Simple Summary
Varroa destructor is a tiny parasitic mite that threatens honey bee populations worldwide by spreading deadly viruses. While efforts to control these mites often rely on chemical treatments, these can harm beneficial insects and lead to resistant mite populations. A promising alternative is using RNA-based treatments to disrupt essential mite functions, such as immunity. However, scientists still know little about how Varroa defends itself against infections. In this study, we analyzed the genetic makeup of Varroa’s immune system and compared it to that of related species, such as ticks. Our results show that, similar to ticks, Varroa lacks certain immune system components found in other arthropods. Its repertoire of antimicrobial peptides is more closely related to that of ticks and spiders than to that of insects. These findings help us better understand how the mite fights infections and provide a foundation for developing new, targeted methods to weaken and control Varroa populations. By improving our knowledge of mite immunity, this research contributes to safer and more effective ways to protect honey bee colonies, ensuring their survival and the vital pollination services they provide to ecosystems and agriculture.
Abstract
Despite its ecological and economic importance, many aspects of Varroa destructor’s biology remain poorly understood, particularly its defense mechanisms against pathogens. The limited knowledge of Varroa’s immunity has hindered the development of RNA interference (RNAi)-based strategies targeting immune-related genes. In this study, we investigated the immune gene repertoire of V. destructor by querying its NCBI nr protein database and comparing it to model species of ticks (Ixodes scapularis) and mites (Galendromus occidentalis and Tetranychus urticae). Transcription of candidate immune genes was confirmed by analyzing a de novo assembled transcriptome of V. destructor. Our findings reveal that V. destructor shares key immunological traits with ticks, including lysozymes, chitinases, and thioester-containing proteins (TEPs), but also shares the absence of transmembrane peptidoglycan recognition proteins (PGRPs), Gram-negative binding proteins, and several lectin families involved in pathogen recognition. Additionally, Varroa mites, like ticks, lack homologs of crucial immune signaling components, such as the unpaired ligand (JAK/STAT), Eiger (JNK), and multiple elements of the IMD pathway. They also do not encode canonical antimicrobial peptides (AMPs) like defensins but possess putative homologs of ctenidins, AMPs previously identified in spiders and ticks, which may be adopted as a novel genetic readout for immune response in mites. Our findings lay the groundwork for future functional studies on mite immunity and open new avenues for RNAi-based biocontrol strategies targeting immune pathways to enhance Varroa management.
1. Introduction
Honey bees (Apis mellifera) are facing a serious trend of increasing colony losses caused by a multifactorial syndrome in which parasites and pathogens play a major role [1,2]. As indicated by most surveys, colony losses are associated with high levels of Varroa destructor and pathogenic viruses, which are vectored by the mites [3,4]. This arthropod is an obligate ectoparasite of honey bees, and has a negative impact on the host, both at the colony and individual level [5,6], since it feeds on liquid tissues through a wound made on the integument of the host [7,8]. This trophic activity has important consequences for the honey bee host, such as the reduction in longevity and weight at emergence [9,10], and the transmission of viral pathogens [11]. In addition, Varroa can also impact reproductive fitness, homing, orientation abilities, and also the immune response of bees [12,13].
With a nearly global presence [11,14], V. destructor can significantly weaken or even cause the collapse of honey bee colonies if left untreated [15]. Currently, most beekeepers rely heavily on chemical controls to manage the mite [16], despite their detrimental effects on non-target organisms, their residual presence in hive products, and the insurgence of acaricide resistance [17]. This highlights the importance of adopting an Integrated Pest Management (IPM) approach for Varroa destructor, which should combine different control methods, including genetic, mechanical, biological, and biotechnical tools, to reduce the impact of treatments with synthetic pesticides [18].
Additional tools for Varroa control include “soft” acaricides, such as oxalic acid and thymol, as well as biopesticides like dsRNA [19]. dsRNA works by inducing the degradation of RNA sequences with similar structures, resulting in targeted gene silencing. This process interferes with the synthesis of the encoded protein, enabling selective targeting of specific organismal functions [20].
dsRNA-based control strategies can be classified as either direct killing, targeting vital genes essential for survival, or indirect killing, where lethality occurs only when combined with another treatment. For instance, targeting immune genes with dsRNA combined with a sublethal dose of an entomopathogen-based biopesticide has been proposed as an indirect killing strategy [21].
RNA interference (RNAi) via dsRNA treatments is effective against a wide range of mite species. Since the first application of RNAi through injection in the spider mite Tetranychus urticae, silencing the Distal-less gene [22], an increasing number of herbivore and also predator mite species have been shown to be susceptible to RNAi such as Dermatophagoides pteronyssinus, Metaseiulus occidentalis, Panonychus citri, Phytoseiulus persimilis, Sarcoptes scabiei, Tetranychus cinnabarinus, Tetranychus urticae, and Varroa destructor. The genome sequencing of T. urticae allowed research on mites to enter the genome era. A study showed that soaking of mites in the dsRNA solution and mites feeding on dsRNA-coated leaves were the two most efficient methods, and this delivery method may act as a mimic for sprayable dsRNA in future field applications [23].
dsRNA proved to be both (1) deliverable to V. destructor by adding it to the artificial diet of adult bees [24,25] or through engineering bees’ symbiontic bacteria [26] and (2) effective in controlling mites by targeting several genes. Promising results were observed in the simultaneous silencing of 14 vital genes, including those involved in the cytoskeleton, energy metabolism, and RNA polymerase, significantly reducing mite populations [24,26]. Additionally, salivary genes, which are likely critical for mite feeding, were identified as vital; their silencing corresponded with reduced mite survival [27,28]. More recently, targeting the calmodulin gene to disrupt Varroa reproduction led to a significant decrease in mite offspring, though it did not affect mite survival [25]. Despite these advances, the impact of RNAi on many biological functions of V. destructor remains unexplored, limiting the potential for indirect strategies such as those that interfere with the mite’s immune response.
Arthropods have a more elementary cell-mediated defense system than vertebrates because of the lack of an antigen–antibody complex and memory cells [29]. However, arthropods have evolved effective innate immunity that acts through both cellular and humoral responses directed against invaders [30]. Activation of these responses occurs through the recognition of pathogen-associated molecular patterns (PAMPs) by receptors, known as pattern recognition receptors (PRRs), located on hemocytes (immune cells) and epithelial cells from barrier sites throughout the body [31,32]. Cellular immune responses include phagocytosis, nodulation, encapsulation, and melanization events mediated by hemocytes [33,34,35]. Humoral responses orchestrated by signaling pathways such as IMD, Toll, JAK/STAT, and JNK lead to the synthesis of various defense enzymes, complement-like proteins, and antimicrobial peptides (AMPs) in response to infection [36,37,38]. Most of the knowledge about the immunity of the Acarina group (mites and ticks) comes from research on the immune responses of ticks, which are structured differently from those of insects and crustaceans. [39]. In ticks and other chelicerates, core intracellular components of the “canonical” IMD pathway, such as the NF-κB transcription factor Relish, are present. However, genome and functional studies indicate that ticks lack key upstream elements, including transmembrane PGRPs and the adapter molecules IMD and FADD [30,40,41,42,43].
In this study, we investigated the immune gene repertoire of V. destructor by querying its NCBI nr protein database. To place these genes in an evolutionary context, we conducted additional analyses, including comparisons with model species of ticks (Ixodes scapularis) and mites (Galendromus occidentalis and Tetranychus urticae).
2. Materials and Methods
We used a broadly used approach to identify immunity-related genes by inferring protein homology based on sequence similarity through BLASTp searches [44,45]. Immune gene candidates from D. melanogaster, reported in Table 1, Table 2 and Table 3, were used to query the NCBI nr protein database by limiting the target organism to V. destructor, Ixodes scapularis, Galendromus occidentalis, and Tetranychus urticae. Only hits with an E-Value less than 1 × 10−5 were considered positive.
To validate the identified candidate immune genes, we used their protein sequences to query a de novo assembled transcriptome of V. destructor. The transcriptome was assembled using Illumina reads obtained from a previous study (SRA BioProject ID: PRJNA1097857), in which we analyzed the salivary gland transcriptome of V. destructor by comparing it with the transcriptome of the rest of the adult female body [28]. First, reads of all the samples, obtained from more than 140 mites, were cleaned using Trimmomatic [46], merged, and de novo assembled with Trinity [47]. Then, the candidate immune genes were used as the query in tBLASTn searches against the assembled transcriptome. We considered positivity at the transcription level only if the query sequenced had more than 80% identity with the best hit in the transcriptome.
To reconstruct the immune gene phylogeny, putative homologous sequences in other insect species (Apis mellifera, Tribolium castaneum, and Bombyx mori) were identified by sequence similarity searches through BLASTp, using D. melanogaster proteins (Table 1, Table 2 and Table 3) as the query versus the non-redundant NCBI database (nr NCBI db). One best hit per query was selected and all the protein sequences were aligned using Muscle 3.8 [48], with default settings.
Alignments were automatically trimmed using Gblocks version 0.91b [49] to avoid comparisons of non-conserved regions present only in a subset of the taxa. The best-fit model of amino acid substitution and phylogenetic reconstruction was generated using RAxML 8.2.12 [50]. The maximum likelihood tree was run for 1000 bootstrap replicates and the tree figure was plotted using FigTree v1.4.3. Protein sequences were analyzed with ScanProsite (https://prosite.expasy.org/scanprosite/, accessed on 5 November 2024) and InterProScan [51], in order to identify active sites, conserved patterns, and domains [52]. For PGRPs, we scanned putative homologs for transmembrane helices using DeepTMHMM, which predicts alpha and beta transmembrane proteins using deep neural networks [53].
3. Results and Discussion
3.1. Overview of the Immune Gene Survey
We selected a subset of genes involved in the three phases of the arthropod immune response: recognition, signaling, and response. By using protein sequences from D. melanogaster, known to be involved in immunity, as queries in BLASTp searches, we successfully identified putative homologs of Drosophila canonical immune genes in V. destructor. Specifically, we identified proteins related to recognition, signaling, and response to pathogenic microorganisms (Figure 1).
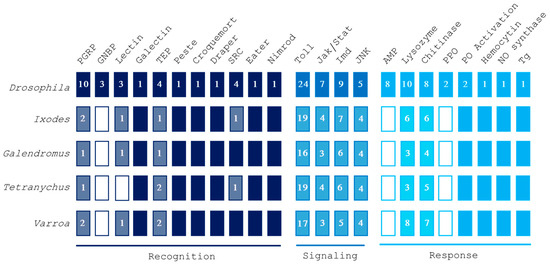
Figure 1.
Overview of gene families involved in recognition, signaling, and response pathways in Varroa destructor, Tetranychus urticae, Galendromus occidentalis, and Ixodes scapularis. Putative homology was inferred by using Drosophila melanogaster queries. Numbers in blocks indicate the amount of different genes identified for each protein family. Filled boxes imply the presence of all the analyzed genes, while empty boxes represent a total absence of them. Abbreviations: peptidoglycan recognition protein (PGRP), Gram-negative binding protein (GNBP), thioester-containing protein (TEP), scavenger receptor class C (SRC), antimicrobial peptide (AMP), pro-phenoloxidase (PPO), phenoloxidase (PO), nitric oxide (NO), transglutaminase (Tg).
3.2. Recognition Genes
Our analysis pointed out the occurrence of proteins encoded by the V. destructor genome with significant matches with Drosophila genes involved in recognition (Table 1). Among these, only one candidate recognition gene was not transcribed in adult female mites (Supplementary File S1). As occurs in ticks and predatory/phytophagous mites (Tables S1–S3), V. destructor lacks transmembrane peptidoglycan recognition proteins (PGRPs), Gram-negative binding proteins, and some members of the c-type lectin family.

Table 1.
Immune-related proteins of Varroa destructor involved in recognition. Orthologs not found in V. destructor are in italics.
Table 1.
Immune-related proteins of Varroa destructor involved in recognition. Orthologs not found in V. destructor are in italics.
| Gene Name | Role | D. melanogaster | V. destructor 1 | E-Value | Identity | Coverage | Transcription |
|---|---|---|---|---|---|---|---|
| PGRP-LC, peptidoglycan recognition protein | bacterial recognition | AAF50302.3 | XP_022660134.1 | 2 × 10−30 | 36.71% | 31% | YES |
| PGRP-LE, peptidoglycan recognition protein | activation of PPO cascade and autophagy | NP_573078.1 | XP_022660134.1 | 5 × 10−25 | 31.01% | 45% | YES |
| PGRP-SA, peptidoglycan recognition protein | bacterial recognition | AAF48056.1 | XP_022660135.1 | 9 × 10−34 | 32.93% | 82% | YES |
| PGRP-SD, peptidoglycan recognition protein | bacterial recognition | CAD89193.1 | XP_022660134.1 | 5 × 10−27 | 33.54% | 86% | YES |
| PGRP-LB, peptidoglycan recognition protein | bacterial recognition | NP_650079.1 | XP_022660134.1 | 2 × 10−31 | 35.09% | 79% | YES |
| PGRP-SC1a, peptidoglycan recognition protein | bacterial recognition | CAD89161.1 | XP_022660134.1 | 3 × 10−32 | 30.62% | 86% | YES |
| PGRP-SC2, peptidoglycan recognition protein | bacterial recognition | CAD89187.1 | XP_022660134.1 | 3 × 10−35 | 31.71% | 89% | YES |
| PGRP-SB1, peptidoglycan recognition protein | PGN degradation and antibacterial activity | CAD89136.1 | XP_022660135.1 | 6 × 10−35 | 35.22% | 83% | YES |
| PGRP-LF, peptidoglycan recognition protein | blocking of IMD pathway | NP_648299.3 | XP_022660134.1 | 3 × 10−38 | 37.35% | 79% | YES |
| PGRP-LA, peptidoglycan recognition protein | activation of IMD pathway | AAF50304.2 | Not found | Not found | Not found | Not found | - |
| GNBP1, Gram-negative binding protein 1 | bacterial and fungal pattern recognition | Q9NHB0.2 | Not found | Not found | Not found | Not found | - |
| GNBP2, Gram-negative binding protein 2 | bacterial and fungal pattern recognition | ACU30172.1 | Not found | Not found | Not found | Not found | - |
| GNBP3, Gram-negative binding protein 3 | bacterial and fungal pattern recognition | CAJ18910.1 | Not found | Not found | Not found | Not found | - |
| DL1, c-type lectin 1 | bacterial recognition, induction of PPO cascade | AAF53793.1 | Not found | Not found | Not found | Not found | - |
| DL2, c-type lectin 2 | bacterial recognition, induction of PPO cascade | NP_001014489.1 | Not found | Not found | Not found | Not found | - |
| DL3, c-type lectin 3 or solute carrier | bacterial recognition, induction of PPO cascade | NP_001014490.1 | XP_022646715.1 | 4 × 10−7 | 23.58% | 79% | YES |
| galectin 4 | several roles have been hypothesized | ADZ99399.1 | XP_022654763.1 | 1 × 10−9 | 34.75% | 29% | YES |
| TEP1, CD109 antigen-like | mark pathogens for phagocytosis | CAB87807.1 | XP_022645189.1 | 0.0 | 31.32% | 98% | YES |
| TEP2, CD109 antigen-like | mark pathogens | CAB87808.1 | XP_022645189.1 | 0.0 | 33.79% | 98% | YES |
| TEP3, CD109 antigen-like | mark pathogens | AAL39195.1 | XP_022645188.1 | 0.0 | 32.35% | 97% | YES |
| TEP4, CD109 antigen-like | mark pathogens | NP_523603.2 | XP_022645188.1 | 0.0 | 30.90% | 98% | YES |
| Pes, scavenger receptor class B member 1-like | bacterial and fungal recognition | AHN54246.1 | XP_022673026.1 | 4 × 10−61 | 29.15% | 78% | YES |
| Crq, croquemort, lysosome membrane protein 2-like | bacterial and fungal recognition | AAF51494.1 | XP_022672747.1 | 5 × 10−70 | 28.81% | 96% | YES |
| Drpr, protein draper-like | bacterial and fungal recognition | NP_477450.1 | XP_022656525.1 | 5 × 10−69 | 39.01% | 72% | YES |
| sr-CI, scavenger receptor class C, type i | bind to lipoproteins and bacteria | AAW79470.1 | XP_022658153.1 | 3 × 10−24 | 28.62% | 45% | YES |
| sr-CII, scavenger receptor class C, type ii | bind to lipoproteins and bacteria | AAF58551.1 | XP_022658154.1 | 4 × 10−23 | 27.16% | 49% | YES |
| sr-CIII, scavenger receptor class C, type iii | bind to lipoproteins and bacteria | AAF37564.1 | XP_022658154.1 | 2 × 10−9 | 21.00% | 90% | YES |
| sr-CIV, scavenger receptor class C, type iv | bind to lipoproteins and bacteria | AAF51092.1 | XP_022658154.1 | 5 × 10−19 | 26.59% | 73% | YES |
| eater | receptor in phagocytosis and microbial binding | AAF56664.5 | XP_022668925.1 | 2 × 10−17 | 33.72% | 77% | NO |
| Drp, protein draper-like | receptor in phagocytosis and microbial binding | AAF53364.2 | XP_022656529.1 | 6 × 10−12 | 27.66% | 62% | YES |
1 Best hit on the E-Value basis obtained by BLASTp searches against the NCBI nr database.
3.2.1. Peptidoglycan Receptor Proteins
Nearly all bacteria have peptidoglycans as crucial components of the cell wall, which are detected by the immune system thanks to pathogen recognition receptors (PRRs). Various families of pattern recognition molecules that identify peptidoglycans have been discovered in insects, and the function of peptidoglycan recognition proteins (PGRPs) in immune defense is relatively well understood in Drosophila [54]. Recognition through PGRPs triggers both the Toll and IMD/JNK signaling pathways, resulting in the activation of prophenoloxidase (proPO) or the production of antimicrobial peptides [55]. Most insect species have multiple PGRP genes that vary in both structure and function. For instance, Drosophila contains 13 PGRP genes that produce 19 different proteins, whereas Anopheles gambiae has 7 PGRP genes that encode 9 distinct proteins [54]. Conversely, our analysis revealed a limited set of PGRPs, identifying only a single putative homolog gene in the species we studied (Figure 2). This gene encodes two isoforms in V. destructor and I. scapularis, and a single protein in G. occidentalis and T. urticae. Furthermore, TMHMM analysis indicated that the PGRPs in V. destructor lack transmembrane domains, consistent with observations in other mites and ticks [30].
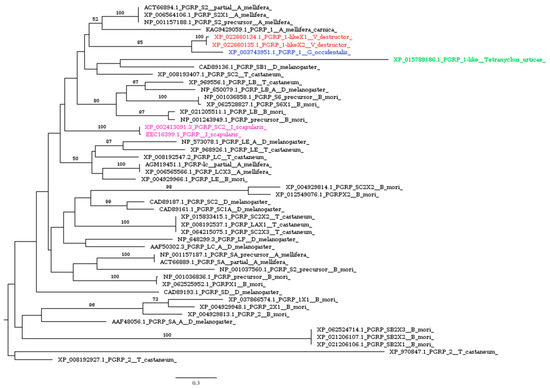
Figure 2.
Phylogenetic tree based on maximum likelihood analysis of PGRP putative homologs identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
This lack of transmembrane PGPRs in Acarina seems to be associated with the evolution of alternative protein sensors. Indeed, previous works demonstrated that the IMD and JNK pathways of the tick I. scapularis are activated by lipid molecular patterns [43], such as 1-palmitoyl-2-oleoyl-sn-glycero-phosphoglycerol (POPG), sensed by the homolog of croquemort (Crq) [39], a CD36-like protein originally identified in Drosophila [56]. Here, we report the presence of the Crq homolog in V. destructor (Table 1), which suggests a similar mechanism of recognition in mites.
3.2.2. Gram-Negative Binding Proteins
The structure of Gram-negative binding proteins (GNBPs) is characterized by a carbohydrate-binding module (CBM) at the N-terminus and a glucanase-like domain (Glu) at the C-terminus [57]. The CBM engages with microbial polysaccharides, whereas the Glu domain interacts with subsequent proteases, thus triggering immune signaling pathways [58]. GNBPs detect both bacterial and fungal pathogens, thus activating immune signaling cascades in insects [59]. In Drosophila, GNBP1 and peptidoglycan-recognition protein SA (PGRP-SA) work together to trigger the Toll pathway in response to Gram-positive bacterial infections [60], while GNBP3 plays a crucial role in activating the Toll pathway during fungal infections [61].
The comparative study of Palmer and Jiggins (2015) revealed that GNBPs, also known as βGRPs, have been lost from chelicerates, likely through a single-loss event [30]. V. destructor is no exception and, like other Acarina species (Tables S1–S3), shows no similarities to known GNBPs of D. melanogaster (Table 1).
3.2.3. Lectins
Lectins, a diverse class of carbohydrate-binding proteins, play a crucial role in the immune defense of numerous insect species. These proteins are recognized for their wide-ranging ability to bind pathogens and their participation in various immune functions, including opsonization, melanization, synthesis of antibacterial peptides, encapsulation, and the direct elimination of bacteria [62,63]. The Drosophila c-type lectins (CTLs) DL2 and DL3 agglutinate Gram-negative Escherichia coli in a calcium-dependent manner [64]. Our analysis pointed out a reduced CTL repertoire in Acarina. V. destructor lacks homologs of DL1 and DL2 but possesses a putative homolog of DL3 (Table 1). Notably, no CTLs were detected in the genomes of I. scapularis, G. occidentalis, and T. urticae when using Drosophila sequences as the query (Tables S1–S3). However, by using the DL3 putative homolog identified in V. destructor as the query, significant hits were obtained for both I. scapularis and G. occidentalis, though not for T. urticae (Table S4), where the best match was with a sushi, von Willebrand factor type A, EGF, and pentraxin domain-containing protein 1. This intriguing reduction in CTLs in T. urticae is in line with what was observed after immune challenge in this phytophagous mite by bacteria injection. Indeed, T. urticae has lost the capacity to mount an induced immune response against bacteria, in contrast to other mites and chelicerates but similarly to the phloem-feeding aphid Acyrthosiphon pisum [65]. The suggested evolutionary link between ecological conditions regarding exposure to bacteria and the architecture of the immune response deserves further studies. Although DL3 is predicted to be a secreted protein, it is detected on the surface of Drosophila hemocytes and enhances encapsulation [64]. Similarly, our in silico analysis suggests that the DL3 homolog identified in V. destructor is also predicted to be secreted, indicating a potential analogous role in mite immunity.
Galectins, a widely prevalent family of lectins [66], exhibiting thiol group-dependent activity and present in both intracellular and extracellular spaces [67], are upregulated in mosquitoes upon exposure to bacterial infections as well as malaria parasite invasion [68]. Galectins present in the gut of the hematophagous sand fly Phlebotomus papatasi facilitate a specific interaction between the insect’s midgut and Leishmania major, a critical event for parasite survival, and play a role in determining species-specific vector competence [69]. More broadly, insect galectins are believed to participate in pathogen recognition, agglutination, and phagocytosis [66,70]. Variability in galectin transcripts across different insect species has been highlighted by genome-wide studies: 5 transcripts were identified in Drosophila melanogaster, 8 in Anopheles gambiae, and 12 in Aedes aegypti [71], with 4 identified in several Lepidoptera species [72]. Acarina species (Tables S1–S3), including V. destructor, possess putative homologs of galectins (Table 1). Notably, I. scapularis shows a significant expansion of galectins, compared to the other target species, which have diversified into three distinct clusters (Figure 3).
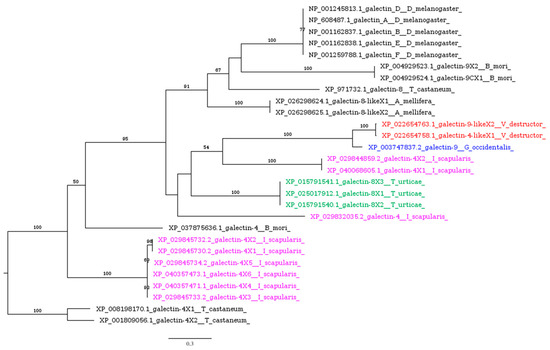
Figure 3.
Phylogenetic tree based on maximum likelihood analysis of galectin putative homologs identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values >50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
3.2.4. Thioester-Containing Proteins
Thioester-containing proteins (TEPs) are a family of proteins that show similarities to vertebrate alpha-2 macroglobulins (A2M) and complement factors [73,74]. Seven members of this family are encoded in the human genome: C3, C4, C5, A2M, pregnancy zone protein (PZP), CD109, and the complement 3 and PZP-like A2M domain-containing 8 (CPAMD8) [75,76,77]. As seen in complement proteins, certain insect TEPs play a role in the opsonization of microbes and pathogens, effectively “labeling” them for phagocytosis, melanization, and the formation of lytic complexes [78,79]. Given their participation in microbe recognition, TEPs are categorized as pattern recognition receptors (PRRs) [80].
Our phylogenetic analysis revealed a distinct clustering of A2M, CD109, and C3 homologs (Figure 4). All Acarina species investigated had at least one representative in each cluster, except for T. urticae, which lacks putative homologs of A2M. Notably, the best BLASTp hits of Drosophila TEP1, TEP2, TEP3, and TEP4 in the V. destructor genome are two isoforms of the same gene (Table 1).
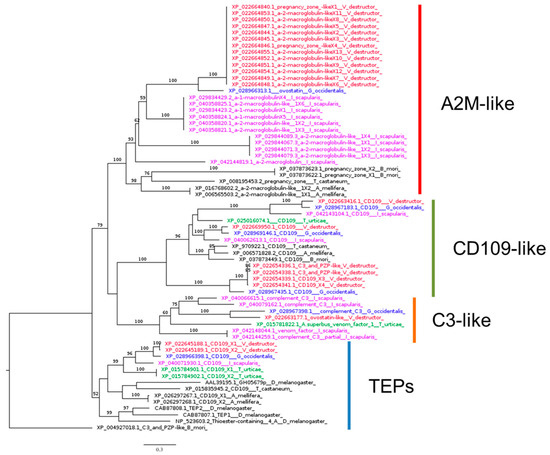
Figure 4.
Phylogenetic tree based on maximum likelihood analysis of thioester-containing protein (TEP) identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
3.3. Signaling Pathways
Our analysis revealed the occurrence of protein sequences encoded by the V. destructor genome with significant matches with genes of Drosophila involved in signaling. Among these, four candidate signaling genes were not transcribed in adult female mites (Supplementary File S1). As occurs in other mites and ticks (Tables S1–S3), the V. destructor genome includes a complete Toll pathway, while it lacks some members of IMD (IMD, dFADD, TAB2), JAK/STAT (unpaired), and JNK (Eiger) signaling pathways, which are present in Drosophila genomes (Table 2).
Table 2.
Immune genes of Varroa destructor involved in signaling. Orthologs not found in V. destructor are in italics.
Table 2.
Immune genes of Varroa destructor involved in signaling. Orthologs not found in V. destructor are in italics.
| Gene Name | Role | D. melanogaster | V. destructor 1 | E-Value | Identity | Coverage | Transcript |
|---|---|---|---|---|---|---|---|
| Spz1-1, spätzle 1B | Toll pathway | NP_733188.1 | XP_022644257.1 | 6 × 10−7 | 25.32% | 62% | NO |
| Spz1-2, spätzle 1Bii | Toll pathway | NP_001138116.1 | XP_022644257.1 | 2 × 10−7 | 25.32% | 52% | NO |
| Spz2, spätzle 2, neurotrophin 1 | Toll pathway | NP_001261417.1 | XP_022654810.1 | 3 × 10−15 | 27.85% | 14% | YES |
| Spz3, spätzle 3 | Toll pathway | NP_609160.2 | XP_022643975.1 | 3 × 10−61 | 40.00% | 43% | NO |
| Spz4, spätzle 4 | Toll pathway | NP_609504.2 | XP_022644415.1 | 5 × 10−51 | 63.55% | 17% | YES |
| Spz5, spätzle 5 | Toll pathway | NP_647753.1 | XP_022663991.1 | 1 × 10−14 | 38.98% | 29% | YES |
| Spz6, spätzle 6 | Toll pathway | NP_611961.1 | XP_022645393.1 | 2 × 10−30 | 63.75% | 18% | YES |
| Toll-1, protein Toll | Toll pathway | NP_524518.1 | XP_022664896.1 | 3 × 10−93 | 29.56% | 74% | YES |
| 18 wheeler, Toll-2 | Toll pathway | NP_476814.1 | XP_022651559.1 | 0.0 | 38.74% | 83% | YES |
| Toll-6 | Toll pathway | NP_001246766.1 | XP_022653722.1 | 0.0 | 40.50% | 77% | NO |
| Toll-7 | Toll pathway | NP_523797.1 | XP_022656113.1 | 0.0 | 41.21% | 82% | YES |
| Tollo, Toll-8 | Toll pathway | NP_524757.1 | XP_022651559.1 | 0.0 | 40.47% | 89% | YES |
| Tube 2, interleukin-1 receptor-associated kinase 4 | Toll pathway | NP_001189164.1 | Not found. A putative homolog was found by using the query from the T. urticae genome 2. | Not found | Not found | Not found | - |
| Myd88, myeloid differentiation primary response gene 2 | Toll pathway | AAF58953.1 | Not found. A putative homolog was found by using the query from the T. urticae genome 2. | Not found | Not found | Not found | - |
| Pll, pelle | Toll pathway | AAF56686.1 | XP_022652064.1 | 1 × 10−64 | 42.48% | 60% | YES |
| Cact, cactus | Toll pathway | AAN10936.1 | XP_022667531.1 | 8 × 10−17 | 30.21% | 48% | YES |
| Cactin | Toll pathway | NP_523422.4 | XP_022668977.1 | 0.0 | 52.09% | 82% | YES |
| Pli, pellino | Toll pathway | NP_524466.1 | XP_022668132.1 | 8 × 10−68 | 42.61% | 59% | YES |
| Traf1, TNF-receptor-associated factor 1 | Toll pathway | AAD34346.1 | XP_022668810.1 | 8 × 10−101 | 46.20% | 66% | YES |
| Traf2,TNF-receptor-associated factor 2 | Toll pathway | AAF46338.1 | XP_022668812.1 | 7 × 10−9 | 24.84% | 30% | YES |
| Traf3, TNF-receptor-associated factor 3 | Toll pathway | NP_727976.1 | XP_022687269.1 | 2 × 10−15 | 25.00% | 38% | YES |
| Dl, dorsal | Toll pathway | AAF53611.1 | XP_022665229.1 | 8 × 10−87 | 50.37% | 39% | YES |
| Dome, domeless 1, interleukine JAK/STAT receptor | JAK/STAT pathway | CAD12503.1 | XP_022659768.1 | 2 × 10−21 | 21.79% | 39% | YES |
| Hops, hopscotch, Janus kinase | JAK/STAT pathway | NP_511119.2 | XP_022650171.1 | 2 × 10−52 | 42.59% | 61% | YES |
| STAT92E, signal transducer and activator of transcription, marelle | JAK/STAT pathway | AAX33462.1 | XP_022661521.1 | 1 × 10−129 | 36.85% | 94% | YES |
| unpaired 1 | JAK/STAT pathway | NP_525095.2 | Not found | Not found | Not found | Not found | - |
| unpaired 2 | JAK/STAT pathway | NP_001356882.1 | Not found | Not found | Not found | Not found | - |
| unpaired 3 | JAK/STAT pathway | NP_001097014.1 | Not found | Not found | Not found | Not found | - |
| IMD, immune deficiency | IMD pathway | NP_573394.1 | Not found | Not found | Not found | Not found | - |
| dFADD | IMD pathway | NP_651006.1 | Not found | Not found | Not found | Not found | - |
| Dredd, death-related ced-3, caspase-1 | IMD pathway | NP_477249.3 | XP_022672682.1 | 2 × 10−17 | 28.98% | 46% | YES |
| Rel, Relish 3 | IMD pathway | NP_477094.1 | XP_022656314.1 | 3 × 10−11 | 33.81% | 14% | YES |
| TAB2, TAK1-associated binding protein 2 | IMD pathway | NP_611408.2 | Not found | Not found | Not found | Not found | - |
| TAK1, TGF-β-activated kinase 1 | IMD pathway | AAF50895.1 | XP_022652311.1 | 8 × 10−81 | 41.39% | 64% | YES |
| Key, Kenny | IMD pathway | NP_523856.2 | Not found | Not found | Not found | Not found | - |
| DIAP2/XIAP, death-associated inhibitor of apoptosis 2 | IMD pathway | NP_477127.1 | XP_022671835.1 | 2 × 10−59 | 31.12% | 86% | YES |
| IRD5, immune response deficiency 5, IK-β, IKKB, I-kappa-B kinase beta | IMD pathway | NP_524751.3 | XP_022693386.1 | 9 × 10−37 | 38.05% | 31% | YES |
| Hep, hemipterous | JNK pathway | NP_727661.1 | XP_022657489.1 | 4 × 10−110 | 56.25% | 25% | YES |
| Bsk, basket | JNK pathway | P92208.1 | XP_022646497.1 | 0.0 | 84.72% | 96% | YES |
| Jra, Jun-related antigen | JNK pathway | AAF58845.1 | XP_022649508.1 | 2 × 10−32 | 34.36% | 78% | YES |
| Kay, kayak | JNK pathway | NP_001027579.1 | XP_022661416.1 | 2 × 10−10 | 41.44% | 23% | YES |
| Egr, Eiger | JNK pathway | AAF58848.2 | Not found | Not found | Not found | Not found | - |
1 Best hit on the E-Value basis obtained by BLASTp searches against the NCBI nr database. 2 See Table S5. 3 This sequence does not contain Relish domains and it cannot be considered a Relish homolog (see Section 3.3.3).
3.3.1. The Toll Signaling Pathway
The Toll pathway in Drosophila plays a pivotal role in both development and innate immunity. The deletion of its constituent genes enhances susceptibility to a range of pathogens, including Gram-positive bacteria, fungal infections, certain Gram-negative bacteria, and viruses [81]. The Toll pathway appears to be intact in V. destructor (Table 2) and other Acarina species (Tables S1–S3), according to other studies on chelicerates immunity-related genes [30,41]. We found convincing matches for genes encoding the extracellular cytokine spätzle, the transmembrane receptor Toll (Figure 5), the kinase pelle, cactin, pellino, Traf, and the transactivator dorsal (Table 2).
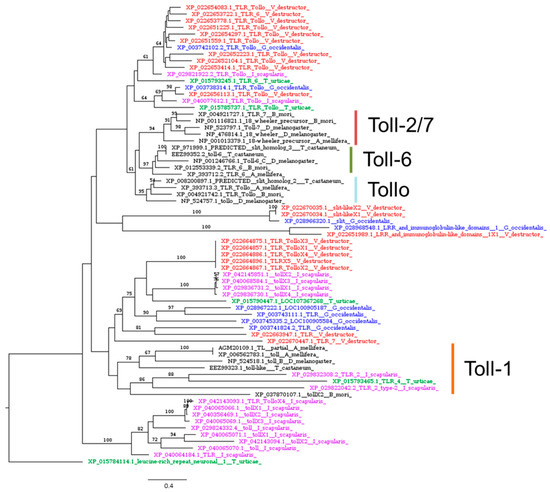
Figure 5.
Phylogenetic tree based on maximum likelihood analysis of Toll-1, Toll-6, Toll-7, and Tollo putative homologs identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
Moreover, our findings include a putative homolog of MyD88 and the tube adaptor in the Acarina species studied (Table S5). In invertebrates, MyD88 together with the adaptor tube form a complex with the kinase pelle, which in turn leads to the activation of cactus, and ultimately to the activation of the NF-κB ortholog dorsal [82]. Notably, Palmer and Jiggins, in their comparative genomics study, failed to find a tube homolog in any of the species studied, suggesting that the cause might be a lack of the power needed to detect the gene [30]. Indeed, in another comparative study the identification of the tube orthologs in arachnids was possible only by using the tube sequence from the mosquito A. aegypti, while no matches were obtained using the Drosophila ortholog as the query [41]. Similarly, we found that BLASTp searches using the Drosophila tube sequence as the query revealed no hits in V. destructor (Table 1), I. scapularis (Table S1), and G. occidentalis (Table S2) genomes, while a significant match was found in T. urticae (Table S3). Using this sequence as an alternative query, we identified significant matches in our target species, suggesting the presence of a tube homolog in the genomes of Acarina, including V. destructor (Table S5). However, tube and pelle are members of the same gene family, both having a death domain and a kinase domain, and orthology is difficult to infer.
Acarina also possess multiple genes encoding Toll receptors (Figure 5), which function as transmembrane receptors in both vertebrates and invertebrates [83,84]. While nine single-copy Toll genes have been identified in D. melanogaster (Toll-1 to Toll-9), it appears that Acarina, like other arthropods, lack some of these genes but have multiple isoforms of others. Indeed, V. destructor exhibited a highly diversified repertoire of Toll putative homologs (Figure 5). Toll-like sequences in Acarina form monophyletic groups (Figure 5), suggesting that diversification of the Toll family occurred after the evolution of the separate lineages of Acarina and insects. This is in line with other phylogenetic analyses, which indicated that Toll receptor genes from different phyla fall into separate clusters, showing that they share a common ancestor but evolved independently by gene duplication [85,86]. Our results suggest a functional divergence between Acarina and insect Toll receptors, which may be disclosed only by functional data.
3.3.2. The JAK/STAT Signaling Pathway
In Drosophila, the JAK/STAT pathway, much like the Toll pathway, plays pivotal roles in both development and immune responses. Although it remains the least detailed among the core insect immune pathways, it appears to be involved in hemocyte overproliferation and antiviral defense [87]. Similar to other Acarina species (Tables S1–S3), V. destructor possesses homologs of all core JAK/STAT genes, including those encoding the cytokine receptor domeless, JAK tyrosine kinase (also referred to as hopscotch), and the STAT92E transcription factor (Table 2). However, homologs for upd (unpaired), a crucial ligand for JAK/STAT activation in Drosophila, were not identified. This ligand is also absent in other insects, such as A. mellifera [88] and Coccidae [45]. The presence of the core JAK/STAT components (Table 2) suggests that the JAK/STAT pathway remains functional in Acarina, activated by an as-yet-unidentified ligand.
3.3.3. IMD and JNK Signaling Pathways
The immune deficiency (IMD) pathway is critical for fighting Gram-negative bacteria in Drosophila [81], and IMD pathway member knockouts influence the susceptibility to some Gram-positive bacteria and fungi as well [89]. In Drosophila, the pathway-initiating receptors, PGRPs, engage the adapter proteins IMD and FADD (Fas-associated protein with death domain) [90,91], with FADD subsequently interacting with Dredd (caspase-8) [92], which in turn cleaves IMD. The K63 polyubiquitylation of IMD is facilitated by the E3 ubiquitin ligase IAP2 (inhibitor of apoptosis 2) and the E2-conjugating enzymes Bendless, Uev1a, and Effete [93,94,95]. This facilitates the binding of Kenny to the inhibitor of NF-κB kinase (IKK)β [92,96]. Additionally, Dredd mediates the cleavage of Relish, leading to the nuclear translocation of its N-terminal fragment, which in turn triggers the expression of antimicrobial peptides (AMPs) [95,97,98,99]. The resulting signaling scaffold leads to cleavage of the NF-κB signaling molecule Relish, which translocates to the nucleus and promotes antimicrobial peptide (AMP) expression [93,95].
As in other Acarina species [100], V. destructor appears to be missing many crucial components of the IMD signaling pathway, such as IMD, dFADD, Kenny, and TAB2 (Table 2). However, the Relish search resulted in a positive hit characterized by only 14% coverage (Table 2), which was considered low. Indeed, InterProScan analysis revealed that similarities between the Relish gene in D. melanogaster and its putative homolog in V. destructor (XP_022656314.1) are only due to the Ankyrin domain’s presence in both sequences, while the V. destructor sequence lacks canonical Relish domains: the NF-kappa-B/Rel/dorsal domain (IPR011539) and Rel homology dimerization domain (IPR032397) [101,102]. However, the putative Relish homolog identified in I. scapularis includes Relish domains and was used as an alternative query to scan the V. destructor genome. This led to the identification of a significantly similar sequence described as proto-oncogene c-Rel-like (XP_022665225.1), showing Relish domains and lacking the Ankyrin domain, that is believed to inhibit its own nuclear translocation [97,102]. However, this loss of the Ankyrin domain, which occurs also in I. scapularis, G. occidentalis, and T. urticae, seems to not alter the functionality of Relish in ticks [103,104,105].
Using the Dredd protein sequence to search the V. destructor genome via BLASTp resulted in a best hit with 46% coverage (Table 2). However, this sequence cannot be considered a true homolog of Dredd (caspase-8) because it lacks the Dredd_N (IPR056259) and Dredd_2nd (IPR056260) domains (Figure 6). These findings align with previous studies reporting the absence of Dredd homologs in ticks and mites [41]. Dredd normally cleaves the loop sequence between the REL and Ankyrin domains of phosphorylated Relish [98,106]. Our results suggest that, in V. destructor and Acarina in general, the absence of Dredd homologs is likely due to reduced selective pressure on this caspase, stemming from the loss of its substrate: the Ankyrin domain in Relish homologs.
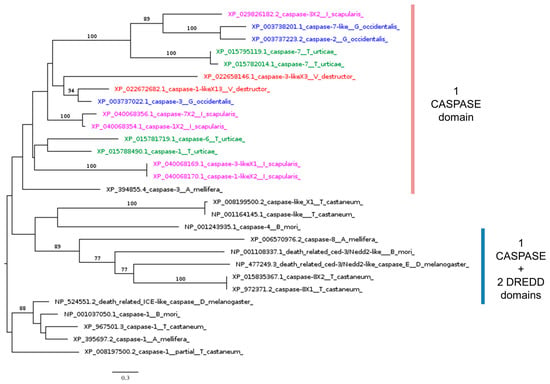
Figure 6.
Phylogenetic tree based on maximum likelihood analysis of Dredd putative homologs identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The Drice homolog sequences are used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
Despite the absence of upstream regulators, core IMD signaling molecules are active against infection, as reported by several studies on tick IMD pathways [107], which rely on Bendless, Effete, Uev1a, and XIAP (X-linked inhibitor of apoptosis), the homolog of IAP2 [43,108]. Our phylogenetic analysis indicated that all model Acarina species studied here, including V. destructor, possess homolog genes of the three ubiquitination-conjugating enzymes (UCEs): Bendless, Effete, and Uev1a (Figure 7).
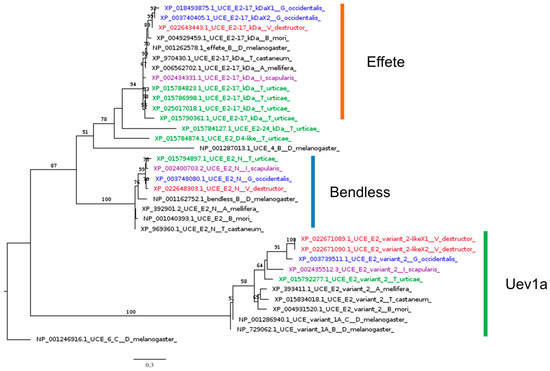
Figure 7.
Phylogenetic tree based on maximum likelihood analysis of ubiquitination-conjugating enzymes (UCEs) Bendless, Effete, and Uev1a, identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
In ticks, once activated, XIAP interacts with the heterodimers Bendless and Uev1a, leading to the ubiquitination of p47 in a K63-dependent manner [43,109]. Ubiquitylated p47 connects to Kenny and induces the phosphorylation of IRD5 and Relish [100]. This mechanism seems well conserved in Acarina. Indeed, we identified putative homologs of XIAP (Table 2) and p47 (Table S6) in our analysis.
Following the initiation of IMD signaling in Drosophila, various proteins are recruited to the molecular scaffold within the activated cell, including transforming growth factor-β-activated kinase 1 (TAK1). TAK1, which we identified also in V. destructor (Table 2), promotes activation of JNK signaling in parallel to NF-κB signaling [95,110].
In Drosophila, the JNK pathway is involved in various developmental processes, along with wound healing, and has been suggested to regulate antimicrobial peptide gene expression and cellular immune responses [81]. The putative homologs identified in V. destructor include kay (kayak), hep (hemipterous), bsk (basket), and jra (Jun-related antigen) (Table 2). Searches for homologs to the Drosophila Eiger gene found no hits in Acarina (Tables S1–S3), including V. destructor (Table 2), as observed also for other phyla such as Coccidae (Insecta, Hemiptera) [45].
3.4. Response Genes
Our analysis revealed the occurrence of several proteins encoded by the V. destructor genome with significant matches with genes of Drosophila involved in the immune response. Among these, only two candidate response genes were not transcribed in adult female mites (Supplementary File S1). As occurs in ticks and mites (Tables S1–S3), V. destructor lacks canonical antimicrobial peptides and prophenoloxidase orthologs, which are present in the D. melanogaster genome (Table 3).
Table 3.
Immune genes of Varroa destructor involved in the immune response. Orthologs not found in V. destructor are in italics.
Table 3.
Immune genes of Varroa destructor involved in the immune response. Orthologs not found in V. destructor are in italics.
| Gene Name | Role | D. melanogaster | V. destructor 1 | E-Value | Identity | Coverage | Transcript |
|---|---|---|---|---|---|---|---|
| Att, Attacin | antimicrobial peptide | NP_523745.1 | Not found | Not found | Not found | Not found | - |
| Cec, Cecropin | antimicrobial peptide | C0HKQ7.1 | Not found | Not found | Not found | Not found | - |
| Def, Defensin | antimicrobial peptide | ANY27112.1 | Not found | Not found | Not found | Not found | - |
| Dro, Dosocin | antimicrobial peptide | XP_016946682.1 | Not found | Not found | Not found | Not found | - |
| Mtk, Metchnikowin | antimicrobial peptide | AAO72489.1 | Not found | Not found | Not found | Not found | - |
| Andropin | antimicrobial peptide | P21663.1 | Not found | Not found | Not found | Not found | - |
| Diptericin | antimicrobial peptide | QER92349.1 | Not found | Not found | Not found | Not found | - |
| Drs, Drosomycin | antimicrobial peptide | ANY27466.1 | Not found | Not found | Not found | Not found | - |
| LysX, Lysozyme X, i-type | microbial degradation | CAL85493.1 | XP_022671742.1 | 2 × 10−30 | 38.30% | 96% | YES |
| LysB, Lysozyme B, i-type | microbial degradation | NP_001261245.1 | XP_022671739.1 | 2 × 10−33 | 41.01% | 97% | YES |
| LysP, Lysozyme, i-type | microbial degradation | NP_476828.1 | XP_022643422.1 | 4 × 10−37 | 43.28% | 95% | NO |
| LysE, Lysozyme E | microbial degradation | CAA80228 | XP_022669180.1 | 3 × 10−32 | 43.09% | 87% | YES |
| LysD, Lysozyme D | microbial degradation | NP_476823.1 | XP_022671742.1 | 7 × 10−33 | 43.90% | 87% | YES |
| LysE, Lysozyme E | microbial degradation | NP_476827.2 | XP_022671742.1 | 6 × 10−32 | 39.42% | 97% | YES |
| LysS, Lysozyme S | microbial degradation | NP_476829.1 | XP_022671742.1 | 1 × 10−34 | 44.36% | 95% | YES |
| Lysozyme E, i-type | microbial degradation | ACD99447.1 | XP_022648598.1 | 5 × 10−23 | 34.55% | 89% | YES |
| Lysozyme, i-type | microbial degradation | NP_611164.3 | XP_022655568.1 | 6 × 10−19 | 35.17% | 88% | YES |
| Lysozyme, i-type | microbial degradation | NP_611163.2 | XP_022644325.1 | 4 × 10−19 | 37.30% | 73% | YES |
| Cht2, Chitinase-like protein 4, flocculation protein | fungal degradation | NP_001261282.1 | XP_022662471.1 | 2 × 10−85 | 40.37% | 73% | YES |
| Cht4, Chitinase-like protein 2, mucin | fungal degradation | NP_524962.2 | XP_022662425.1 | 9 × 10−112 | 38.54% | 94% | YES |
| Cht5, Chitinase-like protein 5, endochitinase | fungal degradation | NP_650314.1 | XP_022664603.1 | 7 × 10−165 | 43.43% | 95% | YES |
| Cht7, Chitinase-like protein 7, chitinase 10 | fungal degradation | NP_647768.3 | XP_022669697.1 | 0.0 | 52.05% | 99% | NO |
| Cht6, Chitinase 6 | fungal degradation | NP_001245599.1 | XP_022662476.1 | 0.0 | 49.73% | 47% | YES |
| idgf6 | fungal degradation | NP_001286499.1 | XP_022662425.1 | 1 × 10−45 | 27.81% | 98% | YES |
| PPO1, Prophenoloxidase 1 | prophenoloxidase response | NP_476812.1 | Not found | Not found | Not found | Not found | - |
| PPO2, Prophenoloxidase 2 | prophenoloxidase response | NP_610443.1 | Not found | Not found | Not found | Not found | - |
| PAF2, Phenoloxidase-activating factor 2 | phenoloxidase activation | AAO24923.1 | XP_022659018.1 | 2 × 10−82 | 41.99% | 77% | YES |
| SP, Serine protease-like precursor | phenoloxidase activation | NP_001097766.1 | XP_022662993.1 | 2 × 10−43 | 37.84% | 56% | YES |
| Hmct, Hemolectin, hemocytin | cell aggregation | NP_001261809.1 | XP_022654008.1 | 8 × 10−158 | 32.23% | 66% | YES |
| Nos, Nitric oxide synthase | production of nitric oxide | NP_001027243.2 | XP_022665384.1 | 6 × 10−68 | 29.27% | 60% | YES |
| Tg, Transglutaminase | clotting | NP_609174.1 | XP_022666443.1 | 9 × 10−121 | 33.06% | 90% | YES |
1 Best hit on the E-Value basis obtained by BLASTp searches against the NCBI nr database.
3.4.1. Antimicrobial Peptides
Antimicrobial peptides (AMPs) are pivotal components of the immune defenses across a broad range of organisms, including insects. These peptides are among the most extensively researched humoral effectors and can be produced either constitutively or in response to specific signaling pathways. Typically composed of 15-to-50 amino acids, AMPs possess an amphipathic structure that enables them to disrupt microbial membrane integrity, thereby inducing alterations in the osmotic equilibrium and ultimately leading to lysis. The activation of the IMD and Toll signaling pathways in Drosophila triggers the transcription of distinct antimicrobial peptides [111]. At present, numerous structural families of insect-derived AMPs are identified, such as defensins, cecropins, drosocins, attacins, diptericins, metchnikowins, and melittins, some of which are exclusive to certain taxonomic groups [112]. However, none of these were identified in V. destructor and other Acarina species studied here by our BLASTp searches (Table 3). In contrast to our findings, defensins were reported from several tick species, including I. scapularis [113], suggesting that BLASTp is not the most appropriate method to identify these antimicrobial peptides in our target species. Indeed, we used the defensins identified in I. scapularis as alternative queries to scan the V. destructor proteome, which again resulted in no significant match. This limitation of BLASTp searches aimed at identifying AMPs was observed by other similar studies. As suggested, the small size of AMPs makes their identification by sequence similarity problematic across large phylogenetic distances [30].
Ctenidins are a family of constitutively expressed, glycine-rich AMPs identified from the spider Cupiennius salei that exhibit activity against Gram-negative bacteria [114]. BLASTp using ctenidin-like peptides identified in ticks as queries to scan the V. destructor genome led to the identification of putative homologs (Table S7). A similar pattern is observed for G. occidentalis, but not in T. urticae, where ctenidin putative homologs were not found (Table S7). These findings are particularly significant given the absence of widely adopted genetic readouts for immune pathways in mites [39].
3.4.2. Lysozyme
Lysozymes, enzymes tasked with degrading bacterial cell walls through the cleavage of the polysaccharide component of peptidoglycan, are categorized into two distinct classes in insects: the c-type, which exhibits muramidase activity, and the i-type, which possesses both muramidase and isopeptidase activities [115,116].
In ticks, lysozymes are expressed in the gut, salivary glands, and hemolymph, and are upregulated in response to bacterial challenge, suggesting a possible role of lysozymes: giving innate immunity to ticks against microorganisms [117]. Our analysis identified seven different proteins of V. destructor with significant similarity to Drosophila lysozymes (Table 3). Among these, only three (XP_022648598, XP_022655568, and XP_022644325) contain the invertebrate (I)-type lysozyme domain profile (LYSOZYME_I). Two of these (XP_022655568 and XP_022644325) also carry the EF hand calcium-binding domain (EF_HAND_1).
By filtering BLASTp results using the same criteria, we identified three lysozyme homologs in G. occidentalis and two in I. scapularis, carrying both LYSOZYME_I and EF_HAND_1. ScanProsite analysis revealed that among the sequences identified in T. urticae, large proteins (>400AA) possess multiple LYSOZYME_Is, while only one sequence (XP_015789674) showed a single LYSOZYME_I and was in the length range of canonical lysozymes (176AA). All these sequences identified in Acarina form a monophyletic group that diverges from the lysozymes with LYSOZYME_I and EF_HAND_1 identified in insect species (Figure 8).
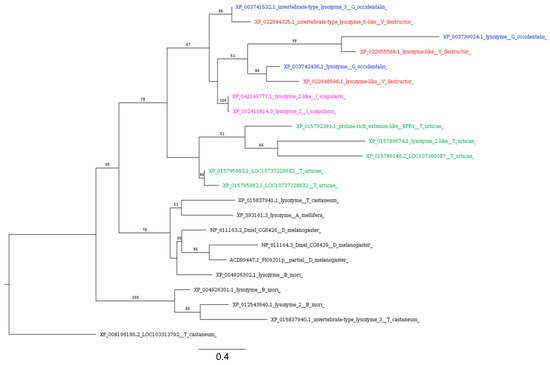
Figure 8.
Phylogenetic tree based on maximum likelihood analysis of i-type lysozymes identified in the genomes of V. destructor (in red), G. occidentalis (in blue), T. urticae (in green), and I. scapularis (in purple). The longest branch of the unrooted tree is used as the outgroup. Bootstrap support values > 50% are indicated at each node. The scale bar indicates the number of amino acid substitutions per site.
3.4.3. Melanization and Coagulation
Melanization, a key immune response in arthropods, involves the encapsulation of pathogens with melanin and their subsequent elimination through toxic quinone precursors [55,118]. Central to this melanization process is the enzyme prophenoloxidase (proPO, PPO), which, upon activation, transforms ortho-diphenols such as dopamine into quinones, precursors to melanin that ultimately encase and neutralize the pathogen. It is believed that in the common ancestor of arthropods, the oxygen-carrying molecule hemocyanin likely evolved from a protein resembling PO [119]. While PO has been detected in insects and crustaceans, specific forms of PO have not been identified in the studied chelicerates. Notably, in the horseshoe crab T. tridentatus, the oxygen-transporting protein hemocyanin exhibits PO activity upon activation [120]. Similarly, a hemocyanin with PO activity has been observed in the tarantula species Eurypelma californicum [121].
There is no evidence of the existence of the PPO cascade in ticks based on the available genomic and transcriptomic data [100]. In line with this, no PPO activity has been reported to be present in the hemolymph of the hard ticks Amblyomma americanum, Dermacentor variabilis, and I. scapularis [122]. Moreover, ticks lack coagulin and hemocyanin, suggesting a unique coagulation system in Acarina, which is largely different from that of insects [123]. Our analysis showed that this pattern occurs also in mites, including V. destructor, which lack PPO but possess transglutaminase homologs (Table 3). Considering these findings, additional studies are required to characterize melanization and the linked coagulation responses in Acarina.
4. Conclusions
Our results shed light on the genetic basis of Varroa and Acarina immunity, which is important for understanding the evolution of immune response in an economically important group of arthropods. While the genomic resources obtained through different sequencing technologies may result in different genome coverage, introducing bias in comparative analysis, we observed a clear pattern of immune gene loss in V. destructor, and chelicerates in general, supported by previous studies on arthropod immunity. Indeed, the immune gene repertoire of V. destructor matches the same pattern that several studies report for ticks, including the lack of transmembrane peptidoglycan recognition proteins (PGRPs), Gram-negative binding proteins, and several lectin families involved in recognition. Regarding signaling pathways, as occurs in ticks, Varroa mites lack homologs of the unpaired ligand (JAK/STAT), Eiger (JNK), and many components of IMD. Varroa mites also lack homologs of canonical AMPs, such as defensins, while their genome encodes putative homologs of ctenidins, AMPs previously identified in spiders and ticks. The same pattern is consistent in the predatory mite G. occidentalis, but not conserved in the phytophagous mite T. urticae, for which no canonical lectins nor ctenidins were identified. This suggests a greater degree of immune gene erosion in herbivorous mites, a trend that warrants further investigation.
Although our findings stem from a conservative homology-based approach using canonical insect immune genes, they represent the first comprehensive checklist of immunity-related genes in V. destructor. We hope this initial overview will inspire future studies to focus on the functional characterization of these genes, an essential step toward studying immune response in mites and developing novel strategies for pest control based on immunosuppression.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/insects16040356/s1, Supplementary File S1: tBLASTn results using candidate recognition genes as query vs. Varroa destructor transcriptome. Table S1: Immune genes involved in recognition, signaling and response in Ixodes scapularis. Table S2: Immune genes involved in recognition, signaling and response in Galendromus occidentalis. Table S3: Immune genes involved in recognition, signaling and response in Tetranychus urticae. Table S4: Results of BLASTp searches against Genbank nr protein database using lectin DL3 putative homolog from Varroa as query. Table S5: Results of BLASTp searches against Genbank nr protein database using tube and Myd88 putative homologs from Tetranychus urticae as query. Table S6: Results of BLASTp searches against Genbank nr protein database using p47 putative homologs from Ixodes scapularis as query. Table S7: Ctenidin-like proteins identified in V. destructor, Galendromus occidentalis and Tetranychus urticae.
Author Contributions
Conceptualization, A.B.; investigation, A.B., A.C. and G.D.L.; formal analysis, A.B. and I.D.L.; data curation, A.B. and G.D.L.; writing—original draft preparation, A.C. and A.B.; writing—review and editing, A.B., I.D.L., and A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All FASTA sequences used, alignments, and tree files are available from Zenodo at https://doi.org/10.5281/zenodo.14837927.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stokstad, E. The Case of the Empty Hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Carreck, N.L. Honey Bee Colony Losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.-P.; vanEngelsdorp, D. Drivers of Colony Losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Kielmanowicz, M.G.; Inberg, A.; Lerner, I.M.; Golani, Y.; Brown, N.; Turner, C.L.; Hayes, G.J.R.; Ballam, J.M. Prospective Large-Scale Field Study Generates Predictive Model Identifying Major Contributors to Colony Losses. PLoS Pathog. 2015, 11, e1004816. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and Control of Varroa destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. S1), S96–S119. [Google Scholar] [CrossRef]
- Noël, A.; Le Conte, Y.; Mondet, F. Varroa Destructor: How Does It Harm Apis mellifera Honey Bees and What Can Be Done about It? Emerg. Top. Life Sci. 2020, 4, 45–57. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor Feeds Primarily on Honey Bee Fat Body Tissue and Not Hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef]
- Han, B.; Wu, J.; Wei, Q.; Liu, F.; Cui, L.; Rueppell, O.; Xu, S. Life-History Stage Determines the Diet of Ectoparasitic Mites on Their Honey Bee Hosts. Nat. Commun. 2024, 15, 725. [Google Scholar] [CrossRef]
- De Jong, D.; De Jong, P.H. Longevity of Africanized Honey Bees (Hymenoptera: Apidae) Infested by Varroa jacobsoni (Parasitiformes: Varroidae). J. Econ. Entomol. 1983, 76, 766–768. [Google Scholar] [CrossRef]
- Bowen-Walker, P.L.; Gunn, A. The Effect of the Ectoparasitic Mite, Varroa destructor on Adult Worker Honeybee (Apis Mellifera) Emergence Weights, Water, Protein, Carbohydrate, and Lipid Levels. Entomol. Exp. Appl. 2001, 101, 207–217. [Google Scholar] [CrossRef]
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa Destructor: A Complex Parasite, Crippling Honey Bees Worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Kralj, J.; Fuchs, S. Parasitic Varroa destructor Mites Influence Flight Duration and Homing Ability of Infested Apis mellifera Foragers. Apidologie 2006, 37, 577–587. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an Ectoparasite on the Immunity and Pathology of an Invertebrate: Evidence for Host Immunosuppression and Viral Amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [PubMed]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.M.; Boots, M. Deformed Wing Virus Is a Recent Global Epidemic in Honeybees Driven by Varroa Mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef]
- Boecking, O.; Genersch, E. Varroosis—The Ongoing Crisis in Bee Keeping. J. Für Verbraucherschutz Und Leb. 2008, 3, 221–228. [Google Scholar] [CrossRef]
- Haber, A.I.; Steinhauer, N.A.; vanEngelsdorp, D. Use of Chemical and Nonchemical Methods for the Control of Varroa destructor (Acari: Varroidae) and Associated Winter Colony Losses in U.S. Beekeeping Operations. J. Econ. Entomol. 2019, 112, 1509–1525. [Google Scholar] [CrossRef]
- Lester, P.J. Integrated Resistance Management for Acaricide Use on Varroa destructor. Front. Bee Sci. 2023, 1, 1297326. [Google Scholar] [CrossRef]
- Way, M.J.; van Emden, H.F. Integrated Pest Management in Practice—Pathways towards Successful Application. Crop Prot. 2000, 19, 81–103. [Google Scholar] [CrossRef]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa destructor from the Laboratory to the Field: Control, Biocontrol and IPM Perspectives—A Review. Insects 2021, 12, 800. [Google Scholar] [CrossRef]
- Agrawal, N.; Dasaradhi, P.V.N.; Mohmmed, A.; Malhotra, P.; Bhatnagar, R.K.; Mukherjee, S.K. RNA Interference: Biology, Mechanism, and Applications. Microbiol. Mol. Biol. Rev. 2003, 67, 657–685. [Google Scholar] [CrossRef]
- Caccia, S.; Astarita, F.; Barra, E.; Di Lelio, I.; Varricchio, P.; Pennacchio, F. Enhancement of Bacillus thuringiensis Toxicity by Feeding Spodoptera littoralis Larvae with Bacteria Expressing Immune Suppressive dsRNA. J. Pest Sci. 2020, 93, 303–314. [Google Scholar] [CrossRef]
- Khila, A.; Grbić, M. Gene Silencing in the Spider Mite Tetranychus urticae: dsRNA and siRNA Parental Silencing of the Distal-Less Gene. Dev. Genes Evol. 2007, 217, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Shen, G.; Christiaens, O.; Smagghe, G.; He, L.; Wang, J. Beyond Insects: Current Status and Achievements of RNA Interference in Mite Pests and Future Perspectives. Pest Manag. Sci. 2018, 74, 2680–2687. [Google Scholar] [CrossRef] [PubMed]
- Garbian, Y.; Maori, E.; Kalev, H.; Shafir, S.; Sela, I. Bidirectional Transfer of RNAi between Honey Bee and Varroa Destructor: Varroa Gene Silencing Reduces Varroa Population. PLoS Pathog 2012, 8, e1003035. [Google Scholar] [CrossRef]
- McGruddy, R.A.; Smeele, Z.E.; Manley, B.; Masucci, J.D.; Haywood, J.; Lester, P.J. RNA Interference as a Next-Generation Control Method for Suppressing Varroa destructor Reproduction in Honey Bee (Apis mellifera) Hives. Pest Manag. Sci. 2024, 80, 4770–4778. [Google Scholar] [CrossRef]
- Leonard, S.P.; Powell, J.E.; Perutka, J.; Geng, P.; Heckmann, L.C.; Horak, R.D.; Davies, B.W.; Ellington, A.D.; Barrick, J.E.; Moran, N.A. Engineered Symbionts Activate Honey Bee Immunity and Limit Pathogens. Science 2020, 367, 573–576. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Tatè, R.; Campbell, E.M.; Gigliotti, S.; Bowman, A.S.; Pennacchio, F. A Salivary Chitinase of Varroa destructor Influences Host Immunity and Mite’s Survival. PLoS Pathog. 2020, 16, e1009075. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Cacace, A.; Parziale, M.; De Leva, G.; Iacopino, S.; Jesu, G.; Di Lelio, I.; Stillittano, V.; Caprio, E.; Pennacchio, F. The Salivary Gland Transcriptome of Varroa destructor Reveals Suitable Targets for RNAi-Based Mite Control. Insect Mol. Biol. 2024. [Google Scholar] [CrossRef]
- Pathak, J.P.N. Insect immunity; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 978-94-011-1618-3. [Google Scholar]
- Palmer, W.J.; Jiggins, F.M. Comparative Genomics Reveals the Origins and Diversity of Arthropod Immune Systems. Mol. Biol. Evol. 2015, 32, 2111–2129. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Zhang, R.; Zhang, J. The Diversity of Pattern Recognition Receptors (PRRs) Involved with Insect Defense against Pathogens. Curr. Opin. Insect Sci. 2019, 33, 105–110. [Google Scholar] [CrossRef]
- Zhao, L.; Niu, J.; Feng, D.; Wang, X.; Zhang, R. Immune Functions of Pattern Recognition Receptors in Lepidoptera. Front. Immunol. 2023, 14, 1203061. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Strand, M.R. The Insect Cellular Immune Response. Insect Sci. 2008, 15, 1–14. [Google Scholar]
- Eleftherianos, I.; Heryanto, C.; Bassal, T.; Zhang, W.; Tettamanti, G.; Mohamed, A. Haemocyte-Mediated Immunity in Insects: Cells, Processes and Associated Components in the Fight against Pathogens and Parasites. Immunology 2021, 164, 401–432. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway Jr, C.A. Innate Immunity: Impact on the Adaptive Immune Response. Curr. Opin. Immunol. 1997, 9, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patočka, J.; Kuča, K. Insect Antimicrobial Peptides, a Mini Review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tettamanti, G.; Bassal, T.; Heryanto, C.; Eleftherianos, I.; Mohamed, A. Regulators and Signalling in Insect Antimicrobial Innate Immunity: Functional Molecules and Cellular Pathways. Cell. Signal. 2021, 83, 110003. [Google Scholar]
- O’Neal, A.J.; Singh, N.; Rolandelli, A.; Laukaitis, H.J.; Wang, X.; Shaw, D.K.; Young, B.D.; Narasimhan, S.; Dutta, S.; Snyder, G.A.; et al. Croquemort Elicits Activation of the Immune Deficiency Pathway in Ticks. Proc. Natl. Acad. Sci. USA 2023, 120, e2208673120. [Google Scholar] [CrossRef]
- Smith, A.A.; Pal, U. Immunity-Related Genes in Ixodes scapularis—Perspectives from Genome Information. Front. Cell. Infect. Microbiol. 2014, 4, 116. [Google Scholar] [CrossRef]
- Bechsgaard, J.; Vanthournout, B.; Funch, P.; Vestbo, S.; Gibbs, R.A.; Richards, S.; Sanggaard, K.W.; Enghild, J.J.; Bilde, T. Comparative Genomic Study of Arachnid Immune Systems Indicates Loss of Beta-1,3-Glucanase-Related Proteins and the Immune Deficiency Pathway. J. Evol. Biol. 2016, 29, 277–291. [Google Scholar] [CrossRef]
- Rosa, R.D.; Capelli-Peixoto, J.; Mesquita, R.D.; Kalil, S.P.; Pohl, P.C.; Braz, G.R.; Fogaça, A.C.; Daffre, S. Exploring the Immune Signalling Pathway-Related Genes of the Cattle Tick Rhipicephalus microplus: From Molecular Characterization to Transcriptional Profile upon Microbial Challenge. Dev. Comp. Immunol. 2016, 59, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.K.; Wang, X.; Brown, L.J.; Chávez, A.S.O.; Reif, K.E.; Smith, A.A.; Scott, A.J.; McClure, E.E.; Boradia, V.M.; Hammond, H.L.; et al. Infection-Derived Lipids Elicit an Immune Deficiency Circuit in Arthropods. Nat. Commun. 2017, 8, 14401. [Google Scholar] [CrossRef]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; et al. Immunity and Other Defenses in Pea Aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Nicoletti, R.; Di Lelio, I.; Russo, E. Immune Gene Repertoire of Soft Scale Insects (Hemiptera: Coccidae). Int. J. Mol. Sci. 2024, 25, 4922. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinforma. Oxf. Engl. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum Likelihood-Based Phylogenetic Analyses with Thousands of Taxa and Mixed Models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- de Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE Signature Matches and ProRule-Associated Functional and Structural Residues in Proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM Predicts Alpha and Beta Transmembrane Proteins Using Deep Neural Networks. bioRxiv 2022. [Google Scholar]
- Kurata, S. Peptidoglycan Recognition Proteins in Drosophila Immunity. Dev. Comp. Immunol. 2014, 42, 36–41. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. Immune Properties of Invertebrate Phenoloxidases. Dev. Comp. Immunol. 2021, 122, 104098. [Google Scholar] [CrossRef] [PubMed]
- Franc, N.C.; Dimarcq, J.-L.; Lagueux, M.; Hoffmann, J.; Ezekowitz, R.A.B. Croquemort, A Novel Drosophila Hemocyte/Macrophage Receptor That Recognizes Apoptotic Cells. Immunity 1996, 4, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.-J.; Zhan, M.-Y.; Pan, Y.-M.; Liu, S.; Yang, P.-J.; Yang, L.-L.; Yu, X.-Q. Immune Functions of Insect βGRPs and Their Potential Application. Dev. Comp. Immunol. 2018, 83, 80–88. [Google Scholar] [CrossRef]
- Takahashi, D.; Garcia, B.L.; Kanost, M.R. Initiating Protease with Modular Domains Interacts with β-Glucan Recognition Protein to Trigger Innate Immune Response in Insects. Proc. Natl. Acad. Sci. USA 2015, 112, 13856–13861. [Google Scholar] [CrossRef]
- Warr, E.; Das, S.; Dong, Y.; Dimopoulos, G. The Gram-Negative Bacteria-Binding Protein Gene Family: Its Role in the Innate Immune System of Anopheles gambiae and in Anti-Plasmodium Defence. Insect Mol. Biol. 2008, 17, 39–51. [Google Scholar] [CrossRef]
- Pili-Floury, S.; Leulier, F.; Takahashi, K.; Saigo, K.; Samain, E.; Ueda, R.; Lemaitre, B. In Vivo RNA Interference Analysis Reveals an Unexpected Role for GNBP1 in the Defense against Gram-Positive Bacterial Infection in Drosophila Adults. J. Biol. Chem. 2004, 279, 12848–12853. [Google Scholar] [CrossRef]
- Gottar, M.; Gobert, V.; Matskevich, A.A.; Reichhart, J.-M.; Wang, C.; Butt, T.M.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual Detection of Fungal Infections in Drosophila via Recognition of Glucans and Sensing of Virulence Factors. Cell 2006, 127, 1425–1437. [Google Scholar] [CrossRef]
- Xia, X.; You, M.; Rao, X.-J.; Yu, X.-Q. Insect C-Type Lectins in Innate Immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, G.; Zhuo, X.; Liu, Y.; Tang, L.; Liu, X.; Wang, J. C-Type Lectin-Mediated Microbial Homeostasis Is Critical for Helicoverpa armigera Larval Growth and Development. PLoS Pathog. 2020, 16, e1008901. [Google Scholar] [CrossRef]
- Ao, J.; Ling, E.; Yu, X.-Q. Drosophila C-Type Lectins Enhance Cellular Encapsulation. Mol. Immunol. 2007, 44, 2541–2548. [Google Scholar] [CrossRef]
- Santos-Matos, G.; Wybouw, N.; Martins, N.E.; Zélé, F.; Riga, M.; Leitão, A.B.; Vontas, J.; Grbić, M.; Van Leeuwen, T.; Magalhães, S.; et al. Tetranychus urticae Mites Do Not Mount an Induced Immune Response against Bacteria. Proc. Biol. Sci. 2017, 284, 20170401. [Google Scholar] [CrossRef]
- Pace, K.E.; Baum, L.G. Insect Galectins: Roles in Immunity and Development. Glycoconj. J. 2002, 19, 607–614. [Google Scholar] [CrossRef]
- Drickamer, K. Two Distinct Classes of Carbohydrate-Recognition Domains in Animal Lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar]
- Dimopoulos, G.; Richman, A.; della Torre, A.; Kafatos, F.C.; Louis, C. Identification and Characterization of Differentially Expressed cDNAs of the Vector Mosquito, Anopheles Gambiae. Proc. Natl. Acad. Sci. USA 1996, 93, 13066–13071. [Google Scholar] [CrossRef] [PubMed]
- Kamhawi, S.; Ramalho-Ortigao, M.; Pham, V.M.; Kumar, S.; Lawyer, P.G.; Turco, S.J.; Barillas-Mury, C.; Sacks, D.L.; Valenzuela, J.G. A Role for Insect Galectins in Parasite Survival. Cell 2004, 119, 329–341. [Google Scholar] [CrossRef]
- Rao, X.-J.; Wu, P.; Shahzad, T.; Liu, S.; Chen, L.; Yang, Y.-F.; Shi, Q.; Yu, X.-Q. Characterization of a Dual-CRD Galectin in the Silkworm Bombyx Mori. Dev. Comp. Immunol. 2016, 60, 149–159. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary Dynamics of Immune-Related Genes and Pathways in Disease-Vector Mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, J.-L.; Cheng, Y.; Wang, J.-X.; Zou, Z. Pattern Recognition Receptors from Lepidopteran Insects and Their Biological Functions. Dev. Comp. Immunol. 2020, 108, 103688. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Thangamani, S.; Ho, B.; Ding, J.L. The Ancient Origin of the Complement System. EMBO J. 2005, 24, 382–394. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, R.; Fujito, N.T.; Nonaka, M. Evolution of the Thioester-Containing Proteins (TEPs) of the Arthropoda, Revealed by Molecular Cloning of TEP Genes from a Spider, Hasarius Adansoni. Dev. Comp. Immunol. 2012, 36, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Sottrup-Jensen, L.; Folkersen, J.; Kristensen, T.; Tack, B.F. Partial Primary Structure of Human Pregnancy Zone Protein: Extensive Sequence Homology with Human Alpha 2-Macroglobulin. Proc. Natl. Acad. Sci. USA 1984, 81, 7353–7357. [Google Scholar] [CrossRef]
- Lin, M.; Sutherland, D.R.; Horsfall, W.; Totty, N.; Yeo, E.; Nayar, R.; Wu, X.-F.; Schuh, A.C. Cell Surface Antigen CD109 Is a Novel Member of the A2 Macroglobulin/C3, C4, C5 Family of Thioester-Containing Proteins. Blood 2002, 99, 1683–1691. [Google Scholar] [CrossRef]
- Li, Z.-F.; Wu, X.; Engvall, E. Identification and Characterization of CPAMD8, a Novel Member of the Complement 3/A2-Macroglobulin Family with a C-Terminal Kazal Domain. Genomics 2004, 83, 1083–1093. [Google Scholar] [CrossRef]
- Lagueux, M.; Perrodou, E.; Levashina, E.A.; Capovilla, M.; Hoffmann, J.A. Constitutive Expression of a Complement-like Protein in Toll and JAK Gain-of-Function Mutants of Drosophila. Proc. Natl. Acad. Sci. USA 2000, 97, 11427–11432. [Google Scholar] [CrossRef]
- Levashina, E.A.; Moita, L.F.; Blandin, S.; Vriend, G.; Lagueux, M.; Kafatos, F.C. Conserved Role of a Complement-like Protein in Phagocytosis Revealed by dsRNA Knockout in Cultured Cells of the Mosquito, Anopheles Gambiae. Cell 2001, 104, 709–718. [Google Scholar] [CrossRef]
- Theopold, U.; Schmid, M. Thioester-Containing Proteins: At the Crossroads of Immune Effector Mechanisms. Virulence 2017, 8, 1468–1470. [Google Scholar] [CrossRef][Green Version]
- Boutros, M.; Agaisse, H.; Perrimon, N. Sequential Activation of Signaling Pathways during Innate Immune Responses in Drosophila. Dev. Cell 2002, 3, 711–722. [Google Scholar] [CrossRef]
- Towb, P.; Sun, H.; Wasserman, S.A. Tube Is an IRAK-4 Homolog in a Toll Pathway Adapted for Development and Immunity. J. Innate Immun. 2009, 1, 309–321. [Google Scholar] [CrossRef]
- Gay, N.J.; Gangloff, M.; Weber, A.N.R. Toll-like Receptors as Molecular Switches. Nat. Rev. Immunol. 2006, 6, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.J.; Gilmore, T.D. Evolutionary Origins of Toll-like Receptor Signaling. Mol. Biol. Evol. 2018, 35, 1576–1587. [Google Scholar] [CrossRef]
- Kanzok, S.M.; Hoa, N.T.; Bonizzoni, M.; Luna, C.; Huang, Y.; Malacrida, A.R.; Zheng, L. Origin of Toll-Like Receptor-Mediated Innate Immunity. J. Mol. Evol. 2004, 58, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Leulier, F.; Lemaitre, B. Toll-like Receptors—Taking an Evolutionary Approach. Nat. Rev. Genet. 2008, 9, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Agaisse, H.; Perrimon, N. The Roles of JAK/STAT Signaling in Drosophila Immune Responses. Immunol. Rev. 2004, 198, 72–82. [Google Scholar] [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.-L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune Pathways and Defence Mechanisms in Honey Bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef]
- Dionne, M.S.; Schneider, D.S. Models of Infectious Diseases in the Fruit Fly Drosophila melanogaster. Dis. Model. Mech. 2008, 1, 43–49. [Google Scholar] [CrossRef]
- Naitza, S.; Rossé, C.; Kappler, C.; Georgel, P.; Belvin, M.; Gubb, D.; Camonis, J.; Hoffmann, J.A.; Reichhart, J.M. The Drosophila Immune Defense against Gram-Negative Infection Requires the Death Protein dFADD. Immunity 2002, 17, 575–581. [Google Scholar] [CrossRef]
- Kaneko, T.; Yano, T.; Aggarwal, K.; Lim, J.-H.; Ueda, K.; Oshima, Y.; Peach, C.; Erturk-Hasdemir, D.; Goldman, W.E.; Oh, B.-H.; et al. PGRP-LC and PGRP-LE Have Essential yet Distinct Functions in the Drosophila Immune Response to Monomeric DAP-Type Peptidoglycan. Nat. Immunol. 2006, 7, 715–723. [Google Scholar] [CrossRef]
- Meinander, A.; Runchel, C.; Tenev, T.; Chen, L.; Kim, C.-H.; Ribeiro, P.S.; Broemer, M.; Leulier, F.; Zvelebil, M.; Silverman, N.; et al. Ubiquitylation of the Initiator Caspase DREDD Is Required for Innate Immune Signalling. EMBO J. 2012, 31, 2770–2783. [Google Scholar] [CrossRef] [PubMed]
- Paquette, N.; Broemer, M.; Aggarwal, K.; Chen, L.; Husson, M.; Ertürk-Hasdemir, D.; Reichhart, J.-M.; Meier, P.; Silverman, N. Caspase Mediated Cleavage, IAP Binding and Ubiquitination: Linking Three Mechanisms Crucial for Drosophila NF-κB Signaling. Mol. Cell 2010, 37, 172. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila Melanogaster—from Microbial Recognition to Whole-Organism Physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef]
- Kleino, A.; Silverman, N. The Drosophila IMD Pathway in the Activation of the Humoral Immune Response. Dev. Comp. Immunol. 2014, 42, 25–35. [Google Scholar] [CrossRef]
- Kleino, A.; Myllymäki, H.; Kallio, J.; Vanha-aho, L.-M.; Oksanen, K.; Ulvila, J.; Hultmark, D.; Valanne, S.; Rämet, M. Pirk Is a Negative Regulator of the Drosophila Imd Pathway. J. Immunol. Baltim. Md 1950 2008, 180, 5413–5422. [Google Scholar] [CrossRef] [PubMed]
- Stöven, S.; Ando, I.; Kadalayil, L.; Engström, Y.; Hultmark, D. Activation of the Drosophila NF-κB Factor Relish by Rapid Endoproteolytic Cleavage. EMBO Rep. 2000, 1, 347–352. [Google Scholar] [CrossRef]
- Stöven, S.; Silverman, N.; Junell, A.; Hedengren-Olcott, M.; Erturk, D.; Engström, Y.; Maniatis, T.; Hultmark, D. Caspase-Mediated Processing of the Drosophila NF-κB Factor Relish. Proc. Natl. Acad. Sci. USA 2003, 100, 5991–5996. [Google Scholar] [CrossRef]
- Ertürk-Hasdemir, D.; Broemer, M.; Leulier, F.; Lane, W.S.; Paquette, N.; Hwang, D.; Kim, C.-H.; Stöven, S.; Meier, P.; Silverman, N. Two Roles for the Drosophila IKK Complex in the Activation of Relish and the Induction of Antimicrobial Peptide Genes. Proc. Natl. Acad. Sci. USA 2009, 106, 9779–9784. [Google Scholar] [CrossRef]
- Fogaça, A.C.; Sousa, G.; Pavanelo, D.B.; Esteves, E.; Martins, L.A.; Urbanová, V.; Kopáček, P.; Daffre, S. Tick Immune System: What Is Known, the Interconnections, the Gaps, and the Challenges. Front. Immunol. 2021, 12, 628054. [Google Scholar] [CrossRef]
- Müller, C.W.; Rey, F.A.; Sodeoka, M.; Verdine, G.L.; Harrison, S.C. Structure of the NF-κB P50 Homodimer Bound to DNA. Nature 1995, 373, 311–317. [Google Scholar] [CrossRef]
- Dushay, M.S.; Asling, B.; Hultmark, D. Origins of Immunity: Relish, a Compound Rel-like Gene in the Antibacterial Defense of Drosophila. Proc. Natl. Acad. Sci. USA 1996, 93, 10343–10347. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, V.; Ayllón, N.; de la Lastra, J.M.P.; Galindo, R.C.; Kocan, K.M.; Blouin, E.F.; Mitra, R.; Alberdi, P.; Villar, M.; Fuente, J. de la Reciprocal Regulation of NF-kB (Relish) and Subolesin in the Tick Vector, Ixodes scapularis. PLoS ONE 2013, 8, e65915. [Google Scholar] [CrossRef]
- Capelli-Peixoto, J.; Carvalho, D.D.; Johnson, W.C.; Scoles, G.A.; Fogaça, A.C.; Daffre, S.; Ueti, M.W. The Transcription Factor Relish Controls Anaplasma marginale Infection in the Bovine Tick Rhipicephalus microplus. Dev. Comp. Immunol. 2017, 74, 32–39. [Google Scholar] [CrossRef]
- Fongsaran, C.; Verhoeve, V.I.; Jirakanwisal, K.; Harris, E.K.; Macaluso, K.R. Identification and Characterization of a Relish-Type NF-κB, DvRelish, in Dermacentor variabilis in Response to Rickettsia Rickettsii Infection. Front. Cell. Infect. Microbiol. 2024, 14, 1494450. [Google Scholar] [CrossRef]
- Jalovecka, M.; Malandrin, L.; Urbanova, V.; Mahmood, S.; Snebergerova, P.; Peklanska, M.; Pavlasova, V.; Sima, R.; Kopacek, P.; Perner, J.; et al. Activation of the Tick Toll Pathway to Control Infection of Ixodes ricinus by the Apicomplexan Parasite Babesia Microti. PLoS Pathog. 2024, 20, e1012743. [Google Scholar] [CrossRef]
- Oliva Chávez, A.S.; Shaw, D.K.; Munderloh, U.G.; Pedra, J.H.F. Tick Humoral Responses: Marching to the Beat of a Different Drummer. Front. Microbiol. 2017, 8, 223. [Google Scholar] [CrossRef]
- Severo, M.S.; Choy, A.; Stephens, K.D.; Sakhon, O.S.; Chen, G.; Chung, D.-W.D.; Le Roch, K.G.; Blaha, G.; Pedra, J.H.F. The E3 Ubiquitin Ligase XIAP Restricts Anaplasma phagocytophilum Colonization of Ixodes scapularis Ticks. J. Infect. Dis. 2013, 208, 1830–1840. [Google Scholar] [CrossRef]
- McClure Carroll, E.E.; Wang, X.; Shaw, D.K.; O’Neal, A.J.; Oliva Chávez, A.S.; Brown, L.J.; Boradia, V.M.; Hammond, H.L.; Pedra, J.H.F. P47 Licenses Activation of the Immune Deficiency Pathway in the Tick Ixodes scapularis. Proc. Natl. Acad. Sci. USA 2019, 116, 205–210. [Google Scholar] [CrossRef]
- Myllymäki, H.; Valanne, S.; Rämet, M. The Drosophila Imd Signaling Pathway. J. Immunol. Baltim. Md 1950 2014, 192, 3455–3462. [Google Scholar] [CrossRef]
- De Gregorio, E.; Spellman, P.T.; Tzou, P.; Rubin, G.M.; Lemaitre, B. The Toll and Imd Pathways Are the Major Regulators of the Immune Response in Drosophila. EMBO J. 2002, 21, 2568–2579. [Google Scholar] [CrossRef]
- Stączek, S.; Cytryńska, M.; Zdybicka-Barabas, A. Unraveling the Role of Antimicrobial Peptides in Insects. Int. J. Mol. Sci. 2023, 24, 5753. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhou, X.; Chen, Q.; Chen, Z.; Zhang, J.; Yang, L.; Sun, Y.; Wang, G.; Dai, J.; Feng, T. Defensins as a Promising Class of Tick Antimicrobial Peptides: A Scoping Review. Infect. Dis. Poverty 2022, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Baumann, T.; Kämpfer, U.; Schürch, S.; Schaller, J.; Largiadèr, C.; Nentwig, W.; Kuhn-Nentwig, L. Ctenidins: Antimicrobial Glycine-Rich Peptides from the Hemocytes of the Spider Cupiennius salei. Cell. Mol. Life Sci. 2010, 67, 2787–2798. [Google Scholar] [CrossRef]
- Bachali, S.; Jager, M.; Hassanin, A.; Schoentgen, F.; Jollès, P.; Fiala-Medioni, A.; Deutsch, J.S. Phylogenetic Analysis of Invertebrate Lysozymes and the Evolution of Lysozyme Function. J. Mol. Evol. 2002, 54, 652–664. [Google Scholar] [CrossRef]
- Van Herreweghe, J.M.; Michiels, C.W. Invertebrate Lysozymes: Diversity and Distribution, Molecular Mechanism and in Vivo Function. J. Biosci. 2012, 37, 327–348. [Google Scholar] [CrossRef]
- Tanaka, T.; Kawano, S.; Nakao, S.; Umemiya-Shirafuji, R.; Rahman, M.M.; Boldbaatar, D.; Battur, B.; Liao, M.; Fujisaki, K. The Identification and Characterization of Lysozyme from the Hard Tick Haemaphysalis longicornis. Ticks Tick-Borne Dis. 2010, 1, 178–185. [Google Scholar] [CrossRef]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-System: Pros and Cons for Its Role in Invertebrate Immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [PubMed]
- Burmester, T. Origin and Evolution of Arthropod Hemocyanins and Related Proteins. J. Comp. Physiol. B 2002, 172, 95–107. [Google Scholar] [CrossRef]
- Nagai, T.; Osaki, T.; Kawabata, S. Functional Conversion of Hemocyanin to Phenoloxidase by Horseshoe Crab Antimicrobial Peptides. J. Biol. Chem. 2001, 276, 27166–27170. [Google Scholar] [CrossRef]
- Voit, R.; Feldmaier-Fuchs, G.; Schweikardt, T.; Decker, H.; Burmester, T. Complete Sequence of the 24-Mer Hemocyanin of the Tarantula Eurypelma Californicum. Structure and Intramolecular Evolution of the Subunits. J. Biol. Chem. 2000, 275, 39339–39344. [Google Scholar] [CrossRef]
- Zhioua, E.; Browning, M.; Johnson, P.W.; Ginsberg, H.S.; LeBrun, R.A. Pathogenicity of the Entomopathogenic Fungus Metarhizium Anisopliae (Deuteromycetes) to Ixodes scapularis (Acari: Ixodidae). J. Parasitol. 1997, 83, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Simser, J.A.; Macaluso, K.R.; Mulenga, A.; Azad, A.F. Immune-Responsive Lysozymes from Hemocytes of the American Dog Tick, Dermacentor variabilis and an Embryonic Cell Line of the Rocky Mountain Wood Tick, D. Andersoni. Insect Biochem. Mol. Biol. 2004, 34, 1235–1246. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).