
Effects of Sex, Age, and Body Size on Flight Performance of Monochamus alternatus (Coleoptera: Cerambycidae), a Vector of Pine Wood Nematodes, Using Flight Mills


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Insects
2.2. Measurement of Flight Distance, Time, and Speed
2.3. Data Analyses
3. Results
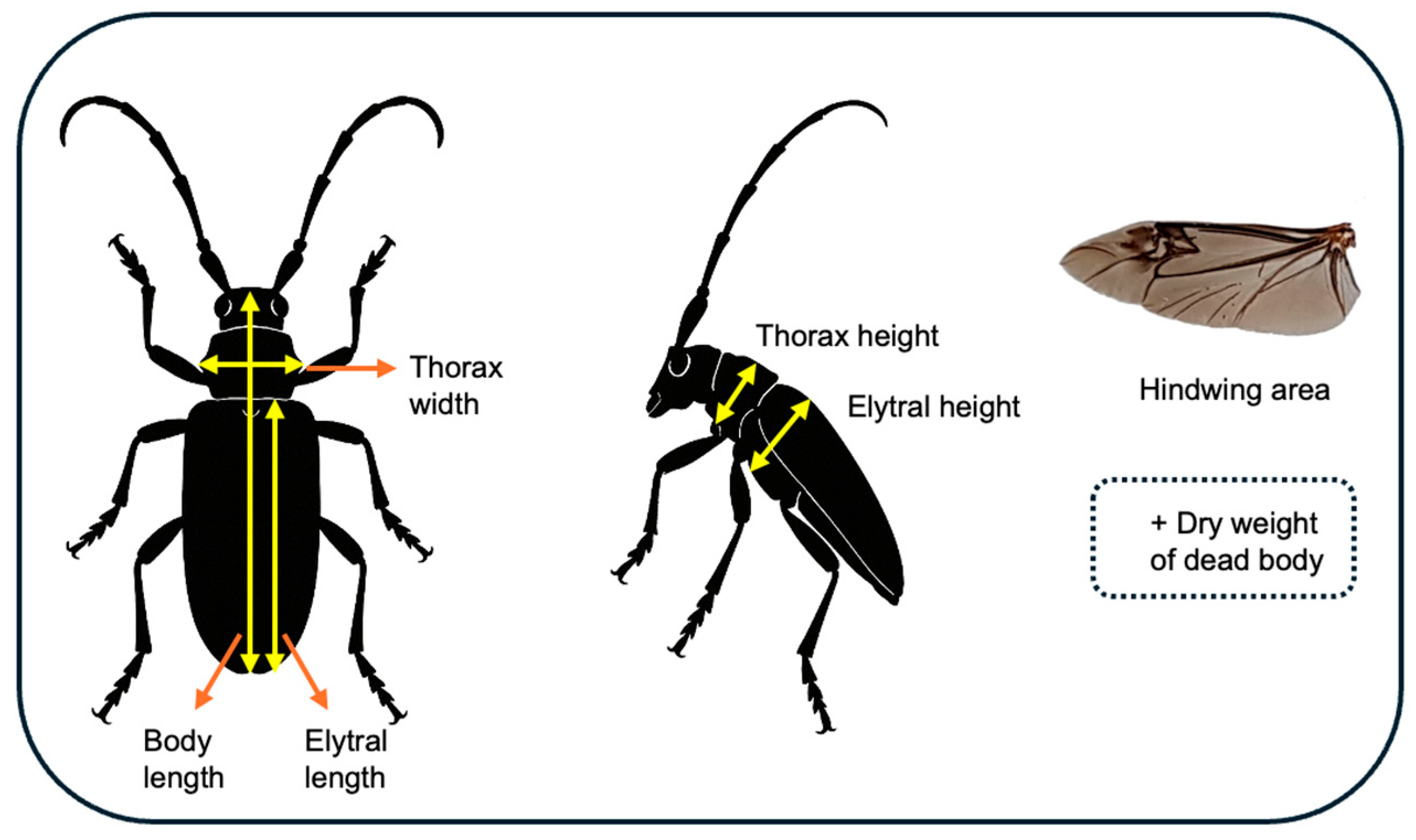
3.1. Summary of Morphological Traits and Flight Experiments
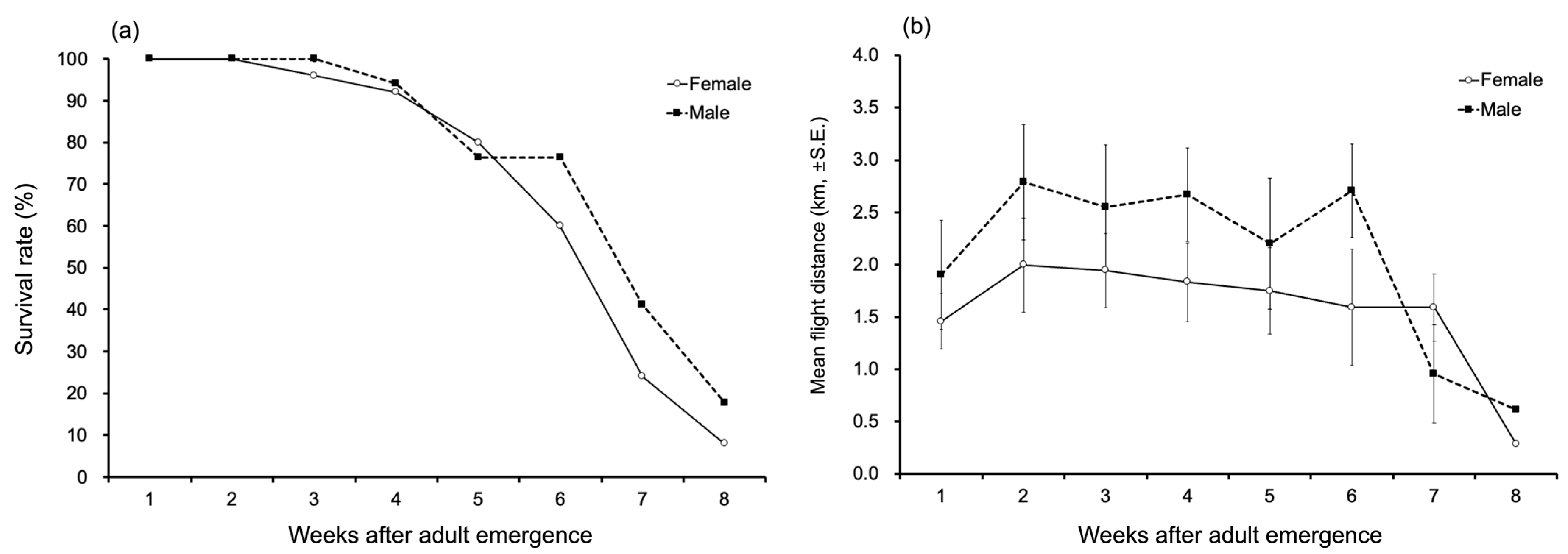
3.2. Change in Survival Rates Across Lifespan
3.3. Total Flight Distance, Time, and Speed
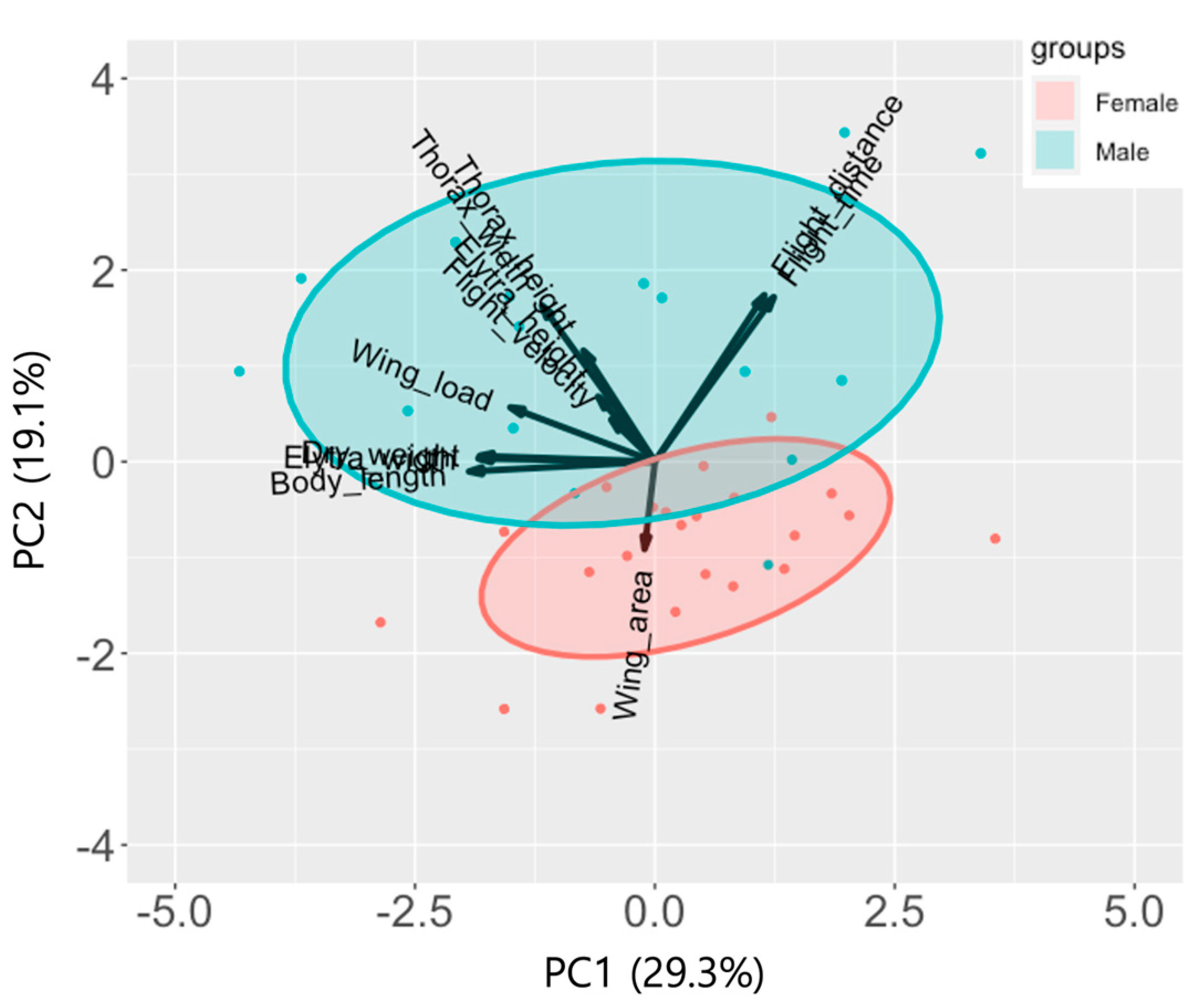
3.4. Principal Component Analysis
4. Discussion
4.1. Flight Capability of M. alternatus Tethered on Flight Mills
4.2. Sex-Dependent Flight Performance of M. alternatus
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| PWN | Pine wood nematode |
| LMM | Linear mixed models |
| PCA | Principal component analysis |
Appendix A

References
- Shin, S.C. Pine Wilt Disease in Korea. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar]
- Akbulut, S.; Stamps, W.T. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Akbulut, S.; Togashi, K.; Linit, M.J. Cerambycids as plant disease vectors with special reference to pine wilt. In Cerambycidae of the World: Biology and Pest Management; Wang, Q., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 209–252. [Google Scholar]
- Rodrigues, J.M.; Sousa, E.; Abrantes, I. Pine wilt disease historical overview. In Pine Wilt Disease in Europe—Biological Interactions and Integrated Management; Sousa, E., Vale, F., Abrantes, I., Eds.; Federação Nacional Associações de Proprietários Florestais: Lisbon, Spain, 2015; pp. 13–32. [Google Scholar]
- Choi, Y.E.; Moon, Y.S. Survey on distribution of pine wood nematode (Bursaphelenchus xylophilus) and its pathogenicity to pine trees in Korea. Kor. J. Plant Pathol. 1989, 5, 277–286. [Google Scholar]
- KFS. A Plan for the Forecasting and Control of Forest Insect Pests and Diseases in 2025, Korea Forest Service. Available online: https://www.forest.go.kr (accessed on 28 February 2025).
- Jung, J.M.; Yoon, S.; Hwang, J.; Park, Y.; Lee, W.H. Analysis of the spread distance of pine wilt disease based on a high volume of spatiotemporal data recording of infected trees. For. Ecol. Manage. 2024, 553, 121612. [Google Scholar] [CrossRef]
- EPPO. Monochamus alternatus. EPPO Datasheets on Pests Recommended for Regulation. Available online: https://gd.eppo.int (accessed on 4 July 2024).
- Kobayashi, F.; Yamane, A.; Ikeda, T. The Japanese pine sawyer beetle as the vector of pine wilt disease. Ann. Rev. Entomol. 1984, 29, 115–135. [Google Scholar] [CrossRef]
- Wang, Z.M.; Pi, Z.Q.; Hou, B. Monochamus alternatus found in Jilin Province. For. Pest Dis. 2006, 25, 35. (In Chinese) [Google Scholar]
- Kwon, T.S.; Lim, J.H.; Sim, S.J.; Kwon, Y.D.; Son, S.K.; Lee, K.Y.; Kim, Y.T.; Park, J.W.; Shin, C.H.; Ryu, S.B.; et al. Distribution patterns of Monochamus alternatus and M. saltuarius (Coleoptera: Cerambycidae) in Korea. J. Kor. For. Soc. 2006, 95, 543–550. [Google Scholar]
- Fauziah, B.A.; Hidaka, T.; Tabata, K. The reproductive behavior of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1987, 22, 272–285. [Google Scholar] [CrossRef]
- Mamiya, M. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar]
- Togashi, K. Change in the activity of adult Monochamus alternatus Hope (Coleoptera: Cerambycidae) in relation to age. Appl. Entomol. Zool. 1990, 25, 153–159. [Google Scholar] [CrossRef]
- Fan, J.T.; Sun, J.H.; Shi, J. Attraction of the Japanese pine sawyer, Monochamus alternatus, to volatiles from stressed host in China. Ann. For. Sci. 2007, 64, 67–71. [Google Scholar] [CrossRef]
- Li, S.Q.; Fang, Y.L.; Zhang, Z.N. Effects of volatiles of non-host plants and other chemicals on oviposition of Monochamus alternatus (Coleoptera: Cerambycidae). J. Pestic. Sci. 2007, 80, 119–123. [Google Scholar] [CrossRef]
- NIFOS. Illustration Book of Forest Insect Pest, 2nd ed.; National Institute of Forest Science; Utgomunhwasa Publisher: Seoul, Republic of Korea, 2008; pp. 223–225. [Google Scholar]
- Sugimoto, H.; Fukuta, M.; Togashi, K. Yearly changes in dispersal and life-history traits of Monochamus alternatus Hope with reference to its outbreak. J. Appl. Entomol. 2020, 144, 459–467. [Google Scholar] [CrossRef]
- Sanchez-Husillos, E.; Etxebeste, I.; Pajares, J. Physiological development and dispersal ability of newly emerged Monochamus galloprovincialis. Entomol. Exp. Appl. 2016, 161, 141–151. [Google Scholar] [CrossRef]
- Robinet, C.; David, G.; Jactel, H. Modeling the distances traveled by flying insects based on the combination of flight mill and mark-release-recapture experiments. Ecol. Modell. 2019, 402, 85–92. [Google Scholar] [CrossRef]
- Lee, H.R.; Lee, S.C.; Lee, D.H.; Jung, M.; Kwon, J.H.; Huh, M.J.; Kim, D.S.; Lee, J.E.; Park, I.K. Identification of aggregation-sex pheromone of the Korean Monochamus alternatus (Coleoptera: Cerambycidae) population, the main vector of pine wood nematode. J. Econ. Entomol. 2018, 111, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jung, Y.H.; Lee, S.M. Diel rhythmicity of field responses to synthetic pheromone lures in the pine sawyer Monochamus saltuarius. Insects 2021, 12, 441. [Google Scholar] [CrossRef]
- David, G.; Giffard, B.; Piou, D.; Jactel, H. Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. J. Appl. Entomol. 2014, 138, 566–576. [Google Scholar] [CrossRef]
- Etxebeste, I.; Sanchez-Husillos, E.; Álvarez, G.; Gisbert, H.M.; Pajares, J. Dispersal of Monochamus galloprovincialis (Col.: Cerambycidae) as recorded by mark–release–recapture using pheromone traps. J. Appl. Entomol. 2016, 140, 485–499. [Google Scholar] [CrossRef]
- Humphry, S.J.; Linit, M.J. Tethered flight of Monochamus carolinensis (Coleoptera: Cerambycidae) with respect to beetle age and sex. Environ. Entomol. 1989, 18, 124–126. [Google Scholar] [CrossRef]
- Humphry, S.J.; Linit, M.J. Effect of pinewood nematode density on tethered flight of Monochamus carolinensis (Coleoptera: Cerambycidae). Environ. Entomol. 1989, 18, 670–673. [Google Scholar] [CrossRef]
- Akbulut, S.; Linit, M.J. Flight performance of Monochamus carolinensis (Coleoptera: Cerambycidae) with respect to nematode phoresis and beetle characteristics. Environ. Entomol. 1999, 28, 1014–1020. [Google Scholar] [CrossRef]
- Kwon, H.J.; Jung, J.K.; Jung, C.S.; Han, H.R.; Koh, S.H. Dispersal capacity of Monochamus saltuarius on flight mills. Entomol. Exp. Appl. 2018, 166, 420–427. [Google Scholar] [CrossRef]
- Ito, K. The tethered flight of the Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae). J. Jap. For. Soc. 1982, 64, 395–397. [Google Scholar]
- Enda, N. The flight ability of the Japanese pine-sawyer (Coleoptera, Cerambycidae) as determined by a flight-mill instrument. In Transactions of the 96th Meeting of Japanese Forest Society; The Japanese Forest Society: Tokyo, Japan, 1985; pp. 517–518. [Google Scholar]
- Choi, W.I.; Song, H.J.; Kim, D.S.; Lee, D.S.; Lee, C.Y.; Nam, Y.; Kim, J.B.; Park, Y.S. Dispersal patterns of pine wilt disease in the early stage of its invasion in South Korea. Forests 2017, 8, 411. [Google Scholar] [CrossRef]
- Jung, J.K.; Kim, M.; Nam, Y.; Koh, S.H. Changes in spatial and temporal distributions of Monochamus beetles along the fire severity in burned Pinus densiflora forests. J. Asia. Pac. Entomol. 2020, 23, 404–410. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 21 December 2024).
- Lee, S.M.; Chung, Y.J.; Kim, D.S.; Choi, K.S.; Kim, Y.G.; Park, C.G. Adult morphological measurements: An indicator to identify sexes of Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae). Kor. J. Appl. Entomol. 2004, 43, 85–89. [Google Scholar]
- Yu, K.; Reddy, G.V.P.; Schrader, J.; Guo, X.; Li, Y.; Jiao, Y.; Shi, P. A nondestructive method of calculating the wing area of insects. Ecol. Evol. 2022, 12, e8792. [Google Scholar] [CrossRef]
- Park, W.H.; Ko, D.W.; Kwon, T.S.; Nam, Y.; Kwon, Y.D. Analysis of pinewood nematode damage expansion in Gyeonggi province based on monitoring data from 2008 to 2015. J. Kor. Soc. For. Sci. 2018, 107, 486–496. [Google Scholar]
- El-Sayed, A.M.; Suckling, D.M.; Wearing, C.H.; Byers, J.A. Potential of mass trapping for long-term pest management and eradication of invasive species. J. Econ. Entomol. 2006, 99, 1550–1564. [Google Scholar] [CrossRef]
- Schroeder, M. Trapping strategy for Monochamus sutor and Monochamus galloprovincialis: Potential vectors of the pine wood nematode in Scandinavia. Agri. For. Entomol. 2019, 21, 372–378. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Traits | Male (n = 17) | Female (n = 25) | t | d.f. | p |
|---|---|---|---|---|---|
| Dry weight (mg) | 153.59 ± 38.65 | 151.48 ± 41.34 | 0.17 | 40 | 0.8686 |
| Body length (mm) | 24.85 ± 3.76 | 25.29 ± 2.38 | −0.46 | 40 | 0.6480 |
| Thorax width (mm) | 6.49 ± 0.52 | 5.17 ± 0.61 | 3.59 | 40 | 0.0009 |
| Thorax height (mm) | 5.99 ± 0.58 | 5.18 ± 0.7 | 3.95 | 40 | 0.0003 |
| Elytral length (mm) | 21.67 ± 2.14 | 23.31 ± 2.37 | −32.35 | 40 | 0.0000 |
| Elytral width (mm) | 7.37 ± 0.76 | 8.01 ± 0.88 | −2.43 | 40 | 0.0196 |
| Hindwing area (mm2) | 147.64 ± 30.36 | 173.74 ± 31.29 | −2.69 | 40 | 0.0105 |
| Wing load (mg/mm2) | 1.12 ± 0.56 | 0.86 ± 0.16 | 2.16 | 40 | 0.0368 |
| Sex | n | Flight Distance (km) | Flight Time (min.) | Flight Speed for Fliers (m s−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| a Total | b Non- Fliers | c Fliers | Mean ± S.E. | Range (Min.~Max.) | Mean ± S.E. | Range (Min.~Max.) | Mean ± S.E. | Range (Min.~Max.) | |
| Total flight | |||||||||
| Female | 25 | 3 | 22 | 6.65 ± 0.75 | 0.13~15.35 | 59.56 ± 31.38 | 1.62~131.50 | 1.86 ± 0.23 | 1.32~2.34 |
| Male | 17 | 1 | 16 | 9.89 ± 1.98 | 0.17~29.01 | 85.83 ± 69.37 | 1.39~241.07 | 1.93 ± 0.22 | 1.34~2.32 |
| Flight per session | |||||||||
| Female | 140 | 60 | 84 | 1.76 ± 0.15 | 0.10~7.09 | 13.84 ± 4.50 | 3.12~17.13 | 1.81 ± 0.16 | 1.52~2.05 |
| Male | 103 | 36 | 67 | 2.36 ± 0.21 | 0.12~7.57 | 17.75 ± 6.93 | 7.51~25.09 | 1.86 ± 0.22 | 1.36~2.04 |
| Dependent Variables | Fixed Effect | Num d.f. | Den d.f. | F | p |
|---|---|---|---|---|---|
| Flight distance | Sex | 1 | 143.95 | 4.12 | 0.044 |
| Session | 7 | 136.67 | 1.48 | 0.181 | |
| Flight time | Sex | 1 | 145.56 | 3.17 | 0.077 |
| Session | 7 | 138.02 | 1.19 | 0.311 | |
| Flight speed | Sex | 1 | 141.21 | 1.51 | 0.222 |
| Session | 7 | 134.98 | 1.72 | 0.109 |
| PC1 | PC2 | PC3 | PC4 | |
|---|---|---|---|---|
| Eigenvalue | 1.794 | 1.449 | 1.306 | 1.137 |
| Proportion of variance | 0.293 | 0.191 | 0.155 | 0.118 |
| Cumulative proportion | 0.293 | 0.484 | 0.639 | 0.756 |
| Flight or morphological traits | ||||
| Flight speed | −0.19 | 0.19 | 0.28 | * −0.40 |
| Flight distance | ** 0.48 | *** 0.73 | * −0.33 | −0.30 |
| Flight time | ** 0.52 | *** 0.72 | −0.32 | −0.26 |
| Dry weight | *** −0.77 | 0.03 | * −0.35 | −0.31 |
| Body length | *** −0.82 | −0.04 | −0.22 | −0.28 |
| Thorax width | ** −0.50 | *** 0.69 | 0.10 | * 0.34 |
| Thorax height | −0.31 | ** 0.49 | 0.06 | *** 0.72 |
| Wing area | −0.05 | * −0.39 | *** −0.85 | 0.10 |
| Elytral width | *** −0.78 | 0.01 | −0.23 | 0.11 |
| Elytral height | −0.25 | 0.29 | ** −0.44 | 0.04 |
| Wing load | *** −0.64 | 0.24 | ** 0.52 | * −0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.-K.; Lee, C.; Jang, B.; Nam, Y. Effects of Sex, Age, and Body Size on Flight Performance of Monochamus alternatus (Coleoptera: Cerambycidae), a Vector of Pine Wood Nematodes, Using Flight Mills. Insects 2025, 16, 444. https://doi.org/10.3390/insects16050444
Jung J-K, Lee C, Jang B, Nam Y. Effects of Sex, Age, and Body Size on Flight Performance of Monochamus alternatus (Coleoptera: Cerambycidae), a Vector of Pine Wood Nematodes, Using Flight Mills. Insects. 2025; 16(5):444. https://doi.org/10.3390/insects16050444
Chicago/Turabian StyleJung, Jong-Kook, Chayoung Lee, Beomjun Jang, and Youngwoo Nam. 2025. "Effects of Sex, Age, and Body Size on Flight Performance of Monochamus alternatus (Coleoptera: Cerambycidae), a Vector of Pine Wood Nematodes, Using Flight Mills" Insects 16, no. 5: 444. https://doi.org/10.3390/insects16050444
APA StyleJung, J.-K., Lee, C., Jang, B., & Nam, Y. (2025). Effects of Sex, Age, and Body Size on Flight Performance of Monochamus alternatus (Coleoptera: Cerambycidae), a Vector of Pine Wood Nematodes, Using Flight Mills. Insects, 16(5), 444. https://doi.org/10.3390/insects16050444