
Modeling the Potential Distribution and Future Dynamics of Important Vector Culex tritaeniorhynchus Under Climate Change Scenarios in China


Simple Summary
Abstract
1. Introduction
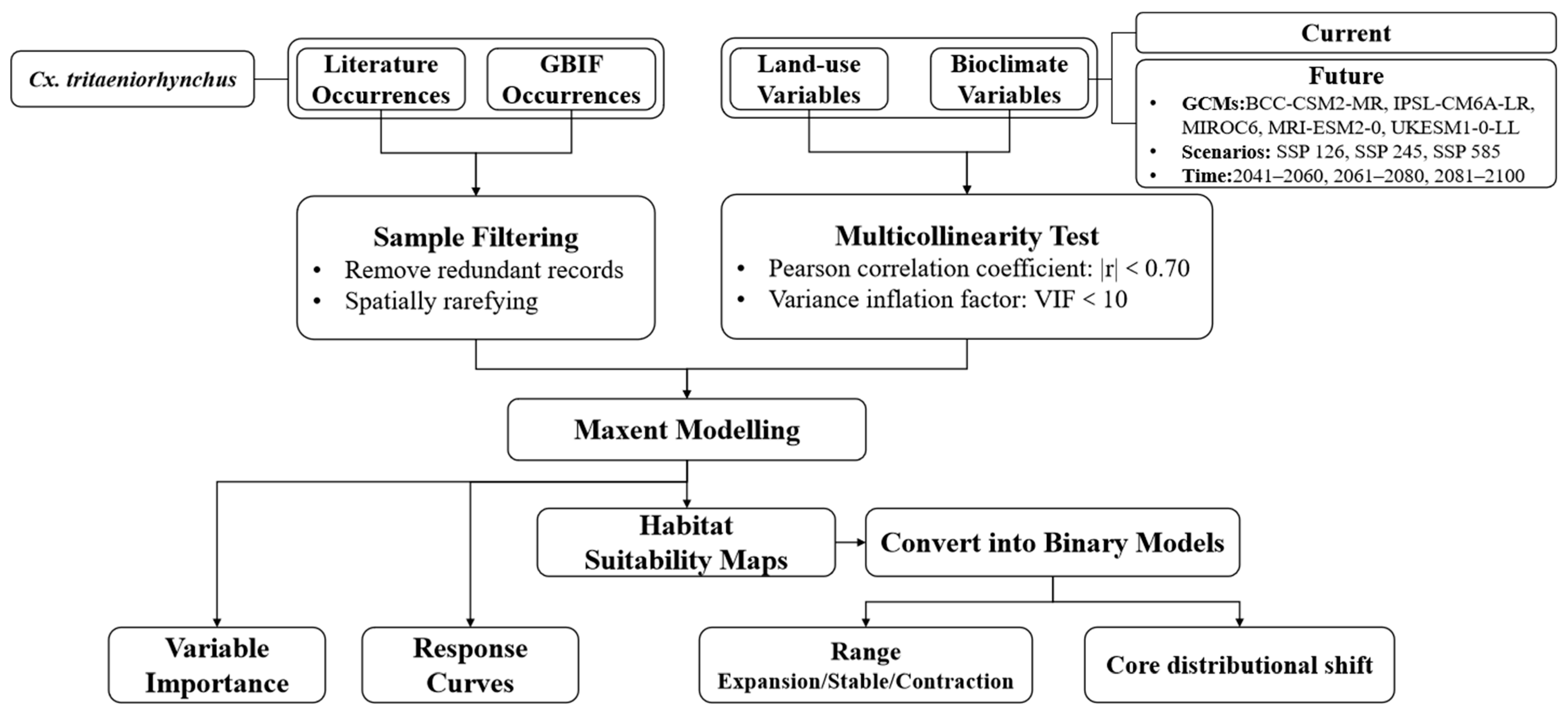
2. Materials and Methods
2.1. Cx. tritaeniorhynchus Occurrence Data
2.2. Environmental Variables
2.3. Maxent Modeling
2.4. Suitable Habitat Shifts
3. Results
3.1. Current Distribution of Cx. tritaeniorhynchus
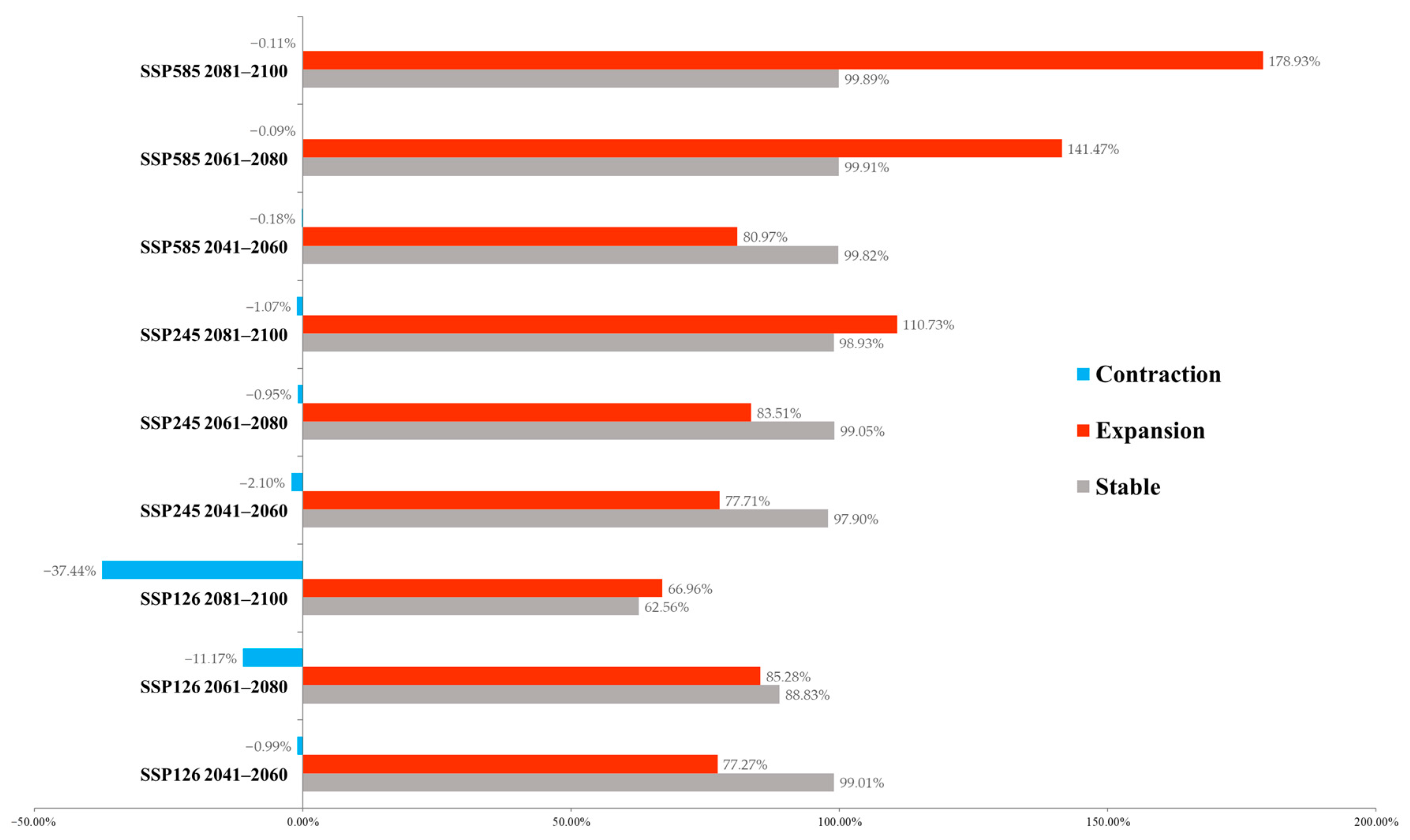
3.2. Future Habitat Changes Based on Climate Change Scenarios
3.3. Core Distributional Shifts
3.4. Variable Importance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ENM | Ecological niche model |
| VBD | Vector-borne disease |
| SSP | Shared socioeconomic pathway |
| GCM | Global climate model |
References
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D. Estimated global incidence of japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [PubMed]
- Liang, G.; Li, X.; Gao, X.; Fu, S.; Wang, H.; Li, M.; Lu, Z.; Zhu, W.; Lu, X.; Wang, L. Arboviruses and their related infections in china: A comprehensive field and laboratory investigation over the last 3 decades. Rev. Med. Virol. 2018, 28, e1959. [Google Scholar]
- Chen, X.J.; Wang, H.Y.; Li, X.L.; Gao, X.Y.; Li, M.H.; Fu, S.H.; He, Y.; Li, F.; Yin, Q.K.; Xu, S.T.; et al. Japanese encephalitis in china in the period of 1950–2018: From discovery to control. Biomed. Environ. Sci. 2021, 34, 175–183. [Google Scholar]
- Yan, W.-Z.; Li, J.-Y.; Guo, Y.-H.; Yue, Y.-J.; Liu, X.-B.; Song, X.-P.; Lu, L.; Zhao, N. Research progress on the epidemic characteristics and virus genotypes of japanese encephalitis in China. Chin. J. Vector Biol. Control 2024, 35, 744–752. [Google Scholar]
- Sanisuriwong, J.; Yurayart, N.; Thontiravong, A.; Tiawsirisup, S. Vector competence of culex tritaeniorhynchus and culex quinquefasciatus (diptera: Culicidae) for duck tembusu virus transmission. Acta Trop. 2021, 214, 105785. [Google Scholar] [PubMed]
- Eifan, S.; Hanif, A.; Nour, I.; Alqahtani, S.; Eisa, Z.M.; Dafalla, O.; Kohl, A. Distribution and molecular identification of culex pipiens and culex tritaeniorhynchus as potential vectors of rift valley fever virus in jazan, saudi arabia. Pathogens 2021, 10, 1334. [Google Scholar] [CrossRef]
- Wang, J.; Xu, H.; Song, S.; Cheng, R.; Fan, N.; Fu, S.; Zhang, S.; Xu, Z.; He, Y.; Lei, W. Emergence of zika virus in culex tritaeniorhynchus and anopheles sinensis mosquitoes in china. Virol. Sin. 2021, 36, 33–42. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, Z.; Li, K.; Rehman, M.U.; Nawaz, S.; Kulyar, M.F.-e.-A.; Hu, M.; Zhang, W.; Zhang, Z.; An, M. Detection of culex tritaeniorhynchus giles and novel recombinant strain of lumpy skin disease virus causes high mortality in yaks. Viruses 2023, 15, 880. [Google Scholar] [CrossRef]
- Kulhanek, S.A.; Leung, B.; Ricciardi, A. Using ecological niche models to predict the abundance and impact of invasive species: Application to the common carp. Ecol. Appl. 2011, 21, 203–213. [Google Scholar]
- Gaikwad, J.; Wilson, P.D.; Ranganathan, S. Ecological niche modeling of customary medicinal plant species used by australian aborigines to identify species-rich and culturally valuable areas for conservation. Ecol. Model. 2011, 222, 3437–3443. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Ma, J.; Jiao, Z.; Xiao, J.; Hayat, M.A.; Wang, H. Modeling the present and future distribution of arbovirus vectors aedes aegypti and aedes albopictus under climate change scenarios in mainland China. Sci. Total Environ. 2019, 664, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, W.; Guan, Y.; Qi, J.; Roberts, N.J.; Wen, D.; Cheng, Z.; Shan, F.; Zhao, Y.; Gu, J. The fine-scale movement pattern of amur tiger (panthera tigris altaica) responds to winter habitat permeability. Wildl. Lett. 2023, 1, 119–130. [Google Scholar] [CrossRef]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A.M. Want to model a species niche? A step-by-step guideline on correlative ecological niche modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Liu, B.; Ma, J.; Jiao, Z.; Gao, X.; Xiao, J.; Wang, H. Risk assessment for the rift valley fever occurrence in china: Special concern in south-west border areas. Transbound. Emerg. Dis. 2021, 68, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Brownstein, J.S.; Holford, T.R.; Fish, D. A climate-based model predicts the spatial distribution of the lyme disease vector ixodes scapularis in the united states. Environ. Health Perspect. 2003, 111, 1152–1157. [Google Scholar] [CrossRef]
- Gao, X.; Xiao, J.; Liu, B.; Wang, H. Impact of meteorological and geographical factors on the distribution of phlebotomus chinensis in northwestern mainland china. Med. Vet. Entomol. 2018, 32, 365–371. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Ma, J.; Jiao, Z.; Xiao, J.; Wang, H. Influence of host and environmental factors on the distribution of the japanese encephalitis vector culex tritaeniorhynchus in china. Int. J. Environ. Res. Public Health 2018, 15, 1848. [Google Scholar] [CrossRef]
- Franklinos, L.H.; Jones, K.E.; Redding, D.W.; Abubakar, I. The effect of global change on mosquito-borne disease. Lancet Infect. Dis. 2019, 19, e302–e312. [Google Scholar] [CrossRef]
- Brown, J.L. Sdm toolbox: A python-based gis toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Wu, J.; Shi, Y.; Xu, Y. Evaluation and projection of surface wind speed over china based on cmip6 gcms. J. Geophys. Res. Atmos. 2020, 125, e2020JD033611. [Google Scholar]
- Yue, Y.; Yan, D.; Yue, Q.; Ji, G.; Wang, Z. Future changes in precipitation and temperature over the yangtze river basin in china based on cmip6 gcms. Atmos. Res. 2021, 264, 105828. [Google Scholar]
- Guo, J.; Wang, X.; Xiao, C.; Liu, L.; Wang, T.; Shen, C. Evaluation of the temperature downscaling performance of precis to the bcc-csm2-mr model over china. Clim. Dyn. 2022, 59, 1143–1159. [Google Scholar]
- Shiru, M.S.; Chung, E.-S.; Shahid, S.; Wang, X.-J. Comparison of precipitation projections of cmip5 and cmip6 global climate models over yulin, china. Theor. Appl. Climatol. 2022, 147, 535–548. [Google Scholar]
- Tian, J.; Zhang, Z.; Ahmed, Z.; Zhang, L.; Su, B.; Tao, H.; Jiang, T. Projections of precipitation over china based on cmip6 models. Stoch. Environ. Res. Risk Assess. 2021, 35, 831–848. [Google Scholar]
- Xiang, Y.; Wang, Y.; Chen, Y.; Zhang, Q. Impact of climate change on the hydrological regime of the yarkant river basin, china: An assessment using three ssp scenarios of cmip6 gcms. Remote Sens. 2022, 14, 115. [Google Scholar]
- Zeng, Y.; Low, B.W.; Yeo, D.C. Novel methods to select environmental variables in maxent: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar]
- Duque-Lazo, J.; Van Gils, H.; Groen, T.; Navarro-Cerrillo, R. Transferability of species distribution models: The case of phytophthora cinnamomi in southwest spain and southwest australia. Ecol. Model. 2016, 320, 62–70. [Google Scholar]
- Kumar, S.; Graham, J.; West, A.M.; Evangelista, P.H. Using district-level occurrences in maxent for predicting the invasion potential of an exotic insect pest in india. Comput. Electron. Agric. 2014, 103, 55–62. [Google Scholar]
- Linnell, M.A.; Davis, R.J.; Lesmeister, D.B.; Swingle, J.K. Conservation and relative habitat suitability for an arboreal mammal associated with old forest. For. Ecol. Manag. 2017, 402, 1–11. [Google Scholar]
- Pradhan, P. Strengthening maxent modelling through screening of redundant explanatory bioclimatic variables with variance inflation factor analysis. Researcher 2016, 8, 29–34. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Zheng, K.; Ma, J.; Jiao, Z.; Xiao, J.; Wang, H. The potential distribution and dynamics of important vectors culex pipiens pallens and culex pipiens quinquefasciatus in china under climate change scenarios: An ecological niche modelling approach. Pest Manag. Sci. 2020, 76, 3096–3107. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, Y.; Liu, Z.; Huo, W.; Cao, J.; Zhao, G.; Zhang, F.-G. Prediction of potential invasion of two weeds of the genus avena in asia under climate change based on maxent. Sci. Total Environ. 2024, 950, 175192. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M. The importance of correcting for sampling bias in maxent species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Cao, Y.; Dewalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using maxent to model the historic distributions of stonefly species in illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar]
- Mwakapeje, E.R.; Ndimuligo, S.A.; Mosomtai, G.; Ayebare, S.; Nyakarahuka, L.; Nonga, H.E.; Mdegela, R.H.; Skjerve, E. Ecological niche modeling as a tool for prediction of the potential geographic distribution of bacillus anthracis spores in tanzania. Int. J. Infect. Dis. 2019, 79, 142–151. [Google Scholar] [CrossRef]
- Wang, J.; Cheng, J.; Zhang, C.; Feng, Y.; Jin, L.; Wei, S.; Yang, H.; Cao, Z.; Peng, J.; Luo, Y. Prediction of the future evolution trends of prunus sibirica in china based on the key climate factors using maxent modeling. Biology 2024, 13, 973. [Google Scholar] [CrossRef]
- Tang, W.; Ye, S.; Qin, S.; Fan, Q.; Tang, J.; Zhang, H.; Liu, J.; Huang, Z.; Liu, W. Habitat for coilia nasus in southern zhejiang province, china, based on a maximum entropy model. Sci. Rep. 2024, 14, 19254. [Google Scholar]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [PubMed]
- Cunze, S.; Koch, L.K.; Kochmann, J.; Klimpel, S. Aedes albopictus and aedes japonicus-two invasive mosquito species with different temperature niches in Europe. Parasites Vectors 2016, 9, 573. [Google Scholar]
- Li, S.; Wang, Z.; Zhu, Z.; Tao, Y.; Xiang, J. Predicting the potential suitable distribution area of emeia pseudosauteri in zhejiang province based on the maxent model. Sci. Rep. 2023, 13, 1806. [Google Scholar]
- Yuan, Y.; Tang, X.; Liu, M.; Liu, X.; Tao, J. Species distribution models of the spartina alterniflora loisel in its origin and invasive country reveal an ecological niche shift. Front. Plant Sci. 2021, 12, 738769. [Google Scholar]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- World Health Organization. Ethics and Vector-Borne Diseases: WHO Guidance. 2020. Available online: https://iris.who.int/bitstream/handle/10665/336075/9789240012738-eng.pdf (accessed on 3 February 2025).
- Rocklöv, J.; Dubrow, R. Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Yi, H.; Devkota, B.R.; Yu, J.S.; Oh, K.C.; Kim, J.; Kim, H.J. Effects of global warming on mosquitoes & mosquito-borne diseases and the new strategies for mosquito control. Entomol. Res. 2014, 44, 215–235. [Google Scholar]
- Shapiro, L.L.; Whitehead, S.A.; Thomas, M.B. Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLoS Biol. 2017, 15, e2003489. [Google Scholar]
- Liu-Helmersson, J.; Stenlund, H.; Wilder-Smith, A.; Rocklöv, J. Vectorial capacity of aedes aegypti: Effects of temperature and implications for global dengue epidemic potential. PLoS ONE 2014, 9, e89783. [Google Scholar]
- Yan, T.; Liu, M.-D.; Liu, T.; Fu, X.-F.; Zhou, X.-J.; Tong, Y. Investigation on main vectors in communities of beijing city in 2018 and 2019. Chin. J. Hyg. Insectic. Equip. 2021, 27, 412–415. [Google Scholar]
- Zhang Hui, Z.H.; Luo HouQiang, L.H.; Mujeeb-ur-Rehman, M.-u.-R.; Fazul Nabi, F.N.; Li Kun, L.K.; Lan YanFang, L.Y.; Huang ShuCheng, H.S.; Zhang LiHong, Z.L.; Khalid Mehmood, K.M.; Muhammad Shahzad, M.S. Evidence of jev in culex tritaeniorhynchus and pigs from high altitude regions of tibet, china. J. Vector Borne Dis. 2017, 54, 69. [Google Scholar]
- Takagi, M.; Maruyama, K.; Sugiyama, A. A laboratory experiment on the larval development of culex tritaeniorhynchus (diptera: Culicidae) giles under different temperatures and densities. Trop. Med. 1996, 38, 69–77. [Google Scholar]
- Reisen, W.K.; Siddiqui, T.F.; Aslam, Y.; Malik, G.M. Geographic variation among the life table characteristics of culex tritaeniorhynchus from asia. Ann. Entomol. Soc. Am. 1979, 72, 700–709. [Google Scholar]
- Foster, W.A. Mosquito sugar feeding and reproductive energetics. Annu. Rev. Entomol. 1995, 40, 443–474. [Google Scholar]
- Lytra, I.; Emmanouel, N. Study of culex tritaeniorhynchus and species composition of mosquitoes in a rice field in greece. Acta Trop. 2014, 134, 66–71. [Google Scholar]
- Lindahl, J.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Occurrence of japanese encephalitis virus mosquito vectors in relation to urban pig holdings. Am. J. Trop. Med. Hyg. 2012, 87, 1076. [Google Scholar]
- Pietrosemoli, S.; Tang, C. Animal welfare and production challenges associated with pasture pig systems: A review. Agriculture 2020, 10, 223. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, M.; Wang, H.; Liang, G. Japanese encephalitis and japanese encephalitis virus in mainland china. Rev. Med. Virol. 2012, 22, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Nasci, R.; Liang, G. The neglected arboviral infections in mainland China. PLoS Neglected Trop. Dis. 2010, 4, e624. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Y.; Liang, X.; Liang, G. Japanese encephalitis in mainland China. Jpn. J. Infect. Dis. 2009, 62, 331–336. [Google Scholar] [PubMed]
- Miller, R.H.; Masuoka, P.; Klein, T.A.; Kim, H.-C.; Somer, T.; Grieco, J. Ecological niche modeling to estimate the distribution of japanese encephalitis virus in asia. PLoS Neglected Trop. Dis. 2012, 6, e1678. [Google Scholar] [CrossRef]
- Wiese, D.; Escalante, A.A.; Murphy, H.; Henry, K.A.; Gutierrez-Velez, V.H. Integrating environmental and neighborhood factors in maxent modeling to predict species distributions: A case study of aedes albopictus in southeastern pennsylvania. PLoS ONE 2019, 14, e0223821. [Google Scholar] [CrossRef]
- Barker, J.R.; MacIsaac, H.J. Species distribution models applied to mosquitoes: Use, quality assessment, and recommendations for best practice. Ecol. Model. 2022, 472, 110073. [Google Scholar] [CrossRef]
- Conley, A.K.; Fuller, D.O.; Haddad, N.; Hassan, A.N.; Gad, A.M.; Beier, J.C. Modeling the distribution of the west nile and rift valley fever vector culex pipiens in arid and semi-arid regions of the middle east and North Africa. Parasites Vectors 2014, 7, 289. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidate Variables | Description | Source | Resolution | Included in the Final Model |
|---|---|---|---|---|
| Bio 1 | Annual mean temperature | WorldClim | 2.5 arcmin | Yes |
| Bio 2 | Mean diurnal range | Yes | ||
| Bio 3 | Isothermality (Bio 2/Bio 7) | |||
| Bio 4 | Temperature seasonality (Standard deviation*100) | |||
| Bio 5 | Maximum temperature of the warmest month | |||
| Bio 6 | Minimum temperature of the coldest month | |||
| Bio 7 | Temperature annual range (Bio 5-Bio 6) | Yes | ||
| Bio 8 | Mean temperature of the wettest quarter | |||
| Bio 9 | Mean temperature of the driest quarter | |||
| Bio 10 | Mean temperature of the warmest quarter | |||
| Bio 11 | Mean temperature of the coldest quarter | |||
| Bio 12 | Annual precipitation | Yes | ||
| Bio 13 | Precipitation of the wettest month | |||
| Bio 14 | Precipitation of the driest month | |||
| Bio 15 | Precipitation seasonality (Coefficient of variation) | Yes | ||
| Bio 16 | Precipitation of the wettest quarter | |||
| Bio 17 | Precipitation of the driest quarter | |||
| Bio 18 | Precipitation of the warmest quarter | |||
| Bio 19 | Precipitation of the coldest quarter | |||
| C3ann | C3 annual crops | LUH2 | 0.25 Degree | Yes |
| C3per | C3 perennial crops | Yes | ||
| C3nfx | C3 nitrogen-fixing crops | Yes | ||
| C4ann | C4 annual crops | Yes | ||
| C4per | C4 perennial crops | Yes | ||
| Primf | Forested primary land | Yes | ||
| Primn | Non-forested primary land | |||
| Secdf | Potentially forested secondary land | |||
| Secdn | Potentially non-forested secondary land | Yes | ||
| Pastr | Managed pasture | Yes | ||
| Range | Rangeland | Yes | ||
| Urban | Urban land | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Li, L.; Zhang, Z.; Ran, H.; Xing, M. Modeling the Potential Distribution and Future Dynamics of Important Vector Culex tritaeniorhynchus Under Climate Change Scenarios in China. Insects 2025, 16, 382. https://doi.org/10.3390/insects16040382
Liu B, Li L, Zhang Z, Ran H, Xing M. Modeling the Potential Distribution and Future Dynamics of Important Vector Culex tritaeniorhynchus Under Climate Change Scenarios in China. Insects. 2025; 16(4):382. https://doi.org/10.3390/insects16040382
Chicago/Turabian StyleLiu, Boyang, Li Li, Zhulin Zhang, Haoyu Ran, and Mingwei Xing. 2025. "Modeling the Potential Distribution and Future Dynamics of Important Vector Culex tritaeniorhynchus Under Climate Change Scenarios in China" Insects 16, no. 4: 382. https://doi.org/10.3390/insects16040382
APA StyleLiu, B., Li, L., Zhang, Z., Ran, H., & Xing, M. (2025). Modeling the Potential Distribution and Future Dynamics of Important Vector Culex tritaeniorhynchus Under Climate Change Scenarios in China. Insects, 16(4), 382. https://doi.org/10.3390/insects16040382