

The Frequency and Spread of a GABA-Gated Chloride Channel Target-Site Mutation and Its Impact on the Efficacy of Ethiprole Against Neotropical Brown Stink Bug, Euschistus heros (Hemiptera: Pentatomidae)



Simple Summary
Abstract

1. Introduction
2. Materials and Methods
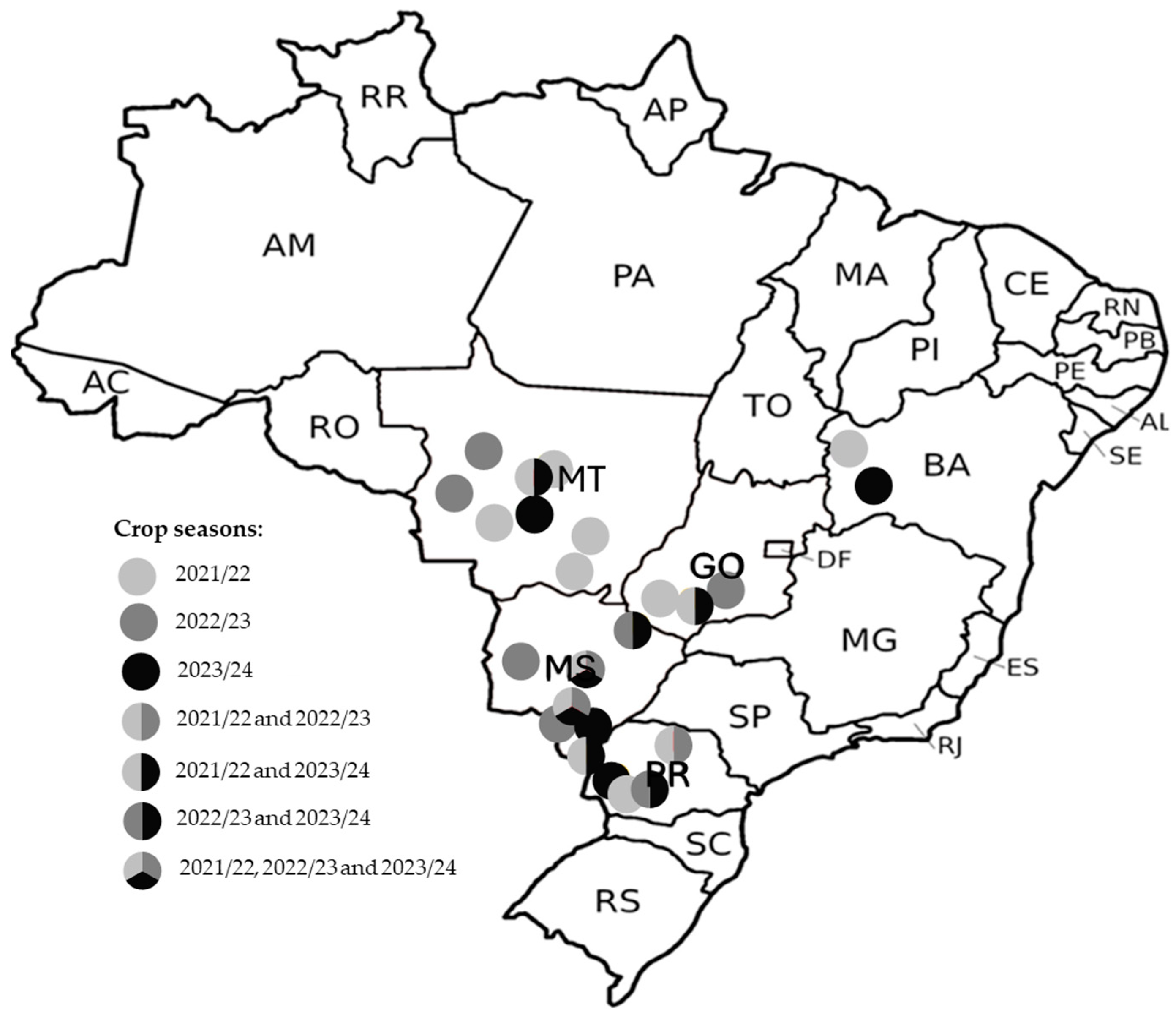
2.1. Brown Stink Bug Sampling and Rearing
2.2. Phenotypic Monitoring of E. heros Susceptibility to Ethiprole
2.3. DNA Extraction
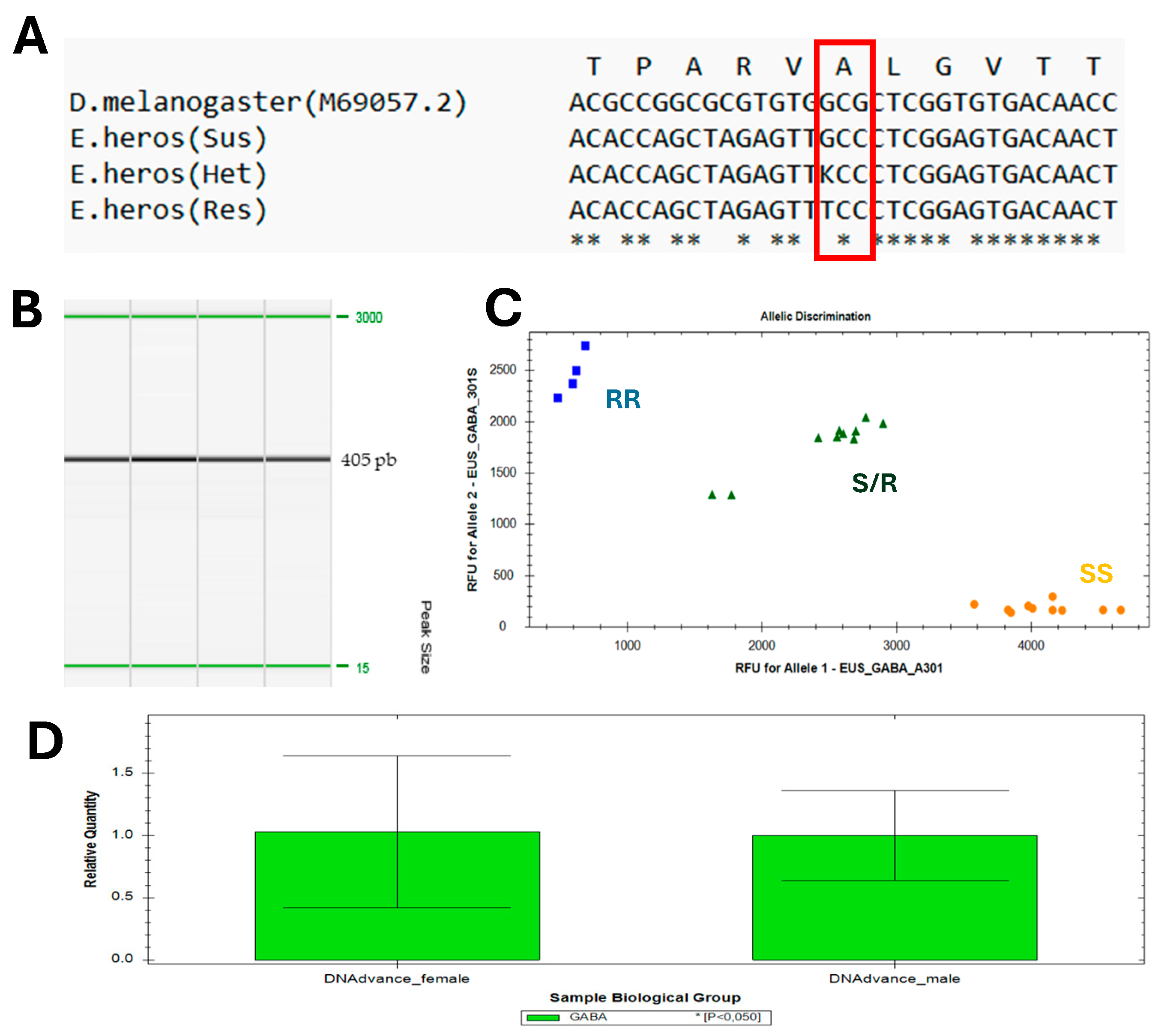
2.4. Partial Sequencing of the RDL-GABA-Gated Chloride Channel in E. heros
2.5. qPCR Assays to Monitor A301S Mutation in E. heros
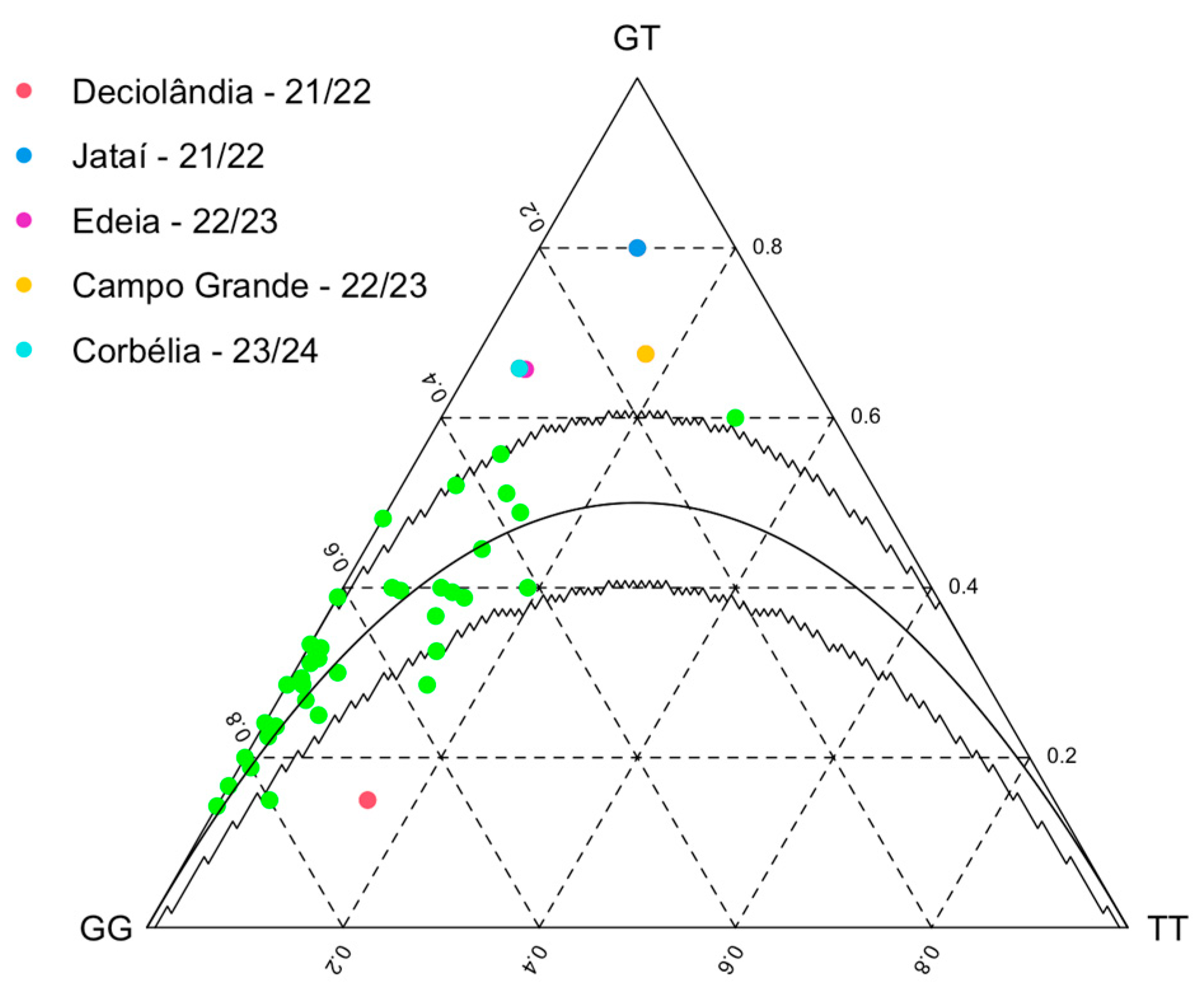
2.6. Statistical Analysis of Genetic Diversity and Population Differentiation
3. Results
3.1. Phenotypic Resistance Monitoring of E. heros Using the Label-Recommended Dose of Ethiprole
3.2. RDL-GABA-Gated Chloride Channel Partial Sequencing
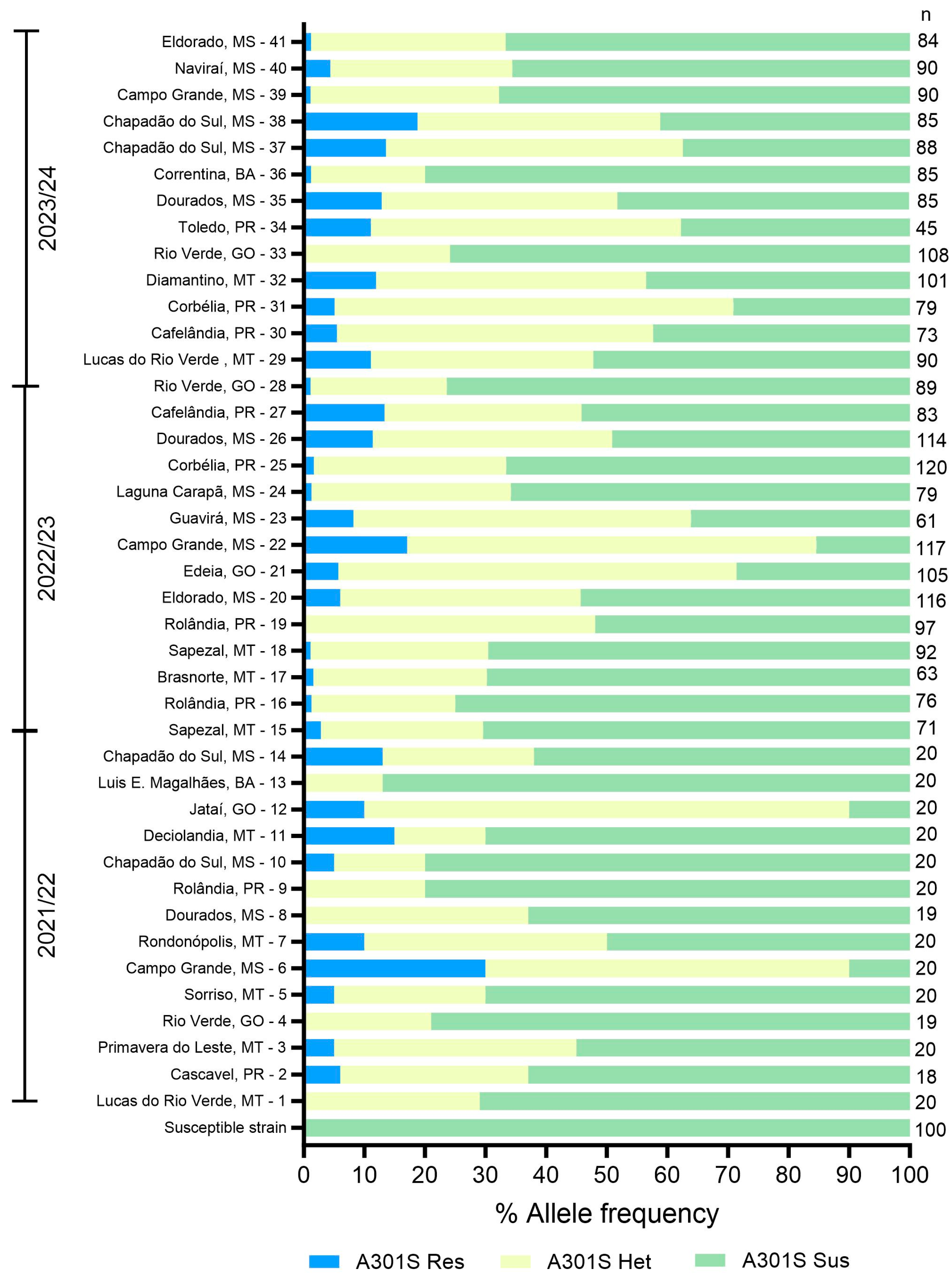
3.3. Genotyping of E. heros Samples
3.4. Relationship Between Ethiprole Efficacy and Resistance Allele Frequency in E. heros
3.5. Genotyping of E. heros Survivors of Vial Bioassays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, T.L.B.; Baldin, E.L.L.; Lima, A.P.S.; Santana, A.S.; Santos, M.C.; Silveira, B.R.R.; Bueno, N.M.; Cabral, I.R.; Soares, M.C.E.; Pinheiro, A.M.; et al. Intraspecific and Interspecific Interaction and Fitness Cost of Stink Bugs Euschistus heros, Diceraeus melacanthus, and Piezodorus guildinii in Soybean. Pest Manag. Sci. 2024, 80, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Soria, M.F.; Degrande, P.E.; Panizzi, A.R.; Toews, M.D. Economic Injury Level of the Neotropical Brown Stink Bug Euschistus heros (F.) on Cotton Plants. Neotrop. Entomol. 2017, 46, 324–335. [Google Scholar] [CrossRef]
- Ferreira Santos de Aquino, M.; Sujii, E.R.; Borges, M.; Blassioli Moraes, M.C.; Laumann, R.A. Diversity of Stink Bug Adults and Their Parasitoids in Soybean Crops in Brazil: Influence of a Latitudinal Gradient and Insecticide Application Intensity. Environ. Entomol. 2019, 48, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Saluso, A.; Xavier, L.; Silva, F.a.C.; Panizzi, A.R. An Invasive Pentatomid Pest in Argentina: Neotropical Brown Stink Bug, Euschistus heros (F.) (Hemiptera: Pentatomidae). Neotrop. Entomol. 2011, 40, 704–705. [Google Scholar]
- Panizzi, A.R. Growing Problems with Stink Bugs (Hemiptera: Heteroptera: Pentatomidae): Species Invasive to the U.S. and Potential Neotropical Invaders. Am. Entomol. 2015, 61, 223–233. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Lucini, T.; Aldrich, J.R. Dynamics in Pest Status of Phytophagous Stink Bugs in the Neotropics. Neotrop. Entomol. 2022, 51, 18–31. [Google Scholar] [CrossRef]
- Silva, F.a.C.; da Silva, J.J.; Depieri, R.A.; Panizzi, A.R. Feeding Activity, Salivary Amylase Activity, and Superficial Damage to Soybean Seed by Adult Edessa meditabunda (F.) and Euschistus heros (F.) (Hemiptera: Pentatomidae). Neotrop. Entomol. 2012, 41, 386–390. [Google Scholar] [CrossRef]
- Tessmer, M.A.; de Azevedo Kuhn, T.M.; Appezzato-da-Glória, B.; Lopes, J.R.S.; Erler, G.; Bonani, J.P. Histology of Damage Caused by Euschistus heros (F.) Nymphs in Soybean Pods and Seeds. Neotrop. Entomol. 2022, 51, 112–121. [Google Scholar] [CrossRef]
- Sosa-Gómez, D.R.; Corrêa-Ferreira, B.S.; Kraemer, B.; Pasini, A.; Husch, P.E.; Delfino Vieira, C.E.; Reis Martinez, C.B.; Negrão Lopes, I.O. Prevalence, Damage, Management and Insecticide Resistance of Stink Bug Populations (Hemiptera: Pentatomidae) in Commodity Crops. Agric. For. Entomol. 2020, 22, 99–118. [Google Scholar] [CrossRef]
- Somavilla, J.C.; da S Gubiani, P.; Reis, A.C.; Führ, F.M.; Machado, E.P.; Bernardi, O. Susceptibility of Euschistus heros and Dichelops furcatus (Hemiptera: Pentatomidae) to Insecticides Determined from Topical Bioassays and Diagnostic Doses for Resistance Monitoring of E. heros in Brazil. Crop Prot. 2020, 138, 105319. [Google Scholar] [CrossRef]
- Sosa-Gomez, D.R.; Corso, I.C.; Morales, L. Insecticide Resistance to Endosulfan, Monocrotophos and Metamidophos in the Neotropical Brown Stink Bug, Euschistus heros (F.). Neotrop. Entomol. 2001, 30, 317–320. [Google Scholar] [CrossRef]
- Sosa-Gómez, D.R.; Silva, J.J.d. Neotropical Brown Stink Bug (Euschistus heros) Resistance to Methamidophos in Paraná, Brazil. Pesqui. Agropecuária Bras. 2010, 45, 767–769. [Google Scholar] [CrossRef]
- Guedes, R.N.C. Insecticide Resistance, Control Failure Likelihood and the First Law of Geography. Pest Manag. Sci. 2017, 73, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Tuelher, E.S.; da Silva, É.H.; Rodrigues, H.S.; Hirose, E.; Guedes, R.N.C.; Oliveira, E.E. Area-Wide Spatial Survey of the Likelihood of Insecticide Control Failure in the Neotropical Brown Stink Bug Euschistus heros. J. Pest Sci. 2018, 91, 849–859. [Google Scholar] [CrossRef]
- Steinhaus, E.A.; Warpechowski, L.F.; Braga, L.E.; Morin, M.; Tenório, C.; Boff, J.S.; Bernardi, O.; Farias, J.R. Intra- and Interspecific Variation in the Susceptibility to Insecticides of Stink Bugs (Hemiptera: Pentatomidae) That Attack Soybean and Maize in Southern Brazil. J. Econ. Entomol. 2022, 115, 631–636. [Google Scholar] [CrossRef]
- Tibola, C.M.; Silva, L.; Sgubin, F.; Omoto, C. Monitoring Resistance of Euschistus heros (Fabricius) (Hemiptera: Pentatomidae) to Insecticides by Using Encapsulated Artificial Diet Bioassay. Insects 2021, 12, 599. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Nauen, R. IRAC: Mode of Action Classification and Insecticide Resistance Management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef]
- Cole, L.M.; Nicholson, R.A.; Casida, J.E. Action of Phenylpyrazole Insecticides at the GABA-Gated Chloride Channel. Pestic. Biochem. Physiol. 1993, 46, 47–54. [Google Scholar] [CrossRef]
- Knipple, D.C.; Soderlund, D.M. The Ligand-Gated Chloride Channel Gene Family of Drosophila melanogaster. Pestic. Biochem. Physiol. 2010, 97, 140–148. [Google Scholar] [CrossRef]
- Ozoe, Y. Chapter Four—γ-Aminobutyrate- and Glutamate-Gated Chloride Channels as Targets of Insecticides. In Advances in Insect Physiology; Cohen, E., Ed.; Target Receptors in the Control of Insect Pests: Part I; Academic Press: Cambridge, MA, USA, 2013; Volume 44, pp. 211–286. [Google Scholar]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D.; et al. Insecticides, Biologics and Nematicides: Updates to IRAC’s Mode of Action Classification—A Tool for Resistance Management. Pestic. Biochem. Physiol. 2020, 167, 104587. [Google Scholar] [CrossRef]
- Casida, J.E.; Durkin, K.A. Neuroactive Insecticides: Targets, Selectivity, Resistance, and Secondary Effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Roush, R.T.; Mortlock, D.; Dively, G.P. Isolation of Dieldrin Resistance from Field Populations of Drosophila melanogaster (Diptera: Drosophilidae). J. Econ. Entomol. 1990, 83, 1733–1737. [Google Scholar] [CrossRef] [PubMed]
- ffrench-Constant, R.H.; Rocheleau, T.A.; Steichen, J.C.; Chalmers, A.E. A Point Mutation in a Drosophila GABA Receptor Confers Insecticide Resistance. Nature 1993, 363, 449–451. [Google Scholar] [CrossRef]
- Le Goff, G.; Hamon, A.; Bergé, J.-B.; Amichot, M. Resistance to Fipronil in Drosophila Simulans: Influence of Two Point Mutations in the RDL GABA Receptor Subunit. J. Neurochem. 2005, 92, 1295–1305. [Google Scholar] [CrossRef]
- Nakao, T. Mechanisms of Resistance to Insecticides Targeting RDL GABA Receptors in Planthoppers. NeuroToxicology 2017, 60, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Garrood, W.T.; Zimmer, C.T.; Gutbrod, O.; Lüke, B.; Williamson, M.S.; Bass, C.; Nauen, R.; Emyr Davies, T.G. Influence of the RDL A301S Mutation in the Brown Planthopper Nilaparvata Lugens on the Activity of Phenylpyrazole Insecticides. Pestic. Biochem. Physiol. 2017, 142, 1–8. [Google Scholar] [CrossRef]
- Wolff, M.A.; Wingate, V.P. Characterization and Comparative Pharmacological Studies of a Functional Gamma-Aminobutyric Acid (GABA) Receptor Cloned from the Tobacco Budworm, Heliothis virescens (Noctuidae: Lepidoptera). Invertebr. Neurosci. 1998, 3, 305–315. [Google Scholar] [CrossRef]
- Remnant, E.J.; Morton, C.J.; Daborn, P.J.; Lumb, C.; Yang, Y.T.; Ng, H.L.; Parker, M.W.; Batterham, P. The Role of Rdl in Resistance to Phenylpyrazoles in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2014, 54, 11–21. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, X.; Yang, Y.; Li, H.; Wang, X.; Yang, B.; Zhang, J.; Li, C.; Millar, N.S.; Liu, Z. Synergistic and Compensatory Effects of Two Point Mutations Conferring Target-Site Resistance to Fipronil in the Insect GABA Receptor RDL. Sci. Rep. 2016, 6, 32335. [Google Scholar] [CrossRef]
- Moreira, R.P.; Palharini, R.B.; Massoli, G.S.; Diniz, L.H.M.; Godoy, D.N.; Warpechowski, L.F.; Steinhaus, E.A.; Stürmer, G.R.; Bernardi, O. Geographic and Interspecific Variation in Susceptibility of Euschistus heros and Diceraeus furcatus (Hemiptera: Pentatomidae) to Selected Insecticides in Southern Brazil. Crop Prot. 2024, 179, 106625. [Google Scholar] [CrossRef]
- Carvalho, R.; Okuma, D.; Bernardi, O.; Nauen, R. The Present and Future of Chemical Control to Manage Stink Bugs in Brazil. In Stink Bugs (Hemiptera: Pentatomidae) Research and Management: Recent Advances and Case Studies from Brazil, Europe, and USA; Bueno, A.F., Panizzi, A.R., Eds.; Springer Nature: Cham, Switzerland, 2024; pp. 199–212. ISBN 978-3-031-69742-5. [Google Scholar]
- Sosa-Gómez, D.R.; Bernardi, O. Insecticide Resistance Management of Stink Bugs. In Stink Bugs (Hemiptera: Pentatomidae) Research and Management: Recent Advances and Case Studies from Brazil, Europe, and USA; Bueno, A.F., Panizzi, A.R., Eds.; Springer Nature: Cham, Switzerland, 2024; pp. 241–254. ISBN 978-3-031-69742-5. [Google Scholar]
- Nakao, T.; Naoi, A.; Kawahara, N.; Hirase, K. Mutation of the GABA Receptor Associated with Fipronil Resistance in the Whitebacked Planthopper, Sogatella furcifera. Pestic. Biochem. Physiol. 2010, 97, 262–266. [Google Scholar] [CrossRef]
- Guest, M.; Goodchild, J.A.; Bristow, J.A.; Flemming, A.J. RDL A301S Alone Does Not Confer High Levels of Resistance to Cyclodiene Organochlorine or Phenyl Pyrazole Insecticides in Plutella Xylostella. Pestic. Biochem. Physiol. 2019, 158, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Nakao, T.; Banba, S.; Nomura, M.; Hirase, K. Meta-Diamide Insecticides Acting on Distinct Sites of RDL GABA Receptor from Those for Conventional Noncompetitive Antagonists. Insect Biochem. Mol. Biol. 2013, 43, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Pym, A.; Garrood, W.T.; Troczka, B.J.; Zimmer, C.T.; Davies, T.G.E.; Nauen, R.; O’Reilly, A.O.; Bass, C. P450 Gene Duplication and Divergence Led to the Evolution of Dual Novel Functions and Insecticide Cross-Resistance in the Brown Planthopper Nilaparvata Lugens. PLoS Genet. 2022, 18, e1010279. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Roush, R.T. Gene Mapping and Cross-Resistance in Cyclodiene Insecticide-Resistant Drosophila melanogaster (Mg.). Genet. Res. 1991, 57, 17–21. [Google Scholar] [CrossRef]
- Remnant, E.J.; Good, R.T.; Schmidt, J.M.; Lumb, C.; Robin, C.; Daborn, P.J.; Batterham, P. Gene Duplication in the Major Insecticide Target Site, Rdl, in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2013, 110, 14705–14710. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Bass, C. Does Resistance Really Carry a Fitness Cost? Curr. Opin. Insect Sci. 2017, 21, 39–46. [Google Scholar] [CrossRef]
- Sosa-Gómez, D.R.; Da Silva, J.J.; de Oliveira Negrao Lopes, I.; Corso, I.C.; Almeida, A.M.R.; Piubelli De Moraes, G.C.; Baur, M.E. Insecticide Susceptibility of Euschistus heros (Heteroptera: Pentatomidae) in Brazil. J. Econ. Entomol. 2009, 102, 1209–1216. [Google Scholar] [CrossRef]
- Qiao, X.; Zhou, T.; Zhang, J.; Zhang, L.; Lu, Y.; Huang, J. Functional Validation of A2′N Mutation of the RDL GABA Receptor against Fipronil via Molecular Modeling and Genome Engineering in Drosophila. Pest Manag. Sci. 2024, 80, 1924–1929. [Google Scholar] [CrossRef]
- Boff, J.S.; Reis, A.C.; Patricia, d.S.G.; Pretto, V.E.; Garlet, C.G.; Melo, A.A.; Bernardi, O. The Effect of Synergistic Compounds on the Susceptibility of Euschistus heros (Hemiptera: Pentatomidae) and Chrysodeixis includens (Lepidoptera: Noctuidae) to Pyrethroids. Environ. Entomol. 2022, 51, 421–429. [Google Scholar] [CrossRef]
- Souza, L.A.; Barbosa, J.C.; Grigolli, J.F.J.; Fraga, D.F.; Maldonado, W.; Busoli, A.C. Spatial Distribution of Euschistus heros (F.) (Hemiptera: Pentatomidae) in Soybean. Neotrop. Entomol. 2013, 42, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Engel, E.; Pasini, M.P.B.; Lúcio, A.D.C. Spatial Distribution and Sample Size to Estimate Euschistus heros Population Density in Wild Plants during Off-Season. J. Plant Dis. Prot. 2021, 128, 1073–1080. [Google Scholar] [CrossRef]
- Fernandes, M.G.; Costa, E.N.; Cavada, L.H.; Mota, T.A.; da Fonseca, P.R.B. Spatial Distribution and Sampling Plan of the Phytophagous Stink Bug Complex in Different Soybean Production Systems. J. Appl. Entomol. 2019, 143, 236–249. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | Size (bp) | |
|---|---|---|---|
| PCR/Sanger | Eh—PF | ATAAGGGTAATGGAGACGG | 405 |
| Eh—PR | AACTAGCAAAGGAGAAAAGG | ||
| qPCR | A301S Eh—PF | CGGGCTCATCGTCATCATCA | 20 |
| A301S Eh—PR | GCGGCGTTAGTAGATGACATG | 21 | |
| Sus A301S Eh | 5′-[VIC]AGTTGCCCTCGGAGTG[MGBNFQ]-3′ | 16 | |
| Res A301S Eh | 5′-[FAM]AGTTTCCCTCGGAGTG[MGBNFQ]-3′ | 16 |
| Season | Population | Ho | He | p-Value |
|---|---|---|---|---|
| 2021/22 | Deciolândia—MS | 0.150 | 0.358 | 0.011 |
| Jataí—GO | 0.800 | 0.513 | 0.007 | |
| 2022/23 | Edeia—GO | 0.657 | 0.476 | 0.000 |
| Campo Grande—MS | 0.657 | 0.502 | 0.000 | |
| 2023/24 | Corbélia—PR | 0.658 | 0.474 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuenca, A.C.P.; Lueke, B.; Vicentini, R.; Nauen, R. The Frequency and Spread of a GABA-Gated Chloride Channel Target-Site Mutation and Its Impact on the Efficacy of Ethiprole Against Neotropical Brown Stink Bug, Euschistus heros (Hemiptera: Pentatomidae). Insects 2025, 16, 422. https://doi.org/10.3390/insects16040422
Cuenca ACP, Lueke B, Vicentini R, Nauen R. The Frequency and Spread of a GABA-Gated Chloride Channel Target-Site Mutation and Its Impact on the Efficacy of Ethiprole Against Neotropical Brown Stink Bug, Euschistus heros (Hemiptera: Pentatomidae). Insects. 2025; 16(4):422. https://doi.org/10.3390/insects16040422
Chicago/Turabian StyleCuenca, Ana C. P., Bettina Lueke, Renato Vicentini, and Ralf Nauen. 2025. "The Frequency and Spread of a GABA-Gated Chloride Channel Target-Site Mutation and Its Impact on the Efficacy of Ethiprole Against Neotropical Brown Stink Bug, Euschistus heros (Hemiptera: Pentatomidae)" Insects 16, no. 4: 422. https://doi.org/10.3390/insects16040422
APA StyleCuenca, A. C. P., Lueke, B., Vicentini, R., & Nauen, R. (2025). The Frequency and Spread of a GABA-Gated Chloride Channel Target-Site Mutation and Its Impact on the Efficacy of Ethiprole Against Neotropical Brown Stink Bug, Euschistus heros (Hemiptera: Pentatomidae). Insects, 16(4), 422. https://doi.org/10.3390/insects16040422