
Cuticular Hydrocarbons of Six Geographic Populations of Ips subelongauts in Northeastern China: Similarities and Evolutionary Hints


Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Sources of I. subelongatus
2.2. Chemicals
2.3. Sampling of Cuticular Hydrocarbons
2.4. DMDS Derivatization Reaction
2.5. Cuticular Hydrocarbon Analyses with GC–MS
2.6. Data Analysis
3. Results
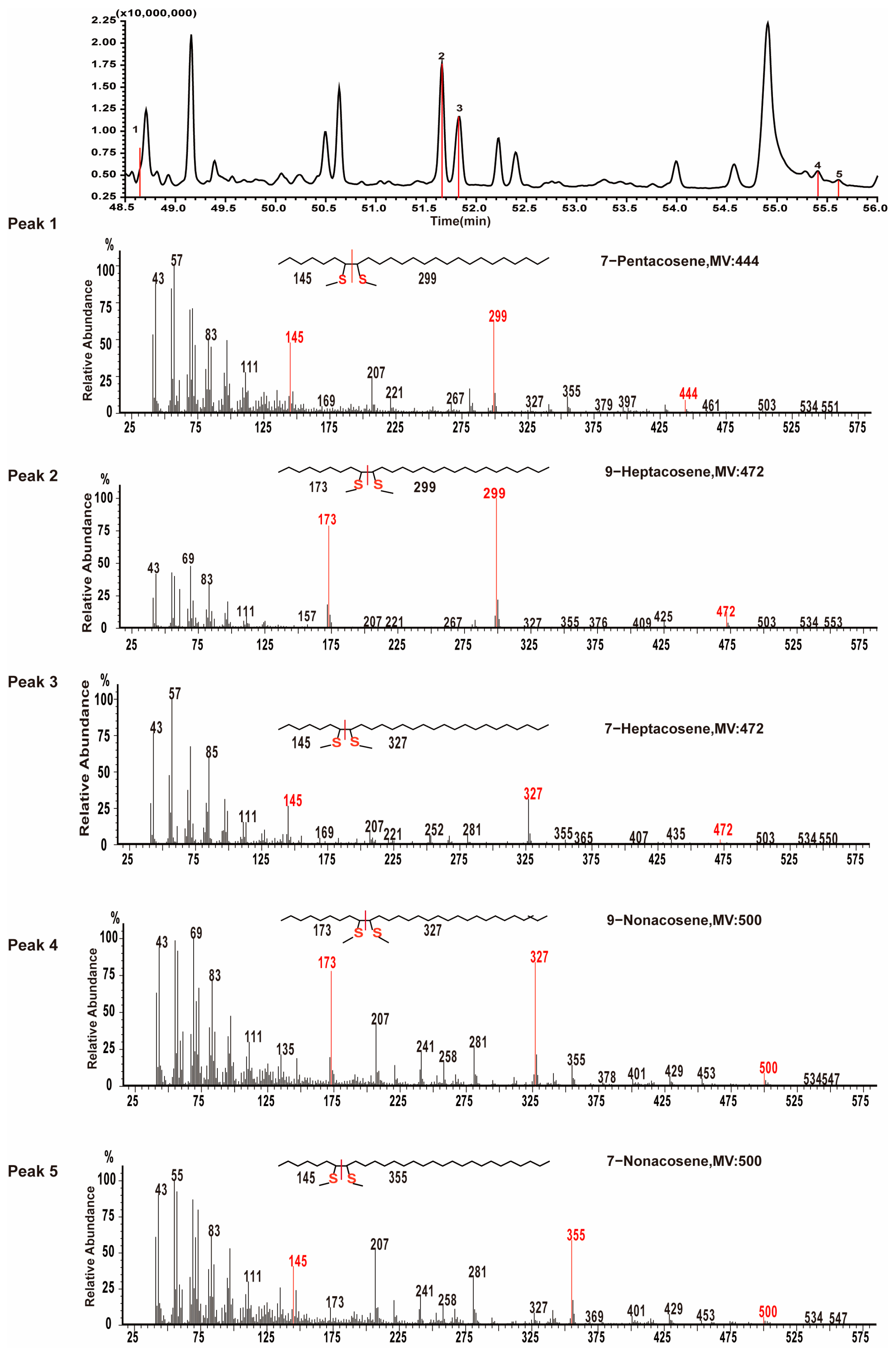
3.1. Hydrocarbon Identification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GC Peak No. | tR/min | KIs | Cuticular Hydrocarbon | Diagnostic EI Ions | % Total Hydrocarbons (Mean ± SE) | |
|---|---|---|---|---|---|---|
| Male (N = 30) | Female (N = 28) | |||||
| 1 | 34.04 | 2480 | #7-pentacosene (7-C25:1) | 83, 97, 111, 350 | 0.6 ± 0.1 | 0.6 ± 0.1 |
| 2 | 34.29 | 2500 | ※pentacosane (n-C25) | 352 | 17.5 ± 1.6 ** | 11.1 ± 1.4 |
| 3 | 35.21 | 2572 | 3-methyl-pentacosane (3-meC25) | 337, 366 | 1.4 ± 0.1 | 1.3 ± 0.1 |
| 4 | 35.56 | 2600 | hexacosane (n-C26) | 366 | 3.1 ± 0.4 | 2.7 ± 0.4 |
| 5 | 36.27 | 2657 | 4-methyl-hexacosane (4-meC26) | 43, 71, 337, 380 | 0.4 ± 0.0 | 0.6 ± 0.1 |
| 6 | 36.52 | 2677 | #9-heptacosene (9-C27:1) | 83, 97, 111, 378 | 16.8 ± 1.7 | 15.8 ± 2.0 |
| 7 | 36.6 | 2684 | #7-heptacosene (7-C27:1) | 83, 97, 111, 378 | 5.3 ± 0.4 | 4.8 ± 0.6 |
| 8 | 36.8 | 2700 | ※heptacosane (n-C27) | 380 | 21.8 ± 1.6 | 19.5 ± 1.7 |
| 9 | 37.19 | 2733 | 11-methyl-heptacosane (11-meC27) | 168, 252 | 0.7 ± 0.1 | 0.9 ± 0.1 |
| 10 | 37.29 | 2741 | 7-methyl-heptacosane (7-meC27) | 112, 309 | 0.2 ± 0.0 ** | 0.4 ± 0.0 |
| 11 | 37.39 | 2750 | 5-methyl-heptacosane (5-meC27) | 85, 337 | 2.0 ± 0.1 | 2.4 ± 0.2 |
| 12 | 37.68 | 2774 | 3-methyl-heptacosane (3-meC27) | 365, 394 | 7.0 ± 0.5 | 7.9 ± 0.5 |
| 13 | 37.99 | 2800 | octacosane (n-C28) | 394 | 1.7 ± 0.2 | 2.0 ± 0.2 |
| 14 | 38.09 | 2809 | 3,7-dimethyl-heptacosane (3,7-dimeC27) | 393, 379, 126, 308 | 1.6 ± 0.1 | 1.7 ± 0.1 |
| 15 | 38.67 | 2858 | 4-methyl-octacosane (4-meC28) | 43, 71, 365 | 1.1 ± 0.1 * | 1.7 ± 0.2 |
| 16 | 38.92 | 2879 | #9-nonacosene (9-C29:1) | 406, 83, 97, 111 | 2.0 ± 0.2 | 2.3 ± 0.2 |
| 17 | 38.99 | 2885 | #7-nonacosene (7-C29:1) | 406, 71, 83, 97, 111 | 1.4 ± 0.1 | 1.6 ± 0.1 |
| 18 | 39.16 | 2900 | ※nonacosane (n-C29) | 408 | 2.4 ± 0.1 | 2.8 ± 0.2 |
| 19 | 39.5 | 2930 | 13-methyl-nonacosane (13-meC29) | 196, 224, 252, 407 | 1.1 ± 0.3 | 1.6 ± 0.4 |
| 20 | 39.6 | 2938 | 7-methyl-nonacosane (7-meC29) | 112, 337, 407 | 1.2 ± 0.2 * | 2.0 ± 0.3 |
| 21 | 39.71 | 2948 | 5-methyl-nonacosane (5-meC29) | 85, 365, 407 | 1.2 ± 0.2 | 1.6 ± 0.2 |
| 22 | 39.83 | 2958 | 9,13-dimethyl-nonacosane (9,13-dimeC29) | 421, 323, 252, 211, 140 | 0.8 ± 0.2 | 1.1 ± 0.3 |
| 23 | 40.04 | 2977 | 5,X-dimethyl-nonacosane (5,X-dimeC29) | 85, 196, 211, 379, 421 | 2.5 ± 0.7 | 4.2 ± 0.9 |
| 24 | 40.31 | 3000 | ※triacontane (n-C30) | 422 | 0.5 ± 0.1 | 0.5 ± 0.1 |
| 25 | 41.15 | 3073 | 7-hentriacontene (7-C31:1) | 83, 97, 111, 434 | 0.5 ± 0.1 | 0.5 ± 0.1 |
| 26 | 41.71 | 3121 | 15-methyl-hentriacontane (15-meC31) | 196, 224, 252 | 1.6 ± 0.5 | 2.6 ± 0.6 |
| 27 | 41.84 | 3133 | 7-methyl-hentriacontane (7-meC31) | 112, 365, 436 | 0.4 ± 0.1 | 0.7 ± 0.2 |
| 28 | 42.04 | 3150 | 11,15-dimethyl-hentriacontane (11,15-meC31) | 168, 239, 252, 323 | 0.7 ± 0.2 | 1.1 ± 0.3 |
| 29 | 42.19 | 3163 | 4-methyl-hentriacontane (4-meC31) | 71, 379, 450 | 0.9 ± 0.2 | 1.5 ± 0.4 |
| 30 | 42.3 | 3173 | 5,17-dimethyl-hentriacontane (5,17-dimeC31) | 85, 126, 168, 225, 267, 407 | 1.7 ± 0.5 | 2.7 ± 0.7 |
| GC Peak No. | CHCs | % Total Hydrocarbons (Mean ± SE) | |||||
|---|---|---|---|---|---|---|---|
| MJHL N = 10 | WDLN N = 12 | GHIM N = 10 | YCHL N = 8 | EDJL N = 12 | ARIM N = 6 | ||
| 1 | #7-C25:1 | 0.7 ± 0.1 a | 0.9 ± 0.1 a | 0.1 ± 0.1 b | 0.1 ± 0.0 b | 0.7 ± 0.1 a | 0.8 ± 0.1 a |
| 2 | ※n-C25 | 12.3 ± 1.3 b | 17.1 ± 1.9 b | 6.5 ± 0.9 c | 5.2 ± 0.8 c | 26.8 ± 1.3 a | 13.4 ± 0.8 b |
| 3 | 3-meC25 | 1.7 ± 0.1 a | 1.6 ± 0.1 a | 0.8 ± 0.1 b | 0.7 ± 0.1 b | 1.7 ± 0.1 a | 1.5 ± 0.2 ab |
| 4 | n-C26 | 5.6 ± 0.5 a | 4.2 ± 0.4 a | 1.2 ± 0.2 c | 0.9 ± 0.1 c | 2.6 ± 0.1 b | 1.6 ± 0.3 bc |
| 5 | 4-meC26 | 0.7 ± 0.1 a | 0.5 ± 0.1 a | 0.4 ± 0.1 ab | 0.8 ± 0.5 ab | 0.3 ± 0.1 b | 0.5 ± 0.1 ab |
| 6 | #9-C27:1 | 13.5 ± 0.8 c | 18.6 ± 1.4 bc | 10.1 ± 1.4 c | 4.4 ± 0.4 d | 17.6 ± 0.7 b | 39.4 ± 2.2 a |
| 7 | #7-C27:1 | 5.3 ± 0.5 a | 7.4 ± 0.7 a | 2.4 ± 0.3 b | 1.8 ± 0.2 b | 6.1 ± 0.5 a | 6.7 ± 0.6 a |
| 8 | ※n-C27 | 23.5 ± 1.8 b | 27.6 ± 1.2 a | 14.1 ± 1.4 c | 8.1 ± 1.1 d | 29.3 ± 1.0 a | 12.0 ± 0.9 cd |
| 9 | 11-meC27 | 1.2 ± 0.2 ab | 0.3 ± 0.1 bc | 1.5 ± 0.1 a | 1.7 ± 0.2 a | 0.02 ± 0.0 c | 0.5 ± 0.1 b |
| 10 | 7-meC27 | 0.3 ± 0.1 b | 0.1 ± 0.1 c | 0.5 ± 0.1 ab | 0.5 ± 0.1 a | 0.1 ± 0.1 c | 0.3 ± 0.1 b |
| 11 | 5-meC27 | 2.9 ± 0.2 a | 1.8 ± 0.2 bc | 2.7 ± 0.2 a | 2.7 ± 0.3 a | 1.3 ± 0.1 c | 1.9 ± 0.2 b |
| 12 | 3-meC27 | 11.2 ± 0.3 a | 7.3 ± 0.4 bc | 8.5 ± 0.5 b | 7.0 ± 0.8 bc | 4.7 ± 0.4 c | 5.0 ± 0.6 c |
| 13 | n-C28 | 3.3 ± 0.3 a | 2.2 ± 0.2 b | 1.5 ± 0.2 bc | 2.2 ± 0.5 abcd | 0.9 ± 0.1 c | 0.5 ± 0.1 d |
| 14 | 3,7-dimeC27 | 2.2 ± 0.2 a | 1.2 ± 0.1 b | 2.2 ± 0.3 a | 1.9 ± 0.1 ab | 1.3 ± 0.1 ab | 1.6 ± 0.2 ab |
| 15 | 4-meC28 | 1.7 ± 0.1 b | 1.0 ± 0.2 c | 1.8 ± 0.1 b | 3.2 ± 0.3 a | 0.2 ± 0.1 d | 1.1 ± 0.1 c |
| 16 | #9-C29:1 | 1.5 ± 0.2 d | 2.0 ± 0.2 c | 2.8 ± 0.2 b | 2.2 ± 0.2 bc | 1.7 ± 0.1 bcd | 3.5 ± 0.3 a |
| 17 | #7-C29:1 | 1.2 ± 0.1 bc | 1.3 ± 0.1 b | 2.1 ± 0.2 a | 2.3 ± 0.1 a | 0.8 ± 0.1 c | 1.8 ± 0.3 a |
| 18 | ※n-C29 | 3.2 ± 0.3 a | 3.1 ± 0.3 a | 2.4 ± 0.2 a | 2.4 ± 0.2 a | 2.7 ± 0.2 a | 1.0 ± 0.1 b |
| 19 | 13-meC29 | 0.6 ± 0.2 b | nd | 3.4 ± 0.3 a | 4.3 ± 0.3 a | nd | 0.6 ± 0.1 b |
| 20 | 7-meC29 | 1.4 ± 0.2 c | 0.6 ± 0.4 c | 2.9 ± 0.3 b | 4.2 ± 0.2 a | 0.1 ± 0.1 d | 1.0 ± 0.1 c |
| 21 | 5-meC29 | 1.7 ± 0.1 c | 0.8 ± 0.2 d | 2.2 ± 0.2 b | 3.1 ± 0.2 a | 0.2 ± 0.1 e | 0.6 ± 0.1 de |
| 22 | 9,13-dimeC29 | nd | nd | 2.5 ± 0.3 a | 3.4 ± 0.4 a | nd | 0.2 ± 0.1 b |
| 23 | 5,X-dimeC29 | 1.5 ± 0.4 b | 0.3 ± 0.1 c | 8.2 ± 1.0 a | 10.6 ± 0.3 a | 0.1 ± 0.1 c | 1.4 ± 0.2 b |
| 24 | ※n-C30 | 0.8 ± 0.1 a | nd | 1.2 ± 0.2 a | 0.8 ± 0.1 a | nd | 0.4 ± 0.1 b |
| 25 | 9-C31:1 | 0.3 ± 0.1 b | 0.1 ± 0.1 b | 0.9 ± 0.1 a | 1.3 ± 0.19 a | 0.2 ± 0.1 b | 0.5 ± 0.2 ab |
| 26 | 15-meC31 | 0.7 ± 0.1 b | nd | 4.3 ± 0.4 a | 8.0 ± 1.08 a | 0.3 ± 0.1 b | 0.7 ± 0.1 b |
| 27 | 7-meC31 | nd | nd | 1.0 ± 0.1 a | 2.1 ± 0.14 a | 0.2 ± 0.1 b | 0.1 ± 0.1 b |
| 28 | 11,15-dimeC31 | nd | nd | 2.4 ± 0.3 a | 3.4 ± 0.36 a | nd | 0.2 ± 0.0 b |
| 29 | 4-meC31 | 0.6 ± 0.1 b | nd | 3.5 ± 0.5 a | 3.2 ± 0.20 a | nd | 0.5 ± 0.1 b |
| 30 | 5,17-dimeC31 | 0.5 ± 0.1 b | nd | 6.0 ± 0.6 a | 7.2 ± 0.3 a | nd | 0.7 ± 0.1 b |
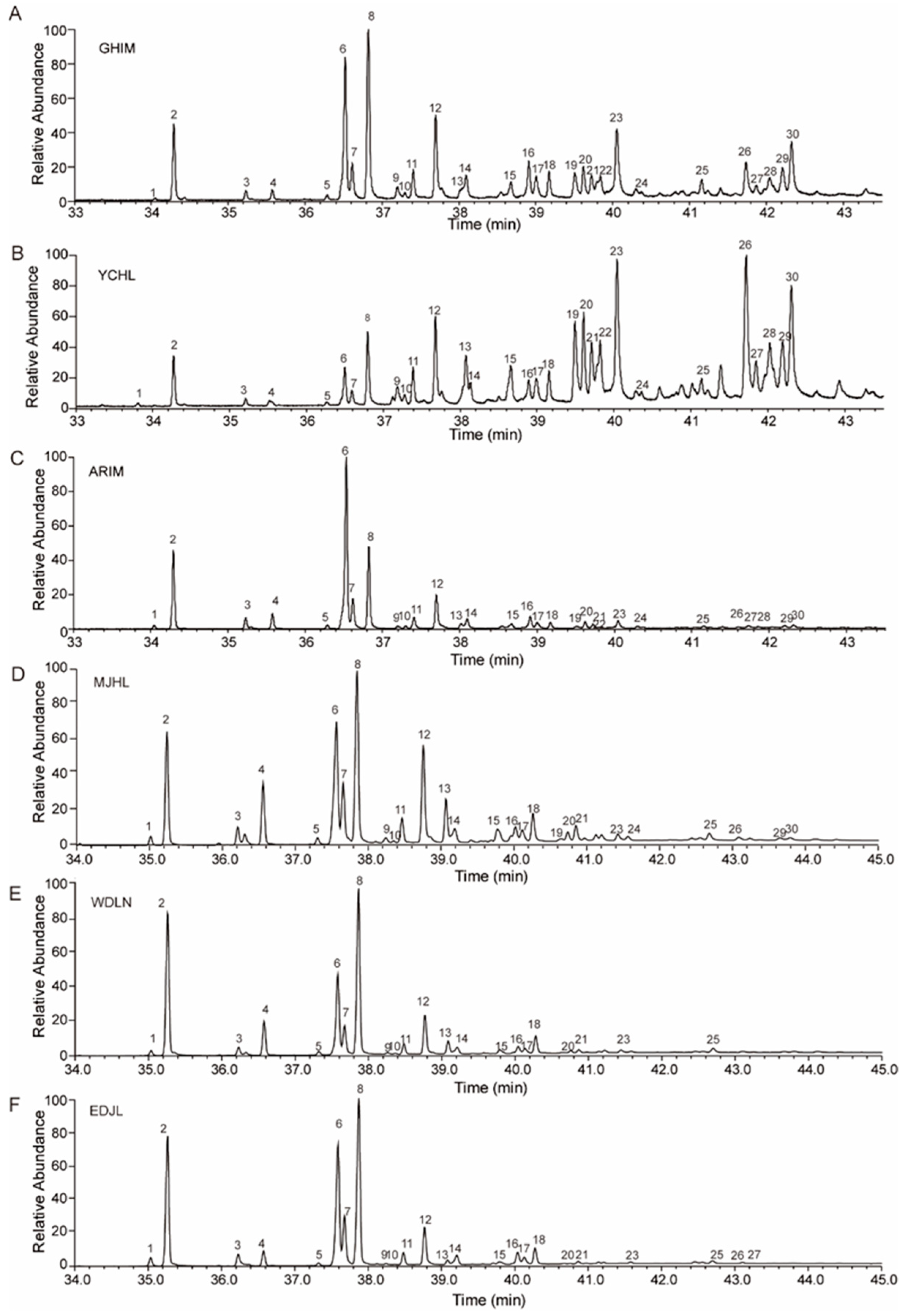
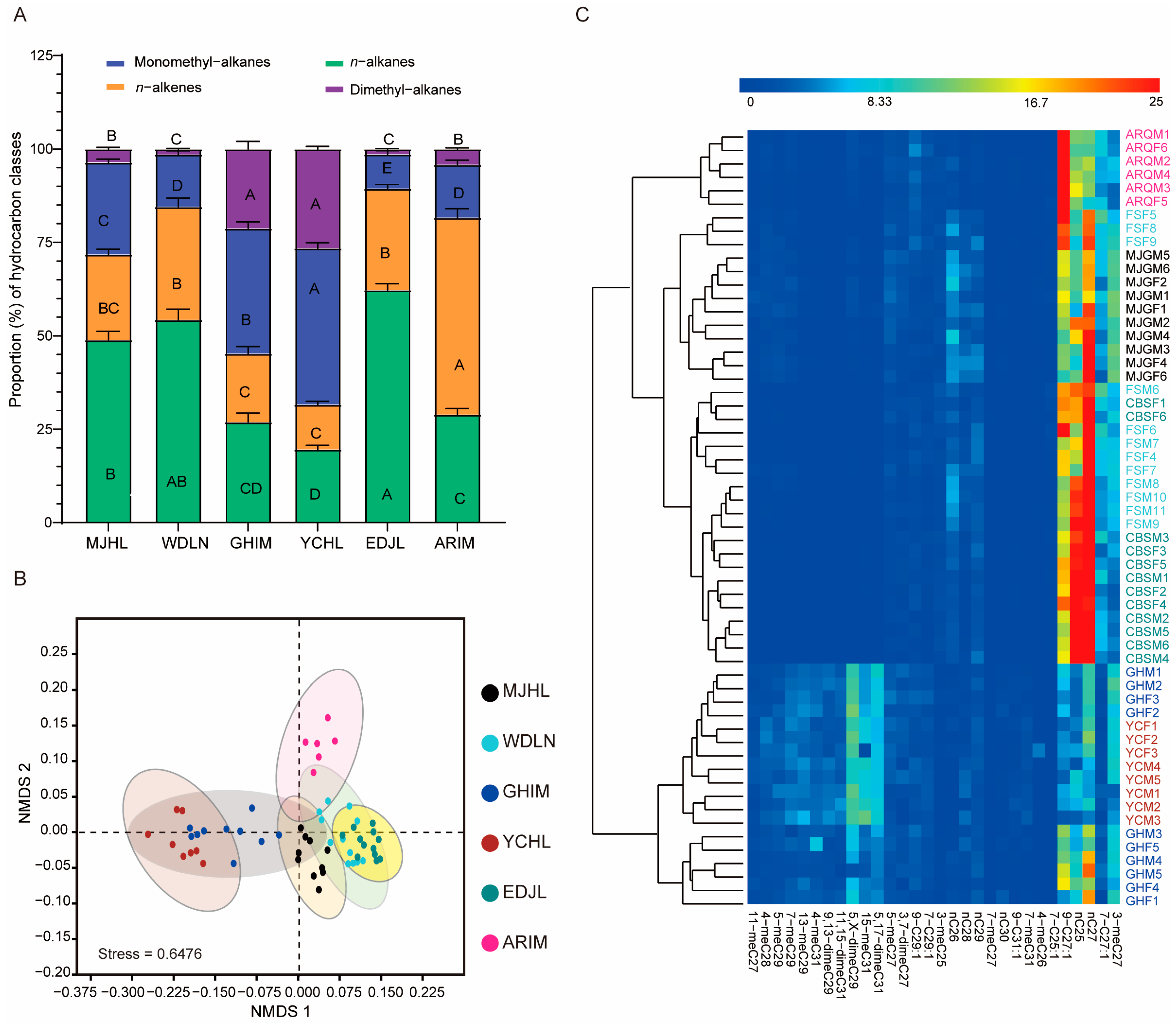
3.2. Percentage Composition of Cuticular Hydrocarbons in I. subelongatus
3.3. Population Differences in Cuticular Hydrocarbon Profiles
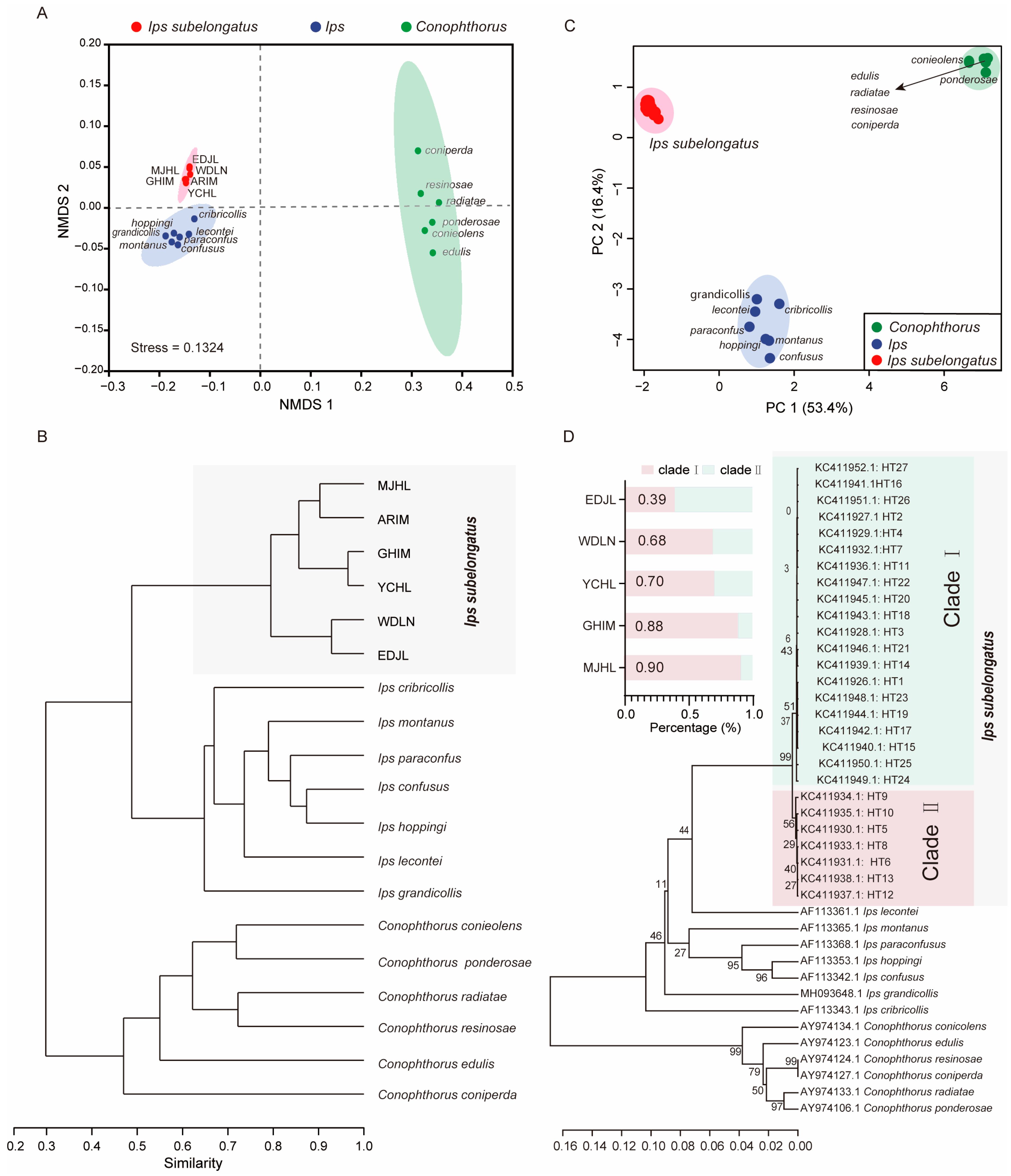
3.4. Cuticular Hydrocarbons as Taxonomic Traits
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Blomquist, G.J.; Tittiger, C.; Jurenka, R. Cuticular hydrocarbons and pheromones of Arthropods. In Hydrocarbons, Oils and Lipids: Diversity, Origin, Chemistry and Fate; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–32. [Google Scholar]
- Gibbs, A.G. Water-Proofing Properties of Cuticular Lipids1. Am. Zool. 1998, 38, 471–482. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Tittiger, C.; MacLean, M.; Blomquist, G.J. Hydrocarbon pheromone production in insects. In Insect Pheromone Biochemistry and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 205–235. [Google Scholar]
- Wang, Z.; Andika, I.P.; Chung, H. Regulation of insect cuticular hydrocarbon biosynthesis. Curr. Opin. Insect Sci. 2025, 67, 101287. [Google Scholar] [CrossRef]
- Peterson, M.A.; Dobler, S.; Larson, E.L.; Juárez, D.; Schlarbaum, T.; Monsen, K.J.; Francke, W. Profiles of cuticular hydrocarbons mediate male mate choice and sexual isolation between hybridising Chrysochus (Coleoptera: Chrysomelidae). Chemoecology 2007, 17, 87–96. [Google Scholar]
- Zhang, B.; Xue, H.J.; Song, K.Q.; Liu, J.; Li, W.Z.; Nie, R.E.; Yang, X.K. Male mate recognition via cuticular hydrocarbons facilitates sexual isolation between sympatric leaf beetle sister species. J. Insect Physiol. 2014, 70, 15–21. [Google Scholar] [CrossRef]
- Fang, J.X.; Chen, D.F.; Shi, X.; Zhang, S.F.; Liu, F.; Shen, W.X.; Jia, C.Y.; Ma, S.C.; Zhang, Z.; Kong, X.B. Solid-phase microextraction and cuticular hydrocarbon differences related to reproductive activity in juniper bark borer Semanotus bifasciatus Motschulsky. J. Syst. Evol. 2023, 61, 498–505. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [PubMed]
- Moore, H.E.; Hall, M.J.R.; Drijfhout, F.P.; Cody, R.B.; Whitmore, D. Cuticular hydrocarbons for identifying Sarcophagidae (Diptera). Sci. Rep. 2021, 11, 7732. [Google Scholar] [CrossRef] [PubMed]
- Koedam, D.; Morgan, E.; Nunes, T.M.; Patricio, E.; Imperatriz Fonseca, V.L. Selective preying of the sphecid wasp Trachypus boharti on the meliponine bee Scaptotrigona postica: Potential involvement of caste–specific cuticular hydrocarbons. Physiol. Entomol. 2010, 36, 187–193. [Google Scholar]
- Endo, S.; Itino, T. Myrmecophilous aphids produce cuticular hydrocarbons that resemble those of their tending ants. Popul. Ecol. 2012, 55, 27–34. [Google Scholar] [CrossRef]
- Binz, H.; Kraft, E.F.; Entling, M.H.; Menzel, F. Behavioral response of a generalist predator to chemotactile cues of two taxonomically distinct prey species. Chemoecology 2016, 26, 153–162. [Google Scholar] [CrossRef]
- Page, M.; Nelson, L.J.; Haverty, M.I.; Blomquist, G.J. Cuticular hydrocarbons as chemotaxonomic characters for bark beetles: Dendroctonus ponderosae, D. jeffreyi, D. brevicomis, and D. frontalis (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1990, 83, 892–901. [Google Scholar]
- Braga, M.V.; Pinto, Z.T.; de Carvalho Queiroz, M.M.; Matsumoto, N.; Blomquist, G.J. Cuticular hydrocarbons as a tool for the identification of insect species: Puparial cases from Sarcophagidae. Acta Trop. 2013, 128, 479–485. [Google Scholar] [PubMed]
- Guillem, R.M.; Drijfhout, F.P.; Martin, S.J. Species-specific cuticular hydrocarbon stability within European Myrmica Ants. J. Chem. Ecol. 2016, 42, 1052–1062. [Google Scholar]
- Johnson, J.B. Cuticular hydrocarbon profiling reveals chemotaxonomic diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biol. Life Sci. Forum 2022, 15, 5. [Google Scholar] [CrossRef]
- Drijfhout, F.P.; Kather, R.; Martin, S.J. The role of cuticular hydrocarbons in insects. In Behavioral and Chemical Ecology; Nova Science: New York, NY, USA, 2013; pp. 1–24. [Google Scholar]
- Rodrigues Méndez, M.P.; Muñoz-Rodríguez, D.; Aragón-Pech, R.A.J.; Macías Macías, J.O.; Quezada-Euán, J.J.G. Cuticular hydrocarbon profiles reveal geographic chemotypes in stingless bees (Hymenoptera: Meliponini). Sci. Rep. 2024, 14, 14567. [Google Scholar]
- Kather, R.; Martin, S.J. Evolution of Cuticular Hydrocarbons in the Hymenoptera: A Meta-Analysis. J. Chem. Ecol. 2015, 41, 871–883. [Google Scholar] [PubMed]
- Richter, N.; Dillon, J.T.; Rott, D.M.; Lomazzo, M.A.; Seto, C.T.; Huang, Y. Optimizing the yield of transient mono-dimethyl disulfide adducts for elucidating double bond positions of long chain alkenones. Org. Geochem. 2017, 109, 58–66. [Google Scholar]
- Marques, F.D.; McElfresh, J.S.; Millar, J.G. Kováts retention indexes of monounsaturated C12, C14, and C16 alcohols, acetates and aldehydes commonly found in lepidopteran pheromone blends. J. Braz. Chem. Soc. 2000, 11, 592–599. [Google Scholar]
- Rodríguez-León, D.S.; Uzunov, A.; Costa, C.; Elen, D.; Charistos, L.; Galea, T.; Gabel, M.; Scheiner, R.; Pinto, M.A.; Schmitt, T. Deciphering the variation in cuticular hydrocarbon profiles of six European honey bee subspecies. Ecol. Evol. 2024, 24, 131. [Google Scholar]
- Zhang, Q.H.; Byers, J.A.; Schlyter, F. Optimal attack density in the larch bark beetle, Ips cembrae (Coleoptera: Scolytidae). J. Appl. Ecol. 1992, 29, 672–678. [Google Scholar]
- Zhang, Q.H.; Schlyter, F.; Chen, G.; Wang, Y. Electrophysiological and behavioral responses of Ips subelongatus to semiochemicals from its hosts, non-hosts, and conspecifics in China. J. Chem. Ecol. 2007, 33, 391–404. [Google Scholar] [PubMed]
- Chen, D.F.; Li, Y.J.; Zhang, Q.H.; Zhang, S.F.; Wang, H.B.; Zhang, Z.; Zhao, L.L.; Kong, X.B. Population divergence of aggregation pheromone responses in Ips subelongatus in northeastern China. Insect Sci. 2016, 23, 728–738. [Google Scholar]
- Shi, X.; Zhang, S.F.; Liu, F.; Xu, F.Y.; Zhang, F.B.; Guo, X.B.; Zhang, Z.; Kong, X.B. SEM analysis of sensilla on the mouthparts and antennae of Asian larch bark beetle Ips subelongatus. Micron 2021, 140, 102976. [Google Scholar] [CrossRef]
- Song, L.W.; Zhang, Q.H.; Chen, Y.Q.; Zuo, T.T.; Ren, B.Z. Field responses of the Asian larch bark beetle, Ips subelongatus, to potential aggregation pheromone components: Disparity between two populations in northeastern China. Insect Sci. 2011, 18, 311–319. [Google Scholar]
- Moore, H.; Lutz, L.; Bernhardt, V.; Drijfhout, F.P.; Cody, R.B.; Amendt, J. Cuticular hydrocarbons for the identification and geographic assignment of empty puparia of forensically important flies. Int. J. Leg. Med. 2022, 136, 1791–1800. [Google Scholar]
- Hammer, Y.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Page, M.; Nelson, L.J.; Haverty, M.I.; Blomquist, G.J. Cuticular hydrocarbons of eight species of North American cone beetles, Conophthorus Hopkins. J. Chem. Ecol. 1990, 16, 1173–1198. [Google Scholar]
- Page, M.; Nelson, L.J.; Blomquist, G.J.; Seybold, S.J. Cuticular hydrocarbons as chemotaxonomic characters of pine engraver beetles (Ips spp.) in the grandicollis subgeneric group. J. Chem. Ecol. 1997, 23, 1053–1099. [Google Scholar]
- Ricotta, C.; Podani, J. On some properties of the Bray-Curtis dissimilarity and their ecological meaning. Ecol. Complex. 2017, 31, 201–205. [Google Scholar]
- Chen, T.; Li, Q.; Qi, G.; Gao, Y.; Zhao, C.; Lu, L. Cuticular hydrocarbon pattern as a chemotaxonomy marker to assess six species of thrips. J. Asia-Pac. Entomol. 2020, 23, 1255–1263. [Google Scholar] [CrossRef]
- Vaníčková, L.; Virgilio, M.; Tomčala, A.; Břízová, R.; Ekesi, S.; Hoskovec, M.; Kalinová, B.; Do Nascimento, R.R.; De Meyer, M. Resolution of three cryptic agricultural pests (Ceratitis fasciventris, C. anonae, C. rosa, Diptera: Tephritidae) using cuticular hydrocarbon profiling. Bull. Entomol. Res. 2014, 104, 631–638. [Google Scholar] [CrossRef]
- Barbosa, R.R.; Braga, M.V.; Blomquist, G.J.; Queiroz, M.M.d.C. Cuticular hydrocarbon profiles as a chemotaxonomic tool for three blowfly species (Diptera: Calliphoridae) of forensic interest. J. Nat. Hist. 2017, 51, 1491–1498. [Google Scholar] [CrossRef]
- Torres, C.W.; Tonione, M.A.; Ramírez, S.R.; Sapp, J.R.; Tsutsui, N.D. Genetic and chemical divergence among host races of a socially parasitic ant. Ecol. Evol. 2018, 8, 11385–11398. [Google Scholar] [CrossRef]
- Haverty, M.I.; Woodrow, R.J.; Nelson, L.J.; Grace, J.K. Cuticular hydrocarbons of termites of the Hawaiian Islands. J. Chem. Ecol. 2000, 26, 1167–1191. [Google Scholar] [CrossRef]
- Caravantes-Villatoro, L.A.; Cruz-Esteban, S.; Rojas, J.C. Cuticular hydrocarbons of Anastrepha obliqua (Diptera: Tephritidae) as influenced by extraction method, natal host, and age. Fla. Entomol. 2021, 104, 289–296. [Google Scholar] [CrossRef]
- Hais, A.; Casacci, L.P.; d’Ettorre, P.; Badía-Villas, D.; Leroy, C.; Barbero, F. Variation in Ants’ Chemical Recognition Signals across Vineyard Agroecosystems. Int. J. Mol. Sci. 2024, 25, 10407. [Google Scholar] [CrossRef]
- Kruger, E.L.; Pappas, C.D. Geographic variation of cuticular hydrocarbons among fourteen populations of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1993, 30, 544–548. [Google Scholar] [CrossRef]
- Bontonou, G.; Claude, W.T. Sexual communication in the Drosophila genus. Insects 2014, 5, 20. [Google Scholar] [CrossRef]
- Kula, C.; Amendt, J.; Drijfhout, F.P.; Moore, H.E.; Brundage, A. Geographical variation of cuticular hydrocarbon profiles of adult flies and empty puparia amongst three populations of Calliphora vicina (Diptera: Calliphoridae). J. Med. Entomol. 2023, 60, 14–23. [Google Scholar] [CrossRef]
- Jennings, J.H.; Etges, W.J.; Schmitt, T.; Hoikkala, A. Cuticular hydrocarbons of Drosophila montana: Geographic variation, sexual dimorphism and potential roles as pheromones. J. Insect Physiol. 2014, 61, 16–24. [Google Scholar] [CrossRef]
- Veltsos, P.; Wicker-Thomas, C.; Butlin, R.K.; Hoikkala, A.; Ritchie, M.G. Sexual selection on song and cuticular hydrocarbons in two distinct populations of Drosophila montana. Ecol. Evol. 2011, 2, 80–94. [Google Scholar]
- Bonelli, M.L.; Lorenzi, M.C.; Christidès, J.P.; Dupont, S.; Bagnères, A. Population diversity in cuticular hydrocarbons and mtDNA in a mountain social wasp. J. Chem. Ecol. 2014, 41, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Otte, T.; Hilker, M.; Geiselhardt, S. Phenotypic Plasticity of cuticular hydrocarbon profiles in insects. J. Chem. Ecol. 2018, 44, 235–247. [Google Scholar] [CrossRef]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2020, 126, 219–234. [Google Scholar]
- Page, M.; Nelson, L.J.; Forschler, B.T.; Haverty, M.I. Cuticular hydrocarbons suggest three lineages in Reticulitermes (Isoptera: Rhinotermitidae) from North America. In Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2002; Volume 131, pp. 305–324. [Google Scholar]
- Martin, S.; Drijfhout, F. A review of ant Cuticular hydrocarbons. J. Chem. Ecol. 2009, 35, 1151–1161. [Google Scholar] [CrossRef]
- Menzel, F.; Schmitt, T.; Blaimer, B.B. The evolution of a complex trait: Cuticular hydrocarbons in ants evolve independent from phylogenetic constraints. J. Evol. Biol. 2017, 30, 1372–1385. [Google Scholar]
- Copren, K.A.; Nelson, L.J.; Vargo, E.L.; Haverty, M.I. Phylogenetic analyses of mtDNA sequences corroborate taxonomic designations based on cuticular hydrocarbons in subterranean termites. Mol. Phylogenet. Evol. 2005, 35, 689–700. [Google Scholar]
- Mullen, S.P.; Millar, J.G.; Schal, C.; Shaw, K.L. Identification and characterization of cuticular hydrocarbons from a rapid species radiation of Hawaiian swordtailed crickets (Gryllidae: Trigonidiinae: Laupala). J. Chem. Ecol. 2008, 34, 198–204. [Google Scholar] [PubMed]
- Jallon, J.M.; David, J.R. Variations in cuticular hydrocarbons among the eight species of the Drosophila melanogaster subgroup. Evolution 1987, 41, 9. [Google Scholar]
- Ferveur, J.F. Cuticular hydrocarbons: Their evolution and roles in Drosophila pheromonal communication. Behav. Genet. 2005, 35, 279–295. [Google Scholar]
- Marten, A.; Kaib, M.; Brandl, R. Cuticular hydrocarbon phenotypes do not indicate cryptic species in fungus-growing termites (Isoptera: Macrotermitinae). J. Chem. Ecol. 2009, 35, 572–579. [Google Scholar] [PubMed]
- Dronnet, S.; Lohou, C.; Christides, J.P.; Bagnères, A.G. Cuticular hydrocarbon composition reflects genetic relationship among colonies of the introduced termite Reticulitermes santonensis Feytaud. J. Chem. Ecol. 2006, 32, 1027–1042. [Google Scholar] [PubMed]
- Dapporto, L.; Liebert, A.E.; Starks, P.T.; Turillazzi, S. The relationships between cuticular hydrocarbon composition, faunal assemblages, inter-island distance, and population genetic variation in Tuscan Archipelago wasps. Biochem. Syst. Ecol. 2009, 37, 341–348. [Google Scholar]
- Chen, N.; Bai, Y.; Fan, Y.L.; Liu, T.X. Solid-phase microextraction-based cuticular hydrocarbon profiling for intraspecific delimitation in Acyrthosiphon pisum. PLoS ONE 2017, 12, e0184243. [Google Scholar]
- Cunha, D.A.S.; Menezes, R.S.T.; Costa, M.A.; Lima, S.M.; Andrade, L.H.C.; Antonialli, W.F. Integrated analyses of cuticular hydrocarbons, chromosome and mtDNA in the neotropical social wasp Mischocyttarus consimilis Zikán (Hymenoptera, Vespidae). Neotrop. Entomol. 2017, 46, 642–648. [Google Scholar]
- Blomquist, G.J.; Ginzel, M.D. Chemical ecology, biochemistry, and molecular biology of insect hydrocarbons. Annu. Rev. Entomol. 2021, 66, 45–60. [Google Scholar]
- Ginzel, M.D.; Moreira, J.A.; Ray, A.M.; Millar, J.G.; Hanks, L.M. (Z)-9-Nonacosene–major Ccomponent of the contact sex pheromone of the beetle Megacyllene caryae. J. Chem. Ecol. 2006, 32, 435–451. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, C.; Liu, X.; Lin, X.; Chu, D.; Ding, J.; Kong, X.; Chen, D. Cuticular Hydrocarbons of Six Geographic Populations of Ips subelongauts in Northeastern China: Similarities and Evolutionary Hints. Insects 2025, 16, 384. https://doi.org/10.3390/insects16040384
Zhao Y, Wang C, Liu X, Lin X, Chu D, Ding J, Kong X, Chen D. Cuticular Hydrocarbons of Six Geographic Populations of Ips subelongauts in Northeastern China: Similarities and Evolutionary Hints. Insects. 2025; 16(4):384. https://doi.org/10.3390/insects16040384
Chicago/Turabian StyleZhao, Yuge, Chao Wang, Xinmeng Liu, Xu Lin, Dongdong Chu, Junyi Ding, Xiangbo Kong, and Dafeng Chen. 2025. "Cuticular Hydrocarbons of Six Geographic Populations of Ips subelongauts in Northeastern China: Similarities and Evolutionary Hints" Insects 16, no. 4: 384. https://doi.org/10.3390/insects16040384
APA StyleZhao, Y., Wang, C., Liu, X., Lin, X., Chu, D., Ding, J., Kong, X., & Chen, D. (2025). Cuticular Hydrocarbons of Six Geographic Populations of Ips subelongauts in Northeastern China: Similarities and Evolutionary Hints. Insects, 16(4), 384. https://doi.org/10.3390/insects16040384