J.; Hofstetter, V.; Gindro, K.; Wolfender, J.-L.; Kehrli, P. An advanced metabolomic approach on grape skins untangles cultivar preferences by Drosophila suzukii for oviposition. Front. Plant Sci. 2024, 15, 1435943. [Google Scholar] [CrossRef]
- Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Belien, T.; Bylemans, D. Olfactory Preference of Drosophila suzukii Shifts Between Fruit and Fermentation Cues over the Season: Effects of Physiological Status. Insects 2019, 10, 200. [Google Scholar] [CrossRef]
- Revadi, S.; Vitagliano, S.; Rossi Stacconi, M.V.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory Responses of Drosophila suzukii Females to Host Plant Volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Swoboda-Bhattarai, K.A.; McPhie, D.R.; Burrack, H.J. Reproductive Status of Drosophila suzukii (Diptera: Drosophilidae) Females Influences Attraction to Fermentation-Based Baits and Ripe Fruits. J. Econ. Entomol. 2017, 110, 1648–1652. [Google Scholar] [CrossRef]
- Ebbenga, D.N.; Burkness, E.C.; Clark, M.D.; Hutchison, W.D. Risk of spotted-wing Drosophila injury and associated increases in acetic acid in Minnesota wine grapes. Am. J. Enol. Vitic. 2021, 72, 106–112. [Google Scholar] [CrossRef]
- Weißinger, L.; Arand, K.; Bieler, E.; Kassemeyer, H.-H.; Breuer, M.; Müller, C. Physical and chemical traits of grape varieties influence Drosophila suzukii preferences and performance. Front. Plant Sci. 2021, 12, 664636. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Brewer, L.J.; Dalton, D.T.; Lee, J.C.; Nieri, R.; Park, K.R.; Pfab, F.; Tait, G.; Walton, V. Host Range and Characteristics Affecting Fruit Susceptibility to Spotted-Wing Drosophila; Oregon State University Extension Service: Washington, DC, USA, 2019. [Google Scholar]
- Tonina, L.; Giomi, F.; Sancassani, M.; Ajelli, M.; Mori, N.; Giongo, L. Texture features explain the susceptibility of grapevine cultivars to Drosophila suzukii (Diptera: Drosophilidae) infestation in ripening and drying grapes. Sci. Rep. 2020, 10, 10245. [Google Scholar] [CrossRef]
- Hajdu, E. Viticulture of Hungary. Acta Agrar. Debreceniensis 2018, 150, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Shrader, M.E.; Burrack, H.J.; Pfeiffer, D.G. Drosophila suzukii (Diptera: Drosophilidae) oviposition and adult emergence in six wine grape varieties grown in Virginia. J. Econ. Entomol. 2019, 112, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Farkas, E.A.; Jahnke, G.; Szőke, B.; Deák, T.; Oláh, R.; Oláh, K.; Szigeti, G.K.; Németh, C.; Sárdy, D.Á.N. Clonal selection of autochthonous grape varieties in Badacsony, Hungary. Horticulturae 2023, 9, 994. [Google Scholar] [CrossRef]
- Shaw, B.; Brain, P.; Wijnen, H.; Fountain, M.T. Reducing Drosophila suzukii emergence through inter-species competition. Pest Manag. Sci. 2018, 74, 1466–1471. [Google Scholar] [CrossRef]
- Rombaut, A.; Gallet, R.; Qitout, K.; Samy, M.; Guilhot, R.; Ghirardini, P.; Lazzaro, B.P.; Becher, P.G.; Xuéreb, A.; Gibert, P.; et al. Microbiota-mediated competition between Drosophila species. Microbiome 2023, 11, 201. [Google Scholar] [CrossRef]
- Kidera, H.; Takahashi, K.H. Chemical cues from competitors change the oviposition preference of Drosophila suzukii. Entomol. Exp. Appl. 2020, 168, 304–310. [Google Scholar] [CrossRef]
- Leach, H.; Van Timmeren, S.; Wetzel, W.; Isaacs, R. Predicting Within- and Between-Year Variation in Activity of the Invasive Spotted Wing Drosophila (Diptera: Drosophilidae) in a Temperate Region. Environ. Entomol. 2019, 48, 1223–1233. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, J.S.; Park, J.S.; Choi, D.-S.; Park, J.; Kim, I. Oviposition and development potential of the spotted-wing Drosophila, Drosophila suzukii (Diptera: Drosophilidae), on uninjured Campbell Early grape. Entomol. Res. 2015, 45, 354–359. [Google Scholar] [CrossRef]
- Entling, W.; Anslinger, S.; Jarausch, B.; Michl, G.; Hoffmann, C. Berry skin resistance explains oviposition preferences of Drosophila suzukii at the level of grape cultivars and single berries. J. Pest Sci. 2019, 92, 477–484. [Google Scholar] [CrossRef]
- Stemberger, T.L. Survey of hanging and fallen cherry fruit use by spotted wing Drosophila, Drosophila suzukii (Matsumura, 1931) (Diptera: Drosophilidae), and other Drosophilidae species. Pan-Pac. Entomol. 2016, 91, 347–351. [Google Scholar] [CrossRef]
- Holle, S.G.; Burkness, E.C.; Cira, T.M.; Hutchison, W.D. Influence of previous fruit injury on susceptibility to spotted wing Drosophila (Diptera: Drosophilidae) infestation in the Midwestern United States. J. Entomol. Sci. 2017, 52, 207–215. [Google Scholar] [CrossRef]
- Becher, P.G.; Flick, G.; Rozpędowska, E.; Schmidt, A.; Hagman, A.; Lebreton, S.; Larsson, M.C.; Hansson, B.S.; Piškur, J.; Witzgall, P.; et al. Yeast, not fruit volatiles, mediate Drosophila melanogaster attraction, oviposition, and development. Funct. Ecol. 2012, 26, 822–828. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | SWD 2023 (%) | SWD 2024 (%) | MTD 2023 (%) | MTD 2024 (%) |
|---|---|---|---|---|
| Cabernet Franc | 1.3 | 0.0 | 0.7 | 0.0 |
| Cabernet Sauvignon | 3.3 | 0.0 | 0.0 | 0.0 |
| Furmint | 4.7 | 4.0 | 7.3 | 4.7 |
| Rózsakő | 0.0 | 0.0 | 1.3 | 0.0 |
| Year | Condition | Non- Infested (%) | MTD (%) | SWD (%) | SWD + MTD (%) | Total (N) |
|---|---|---|---|---|---|---|
| 2023 | Intact | 96.33 | 1.67 | 1.83 | 0.17 | 600 |
| 2023 | Hurt | 54.0 | 32.25 | 1.75 | 12.0 | 400 |
| 2024 | Intact | 98.17 | 1.0 | 0.83 | 0 | 600 |
| 2024 | Hurt | 78.0 | 16.0 | 5.25 | 0.75 | 400 |
| Year | Condition | MTD (%) | SWD (%) | Total (N) |
|---|---|---|---|---|
| 2023 | Intact Single Berry | 50.0 | 50.0 | 28 |
| Intact Pooled Berries | 91.4 | 8.6 | 453 | |
| Injured Single Berry | 90.5 | 9.5 | 1037 | |
| Injured Pooled Berries | 97.0 | 3.0 | 2074 | |
| 2024 | Intact Single Berry | 53.8 | 46.2 | 13 |
| Intact Pooled Berries | 73.2 | 26.8 | 168 | |
| Injured Single Berry | 84.8 | 15.2 | 243 | |
| Injured Pooled Berries | 90.3 | 9.7 | 1813 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibn Amor, A.; Kukorellyné Szénási, Á.; Németh, C.; Deutsch, F.; Kiss, B. Assessing Wine Grape Cultivar Susceptibility to Spotted Wing Drosophila and Melanogaster-Type Drosophila in Hungarian Vineyards: Effects of Berry Integrity and Insights into Larval Interactions. Insects 2025, 16, 497. https://doi.org/10.3390/insects16050497
Ibn Amor A, Kukorellyné Szénási Á, Németh C, Deutsch F, Kiss B. Assessing Wine Grape Cultivar Susceptibility to Spotted Wing Drosophila and Melanogaster-Type Drosophila in Hungarian Vineyards: Effects of Berry Integrity and Insights into Larval Interactions. Insects. 2025; 16(5):497. https://doi.org/10.3390/insects16050497
Chicago/Turabian StyleIbn Amor, Abir, Ágnes Kukorellyné Szénási, Csaba Németh, Ferenc Deutsch, and Balázs Kiss. 2025. "Assessing Wine Grape Cultivar Susceptibility to Spotted Wing Drosophila and Melanogaster-Type Drosophila in Hungarian Vineyards: Effects of Berry Integrity and Insights into Larval Interactions" Insects 16, no. 5: 497. https://doi.org/10.3390/insects16050497
APA StyleIbn Amor, A., Kukorellyné Szénási, Á., Németh, C., Deutsch, F., & Kiss, B. (2025). Assessing Wine Grape Cultivar Susceptibility to Spotted Wing Drosophila and Melanogaster-Type Drosophila in Hungarian Vineyards: Effects of Berry Integrity and Insights into Larval Interactions. Insects, 16(5), 497. https://doi.org/10.3390/insects16050497