
Field-Based Evaluation of Insecticide Effectiveness on Megalurothrips usitatus in Guangdong, China: Implications for Pest Control Strategies
 , , and
, , and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Populations
2.2. Insecticides
2.3. Bioassays for Insecticide Toxicity
2.4. Field Experiment
2.5. Data Analysis
3. Results
3.1. Morphological Characteristics of M. usitatus
3.2. Toxicity of 10 Insecticides Against QY Population
3.3. Toxicity of 10 Insecticides Against YF Population
3.4. Toxicity of Tested Insecticides Against MM Population
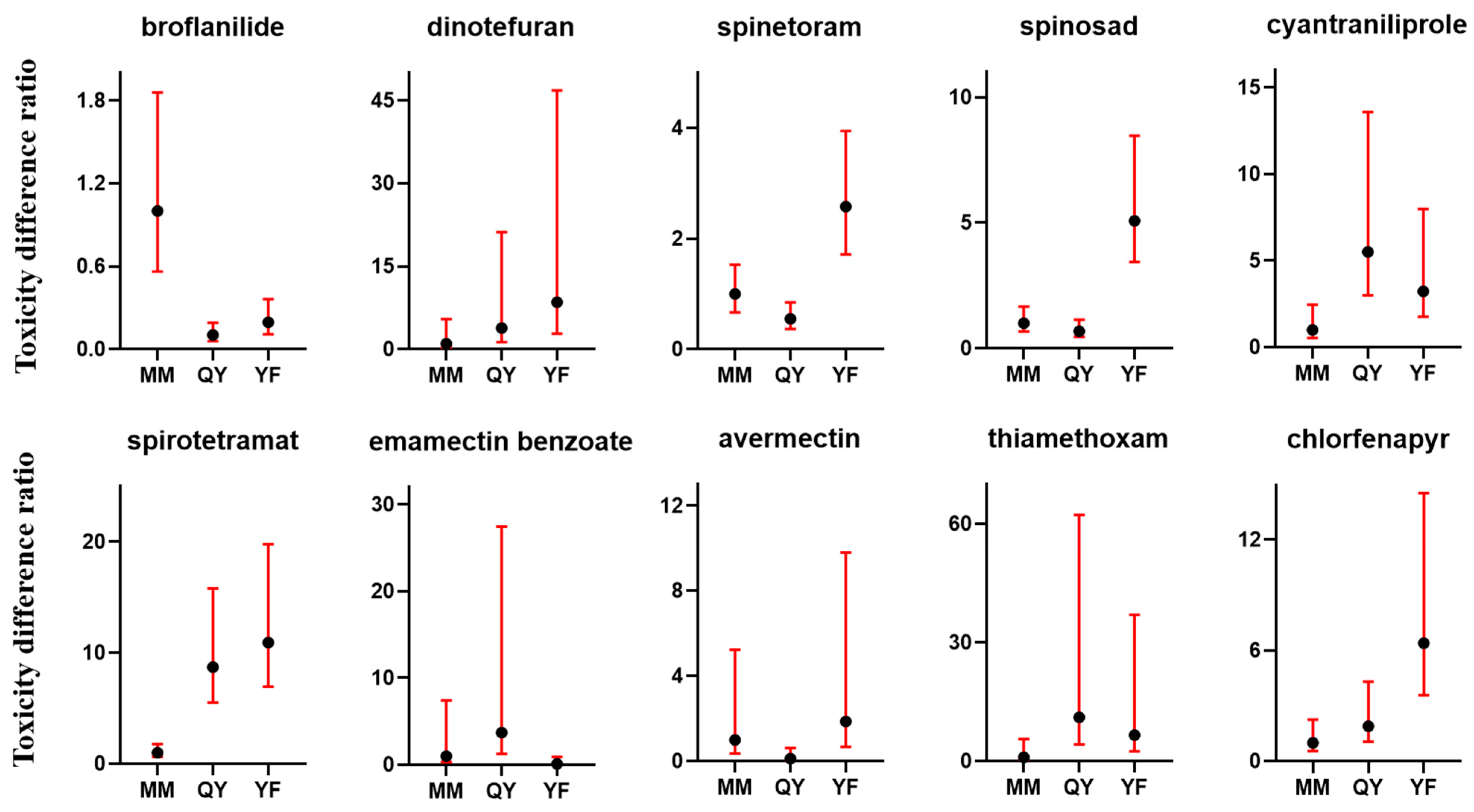
3.5. Variances in Toxicity of Insecticides Against Different Field Populations
3.6. Resistance of M. usitatus to Six Insecticides
3.7. Field Efficacy Trial of Five Selected Insecticides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.H.; Shu, S.Q.; Wanamaker, S.I.; Lo, S.; Tanskanen, J.; Schulman, A.H.; Zhu, T.T.; Luo, M.C. The genome of cowpea (Vigna unguiculata [L.] Walp.). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Herniter, I.A.; Muñoz-Amatriaín, M.; Close, T.J. Genetic, textual, and archeological evidence of the historical global spread of cowpea (Vigna unguiculata [L.] Walp.). Legume Sci. 2020, 2, e57. [Google Scholar] [CrossRef]
- Liang, Q.; Muñoz-Amatriaín, M.; Shu, S.; Lo, S.; Wu, X.; Carlson, J.W.; Davidson, P.; Goodstein, D.M.; Phillips, J.; Janis, N.M. A view of the pan-genome of domesticated Cowpea (Vigna unguiculata [L.] Walp.). Plant Genome 2024, 17, e20319. [Google Scholar] [CrossRef] [PubMed]
- Huan, Z.; Xu, Z.; Luo, J.; Xie, D. Monitoring and exposure assessment of pesticide residues in cowpea (Vigna unguiculata L. Walp) from five provinces of southern China. Regul. Toxicol. Pharmacol. 2016, 81, 260–267. [Google Scholar] [CrossRef]
- Xiao, L.; Li, Y.; Lan, P.; Tan, G.; Ding, M.; Li, R.; Li, F. First report of Tomato spotted wilt virus infecting cowpea in China. Plant Dis. 2016, 100, 233. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Z.; Wu, Z.; Li, T.; Shen, Z.; Zhou, X.; Wu, X.; Li, G.; Zhang, Y. A near-complete assembly of asparagus bean provides insights into anthocyanin accumulation in pods. Plant Biotechnol. J. 2023, 21, 2473–2489. [Google Scholar] [CrossRef]
- Yu, H.; Wu, M.; Li, S.; Li, J.; Zou, X.; Guo, Z.; Wu, Q.; Zhang, Y.; Kong, X.; Xie, W. A Maximum Dose Bioassay to Assess Efficacy of Spinetoram against Cowpea Thrip Megalurothrips usitatus in China. Insects 2024, 15, 412. [Google Scholar] [CrossRef]
- Tang, L.D.; Guo, L.H.; Wu, J.H.; Zang, L.S. Thrips in genus Megalurothrips (Thysanoptera: Thripidae): Biodiversity, bioecology, and IPM. J. Integr. Pest Manag. 2023, 14, 8. [Google Scholar] [CrossRef]
- Liu, P.; Qin, Z.; Feng, M.; Zhang, L.; Huang, X.; Shi, W. The male-produced aggregation pheromone of the bean flower thrips Megalurothrips usitatus in China: Identification and attraction of conspecifics in the laboratory and field. Pest Manag. Sci. 2020, 76, 2986–2993. [Google Scholar] [CrossRef]
- He, Y.; Gao, Y.; Hong, H.; Geng, J.; Chen, Q.; Zhou, Y.; Zhu, Z. Megalurothrips usitatus Directly Causes the Black-Heads and Black-Tail Symptoms of Cowpea along with the Production of Insect-Resistance Flavonoids. Plants 2023, 12, 3865. [Google Scholar] [CrossRef]
- Tang, L.D.; Yan, K.L.; Fu, B.L.; Wu, J.H.; Liu, K.; Lu, Y.Y. The life table parameters of Megalurothrips usitatus (Thysanoptera: Thripidae) on four leguminous crops. Fla. Entomol. 2015, 98, 620–625. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.L.; Kirk, W.D.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B. Thrips. In Polyphagous Pests Crops; Springer: New York, NY, USA, 2021. [Google Scholar]
- Gao, Y.F.; Gong, Y.J.; Cao, L.J.; Chen, J.C.; Gao, Y.L.; Mirab-Balou, M.; Chen, M.; Hoffmann, A.A.; Wei, S.J. Geographical and interspecific variation in susceptibility of three common thrips species to the insecticide, spinetoram. J. Pest Sci. 2021, 94, 93–99. [Google Scholar] [CrossRef]
- Fu, B.; Tao, M.; Xue, H.; Jin, H.; Liu, K.; Qiu, H.; Yang, S.; Yang, X.; Gui, L.; Zhang, Y.; et al. Spinetoram resistance drives interspecific competition between Megalurothrips usitatus and Frankliniella intonsa. Pest Manag. Sci. 2022, 78, 2129–2140. [Google Scholar] [CrossRef]
- Gao, R.; Lu, R.; Qiu, X.; Wang, L.; Zhang, K.; Wu, S. Detection of Putative Mutation I873S in the Sodium Channel of Megalurothrips usitatus (Bagnall) Which May Be Associated with Pyrethroid Resistance. Insects 2023, 14, 388. [Google Scholar] [CrossRef]
- Chen, J.C.; Cao, L.J.; Sun, L.N.; Gao, Y.F.; Cao, H.Q.; Ma, Z.Z.; Ma, L.J.; Shen, X.J.; Wang, J.X.; Gong, Y.J. Variation in the toxicity of a novel meta-diamide insecticide, broflanilide, among thrips pest species and developmental stages. Pest Manag. Sci. 2022, 78, 5090–5096. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, L.; Chen, J.; Huang, H.; Liu, K.; Zhang, Y.; Yang, J.; Wu, S. Mutation V65I in the β1 Subunit of the Nicotinic Acetylcholine Receptor Confers Neonicotinoid and Sulfoxaflor Resistance in Insects. J. Agric. Food Chem. 2024, 72, 5671–5681. [Google Scholar] [CrossRef]
- Tang, L.D.; Fu, B.L.; Qiu, H.Y.; Han, Y.; Li, P.; Liu, K. Studied on the toxicity of different insecticides to against Megalurothrips usitatus by using a modified TIBS method. Chin. J. Trop. Crops 2015, 36, 570–574. [Google Scholar]
- Tang, L.D.; Zhao, H.; Fu, B.L.; Qiu, H.Y.; Wu, J.; Li, P.; Liu, K. Insecticide resistance monitoring of the Hainan field populations of Megalurothrips usitatus and their susceptibility to 6 insecticides. J. Environ. Entomol. 2018, 40, 1175–1181. [Google Scholar]
- Zhang, W.; Jiang, F.; Ou, J. Global pesticide consumption and pollution: With China as a focus. Proc. Int. Acad. Ecol. Environ. Sci. 2011, 1, 125. [Google Scholar]
- Gao, Y.; Reitz, S.R.; Wei, Q.; Yu, W.; Zhang, Z.; Lei, Z. Local Crop Planting Systems Enhance Insecticide-Mediated Displacement of Two Invasive Leafminer Fly. PLoS ONE 2014, 9, e92625. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.C.; Chen, H.; Babendreier, D.; Zhang, J.P.; Zhang, F.; Dai, X.Y.; Sun, Z.W.; Shi, Z.P.; Dong, X.L.; Wu, G.A. Improved control of Frankliniella occidentalis on greenhouse pepper through the integration of Orius sauteri and neonicotinoid insecticides. J. Pest Sci. 2021, 94, 101–109. [Google Scholar] [CrossRef]
- Sun, J. Study on Monitoring and Mechanism of Spinosad Resistance in Megalurothrips usitatus (Bagnall). Master’s Thesis, Hainan University, Haikou, China, 2017. [Google Scholar]
- Qiu, H.Y.; Fu, B.L.; Tan, K.; He, S.; Chen, Z.; Luo, J.; Liu, K. Resistance monitoring to multiple insecticides in a field population of Hainan Megalurothrips usitatus (Bagnall). Chin. Plant Prot. 2022, 42, 67–71. [Google Scholar]
- Van, L.T.; Dermauw, W.; Mavridis, K.; Vontas, J. Significance and interpretation of molecular diagnostics for insecticide resistance management of agricultural pests. Curr. Opin. Insect Sci. 2020, 39, 69–76. [Google Scholar]
- Kirst, H.A. The spinosyn family of insecticides: Realizing the potential of natural products research. J. Antibiot. 2010, 63, 101–111. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crouse, G.D.; Benko, Z.; Demeter, D.; Giampietro, N.C.; Lambert, W.; Brown, A.V. The spinosyns, spinosad, spinetoram, and synthetic spinosyn mimics-discovery, exploration, and evolution of a natural product chemistry and the impact of computational tools. Pest Manag. Sci. 2021, 77, 3637–3649. [Google Scholar] [CrossRef]
- Bielza, P.; Quinto, V.; Contreras, J.; Torne, M.; Martin, A.; Espinosa, P.J. Resistance to spinosad in the western flower thrips, Frankliniella occidentalis (Pergande), in greenhouses of south-eastern Spain. Pest Manag. Sci. 2007, 63, 682–687. [Google Scholar] [CrossRef]
- Funderburk, J. Management of the western flower thrips (Thysanoptera: Thripidae) in fruiting vegetables. Fla. Entomol. 2009, 92, 1–6. [Google Scholar] [CrossRef]
- Wu, S.; Tang, L.D.; Zhang, X.; Xing, Z.; Lei, Z.; Gao, Y. A decade of a thrips invasion in China: Lessons learned. Ecotoxicology 2018, 27, 1032–1038. [Google Scholar] [CrossRef]
- Schnaars-Uvino, K.; Baker, M.B. High-level field-evolved resistance to spinosad in Colorado potato beetle, Leptinotarsa decemlineata, in organically managed fields. Pest Manag. Sci. 2021, 77, 4393–4399. [Google Scholar] [CrossRef]
- Campos, M.R.; Silva, T.B.M.; Silva, W.M.; Silva, J.E.; Siqueira, H.A.A. Spinosyn resistance in the tomato borer Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). J. Pest Sci. 2015, 88, 405–412. [Google Scholar] [CrossRef]
- Yuan, J.; Zheng, X.; Wang, J.; Qian, K.; Feng, J.; Zhang, Y.; Zhang, K.; Zhang, Y.; Liang, P.; Wu, Q. Insecticide resistance of western flower thrips, Frankliniella occidentalis (Pergande) in China. Crop Prot. 2023, 172, 106339. [Google Scholar] [CrossRef]
- Bielza, P.; Guillén, J. Cyantraniliprole: A valuable tool for Frankliniella occidentalis (Pergande) management. Pest Manag. Sci. 2015, 71, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Duan, A.; Li, H.; Yuan, X.; Zhao, M. Toxicity and Effects of Cyantraniliprole against Thrips of Cotton in Fields. Xinjiang Agric. Sci. 2020, 57, 665. [Google Scholar]
- Nakao, T.; Banba, S. Broflanilide: A meta-diamide insecticide with a novel mode of action. Bioorganic Med. Chem. 2016, 24, 372–377. [Google Scholar] [CrossRef]
- Houchat, J.N.; Cartereau, A.; Le, M.A.; Taillebois, E.; Thany, S.H. An overview on the effect of neonicotinoid insecticides on mammalian cholinergic functions through the activation of neuronal nicotinic acetylcholine receptors. Int. J. Environ. Res. Public Health 2020, 17, 3222. [Google Scholar] [CrossRef]
- Yu, X.R.; Tariq, T.; Guo, L.H.; Wu, S.Y.; Tang, L.D.; Zang, L.S. Assessing the effectiveness of imidacloprid and thiamethoxam via root irrigation against Megalurothrips usitatus (Thysanoptera: Thripidae) and its residual effects on cowpea. J. Econ. Entomol. 2023, 116, 1767–1775. [Google Scholar] [CrossRef]
- Peng, Z.; Zheng, H.; Xie, W.; Wang, S.; Wu, Q.; Zhang, Y. Field resistance monitoring of the immature stages of the whitefly Bemisia tabaci to spirotetramat in China. Crop Prot. 2017, 98, 243–247. [Google Scholar] [CrossRef]
- Zhang, K.; Yuan, J.; Wang, J.; Hua, D.; Zheng, X.; Tao, M.; Zhang, Z.; Wan, Y.; Wang, S.; Zhang, Y.; et al. Susceptibility levels of field populations of Frankliniella occidentalis (Thysanoptera: Thripidae) to seven insecticides in China. Crop Prot. 2022, 153, 105886. [Google Scholar] [CrossRef]
- Fu, B.L.; Qiu, H.Y.; Li, Q.; Tang, L.D.; Zeng, D.; Liu, K.; Gao, Y. Flower injection of imidacloprid and spirotetramat: A novel tool for the management of banana thrips Thrips hawaiiensis. J. Pest Sci. 2020, 93, 1073–1084. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, Q.; Yang, L.; Zhu, S.; Zhang, Z.; Xu, H. Efficacy of drip irrigation with thiamethoxam on control of Monolepta hieroglyphica, and uptake, translocation and dietary risk of thiamethoxam in maize. Pest Manag. Sci. 2023, 79, 4931–4941. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.D.; Guo, L.H.; Ali, A.; Desneux, N.; Zang, L.S. Synergism of Adjuvants Mixed with Spinetoram for the Management of Bean Flower Thrips, Megalurothrips usitatus (Thysanoptera: Thripidae) in Cowpeas. J. Econ. Entomol. 2022, 115, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Bilbo, T.R.; Kennedy, G.G.; Walgenbach, J.F. Western flower thrips (Frankliniella occidentalis) field resistance to spinetoram in North Carolina. Crop Prot. 2023, 165, 106168. [Google Scholar] [CrossRef]
- Bojan, V. Bioefficacy, phytotoxicity, safety to natural enemies and residues of cyantraniliprole 10 OD on potato (Solanum tuberosum L.) under open field condition. Crop Prot. 2021, 142, 105505. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Field Population | Insecticide | Number of Tested Thrips | LC50 (mg a.i./L) (95% Confidence Interval) | Slope (SE) | C2 | Df | p |
|---|---|---|---|---|---|---|---|
| QY | broflanilide | 335 | 7.40 (4.45–11.93) | 0.71 ± 0.09 | 1.52 | 4 | 0.82 |
| dinotefuran | 329 | 56.884 (35.732–90.489) | 0.70 ± 0.08 | 0.81 | 4 | 0.94 | |
| spinetoram | 286 | 0.14 (0.03–0.41) | 1.00 ± 0.13 | 7.13 | 5 | 0.07 | |
| spinosad | 264 | 0.27 (0.10–0.73) | 0.91 ± 0.11 | 4.89 | 3 | 0.18 | |
| cyantraniliprole | 321 | 60.77 (15.81–187.39) | 0.76 ± 0.11 | 7.02 | 4 | 0.13 | |
| spirotetramat | 334 | 987.80 (363.00–1686.11) | 2.67 ± 0.42 | 8.83 | 4 | 0.07 | |
| emamectin benzoate | 386 | 17.87 (6.29–43.80) | 0.87 ± 0.09 | 10.58 | 5 | 0.06 | |
| avermectin | 276 | 43.31 (18.96–80.42) | 0.71 ± 0.10 | 2.45 | 3 | 0.48 | |
| thiamethoxam | 383 | 174.91 (60.15–292.15) | 1.83 ± 0.31 | 7.55 | 5 | 0.18 | |
| chlorfenapyr | 283 | 36.35 (12.06–102.01) | 0.70 ± 0.10 | 10.45 | 5 | 0.06 |
| Field Population | Insecticide | Number of Tested Thrips | LC50 (mg a.i./L) (95% Confidence Interval) | Slope (SE) | C2 | Df | p |
|---|---|---|---|---|---|---|---|
| YF | broflanilide | 253 | 13.93 (6.40–25.26) | 0.73 ± 0.12 | 2.11 | 4 | 0.72 |
| dinotefuran | 328 | 112.80 (29.16–326.25) | 0.77 ± 0.16 | 5.56 | 5 | 0.35 | |
| spinetoram | 420 | 0.67 (0.20–1.12) | 3.68 ± 0.70 | 10.20 | 5 | 0.07 | |
| spinosad | 424 | 2.03 (1.16–3.03) | 4.20 ± 0.78 | 6.911 | 5 | 0.23 | |
| cyantraniliprole | 415 | 35.67 (21.54–51.06) | 1.97 ± 0.35 | 1.84 | 5 | 0.87 | |
| spirotetramat | 362 | 1237.15 (605.54–1752.98) | 2.86 ± 0.59 | 5.47 | 5 | 0.36 | |
| emamectin benzoate | 361 | 0.59 (0.14–1.55) | 0.88 ± 0.10 | 8.74 | 4 | 0.07 | |
| avermectin | 385 | 675.73 (341.04–1843.43) | 0.70 ± 0.12 | 3.63 | 5 | 0.60 | |
| thiamethoxam | 382 | 103.97 (29.04–202.76) | 1.43 ± 0.21 | 8.94 | 5 | 0.11 | |
| chlorfenapyr | 360 | 122.28 (59.81–222.18) | 0.99 ± 0.18 | 2.70 | 5 | 0.75 |
| Field Population | Insecticide | Number of Tested Thrips | LC50 (mg a.i./L) (95% Confidence Interval) | Slope (SE) | C2 | Df | p |
|---|---|---|---|---|---|---|---|
| MM | broflanilide | 264 | 71.33 (38.40~126.72) | 0.72 ± 0.12 | 1.63 | 5 | 0.90 |
| dinotefuran | 232 | 13.24 (2.41~39.41) | 0.70 ± 0.10 | 6.01 | 4 | 0.20 | |
| spinetoram | 327 | 0.26 (0.17~0.39) | 1.47 ± 0.19 | 2.31 | 5 | 0.81 | |
| spinosad | 343 | 0.40 (0.24~0.59) | 1.35 ± 0.18 | 2.63 | 5 | 0.76 | |
| cyantraniliprole | 216 | 11.02 (4.47~20.17) | 0.75 ± 0.14 | 0.73 | 3 | 0.40 | |
| spirotetramat | 286 | 113.19 (62.53~178.25) | 1.30 ± 0.18 | 2.94 | 5 | 0.71 | |
| emamectin benzoate | 199 | 4.82 (0.65~14.17) | 0.74 ± 0.13 | 3.49 | 3 | 0.32 | |
| avermectin | 208 | 360.80 (68.94~989.34) | 0.78 ± 0.16 | 3.19 | 3 | 0.36 | |
| thiamethoxam | 285 | 15.74 (2.81~40.69) | 1.00 ± 0.14 | 6.34 | 4 | 0.17 | |
| chlorfenapyr | 243 | 19.08 (8.41~34.15) | 1.19 ± 0.15 | 5.32 | 5 | 0.38 |
| Insecticide | LC50 (95% Confidence Interval) | Resistance Ratio (RR) |
|---|---|---|
| spinetoram | 0.017 (0.010–0.028) Susceptible strain a | 8.47 (QY) |
| 39.47 (YF) | ||
| 15.53 (MM) | ||
| emamectin benzoate | 0.006 (0.004–0.011) Susceptible strain a | 2978 (QY) |
| 98.50 (YF) | ||
| 802.67 (MM) | ||
| avermectin | 0.276 (0.129–0.594) Susceptible strain a | 156.92 (QY) |
| 2448.30 (YF) | ||
| 1307.25 (MM) | ||
| spirotetramat | 3.143 (1.748–5.650) Susceptible strain a | 314.29 (QY) |
| 939.62 (YF) | ||
| 36.01 (MM) | ||
| spinosad | 0.172 (0.091–0.249) Susceptible strain b | 1.59 (QY) |
| 11.80 (YF) | ||
| 2.30 (MM) | ||
| chlorfenapyr | 0.268 (0.155–0.462) Susceptible strain a | 135.63 (QY) |
| 456.27 (YF) | ||
| 79.19 (MM) |
| Insecticides | Dosage (g a.i./ha) | Initial Number (10 Plants) | Number of Thrips (10 Plants) | Reduction Rate (%) | Corrected Control Efficacy (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 dpa | 3 dpa | 5 dpa | 7 dpa | 1 dpa | 3 dpa | 5 dpa | 7 dpa | 1 dpa | 3 dpa | 5 dpa | 7 dpa | |||
| cyantraniliprole | 60 | 100 | 56 | 50 | 42 | 28 | 44.00 | 50.25 | 57.75 | 71.75 | 52.58 a | 59.98 a | 61.08 a | 76.35 a |
| spirotetramat | 100.8 | 86 | 71 | 45 | 41 | 41 | 17.39 | 48.12 | 52.46 | 52.46 | 30.05 a | 58.27 a | 56.21 a | 60.21 ab |
| emamectin benzoate | 3.375 | 104 | 64 | 62 | 44 | 37.5 | 38.65 | 40.58 | 57.25 | 63.77 | 48.05 a | 52.20 a | 60.62 a | 69.67 ab |
| thiamethoxam | 75 | 105 | 74 | 78 | 50 | 52 | 29.12 | 26.01 | 52.51 | 50.36 | 39.98 a | 40.49 a | 56.25 a | 58.44 b |
| spinetoram | 18 | 98 | 66 | 59 | 49 | 29 | 32.91 | 39.54 | 50.26 | 70.41 | 43.19 a | 51.37 a | 54.18 a | 75.23 a |
| Water control | - | 129 | 152 | 160 | 140 | 154 | −18.09 | −24.32 | −8.56 | −19.46 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Z.; Li, M.; Guo, C.; Zheng, H.; Wu, M.; Yin, F.; Xiao, Y.; Wang, H.; Kong, X.; Zalucki, M.P.; et al. Field-Based Evaluation of Insecticide Effectiveness on Megalurothrips usitatus in Guangdong, China: Implications for Pest Control Strategies. Insects 2025, 16, 459. https://doi.org/10.3390/insects16050459
Peng Z, Li M, Guo C, Zheng H, Wu M, Yin F, Xiao Y, Wang H, Kong X, Zalucki MP, et al. Field-Based Evaluation of Insecticide Effectiveness on Megalurothrips usitatus in Guangdong, China: Implications for Pest Control Strategies. Insects. 2025; 16(5):459. https://doi.org/10.3390/insects16050459
Chicago/Turabian StylePeng, Zhengke, Mengni Li, Chaosong Guo, Huixin Zheng, Mingyue Wu, Fei Yin, Yong Xiao, Huanhuan Wang, Xiangyi Kong, Myron P. Zalucki, and et al. 2025. "Field-Based Evaluation of Insecticide Effectiveness on Megalurothrips usitatus in Guangdong, China: Implications for Pest Control Strategies" Insects 16, no. 5: 459. https://doi.org/10.3390/insects16050459
APA StylePeng, Z., Li, M., Guo, C., Zheng, H., Wu, M., Yin, F., Xiao, Y., Wang, H., Kong, X., Zalucki, M. P., Xie, W., & Li, Z. (2025). Field-Based Evaluation of Insecticide Effectiveness on Megalurothrips usitatus in Guangdong, China: Implications for Pest Control Strategies. Insects, 16(5), 459. https://doi.org/10.3390/insects16050459