
Seasonal and Long-Term Population Dynamics of the Peach Fruit Fly in Egypt



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling of Peach Fruit Fly
2.2. Climate Data Collection
2.3. Data Analysis
3. Results
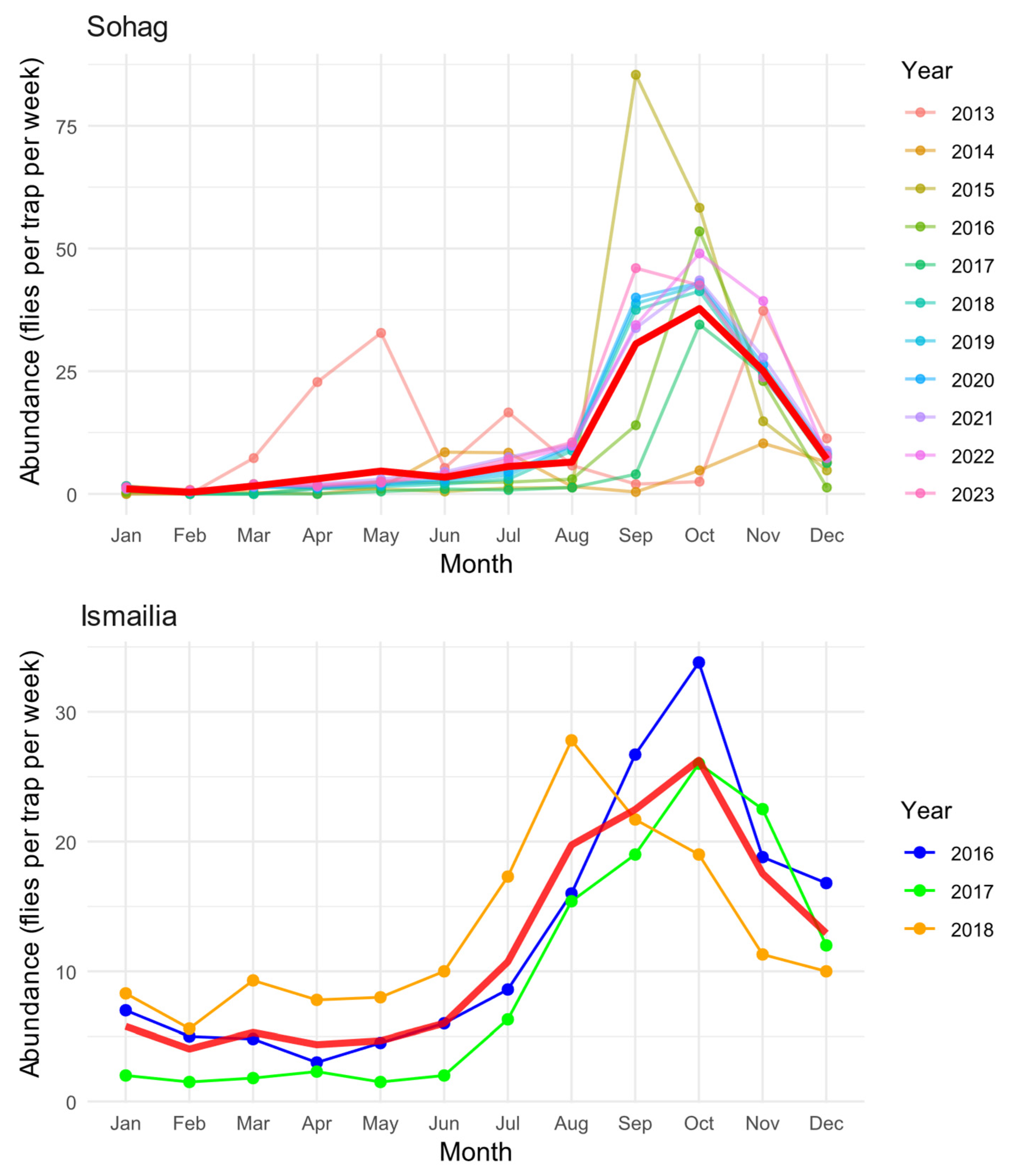
3.1. Seasonal Population Dynamics Across Regions
3.2. Factors Influencing Peach Fruit Fly Abundance
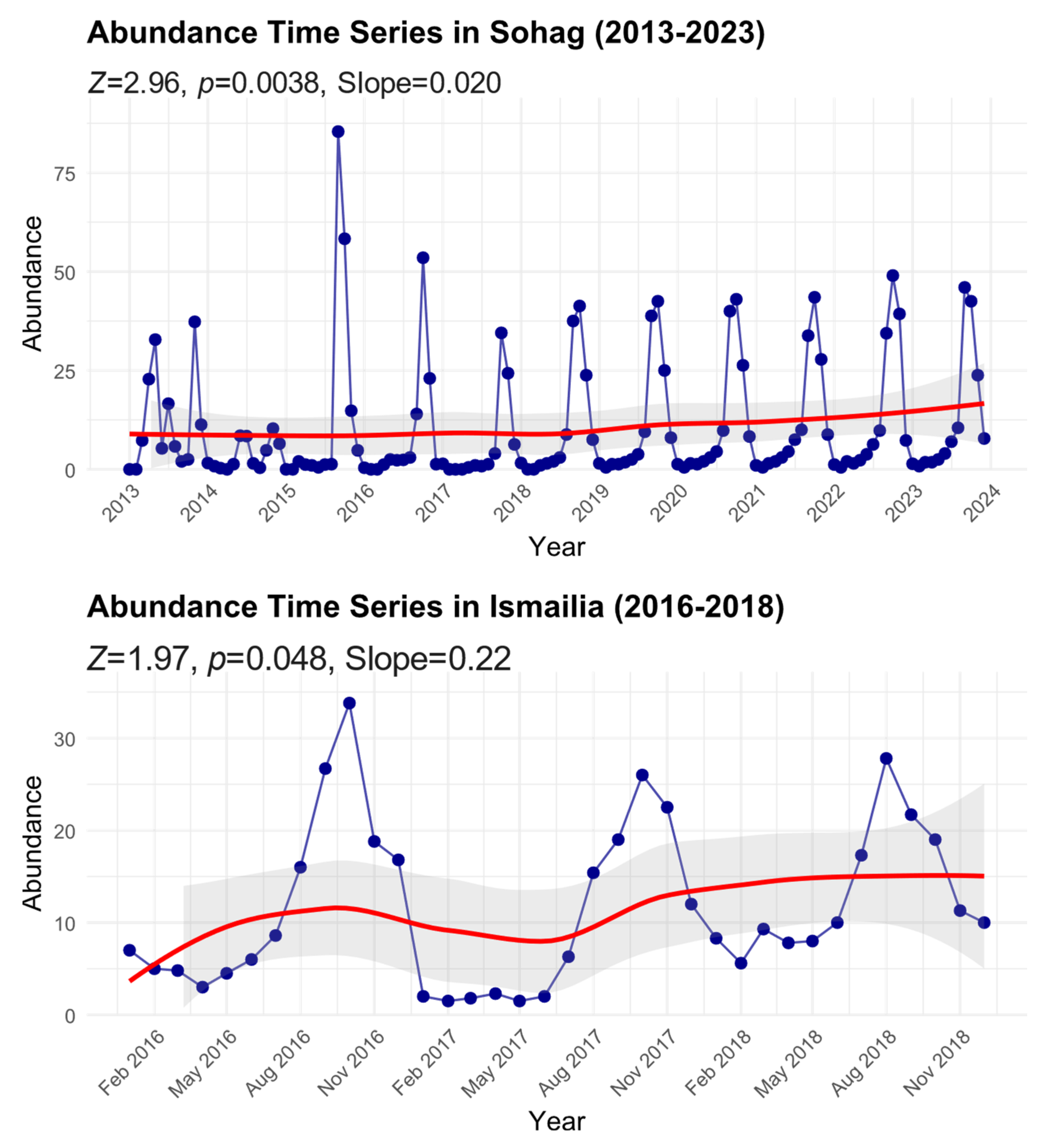
3.3. Long-Term Population Trends (2013–2023)
3.4. Temperature Patterns and Trends
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef]
- Cannon, R.J.C. The Implications of Predicted Climate Change for Insect Pests in the UK, with Emphasis on Non-Indigenous Species. Glob. Chang. Biol. 1998, 4, 785–796. [Google Scholar] [CrossRef]
- Zidon, R.; Tsueda, H.; Morin, E.; Morin, S. Projecting Pest Population Dynamics under Global Warming: The Combined Effect of Inter- and Intra-Annual Variations. Ecol. Appl. 2016, 26, 1198–1210. [Google Scholar] [CrossRef]
- Xue, Q.; Majeed, M.Z.; Zhang, W.; MA, C. Adaptation of Drosophila Species to Climate Change—A Literature Review since 2003. J. Integr. Agric. 2019, 18, 805–814. [Google Scholar] [CrossRef]
- Ni, W.L.; Li, Z.H.; Chen, H.J.; Wan, F.H.; Qu, W.W.; Zhang, Z.; Kriticos, D.J. Including Climate Change in Pest Risk Assessment: The Peach Fruit Fly, Bactrocera zonata (Diptera: Tephritidae). Bull. Entomol. Res. 2012, 102, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Zingore, K.M.; Sithole, G.; Abdel-Rahman, E.M.; Mohamed, S.A.; Ekesi, S.; Tanga, C.M.; Mahmoud, M.E.E. Global Risk of Invasion by Bactrocera zonata: Implications on Horticultural Crop Production under Changing Climatic Conditions. PLoS ONE 2020, 15, e0243047. [Google Scholar] [CrossRef]
- Choudhary, J.S.; Mali, S.S.; Naaz, N.; Malik, S.; Das, B.; Singh, A.K.; Srinivasa Rao, M.; Bhatt, B.P. Spatio and Temporal Variations in Population Abundance and Distribution of Peach Fruit Fly, Bactrocera zonata (Saunders) during Future Climate Change Scenarios Based on Temperature Driven Phenology Model. Clim. Risk Manag. 2021, 32, 100277. [Google Scholar] [CrossRef]
- Hassanein, R.; Romeh, A.; El-Sharkawy, H.; El-Nagar, H. Studies on the Population Fluctuations of the Peach Fruit Fly, Bactrocera zonata in Mango and Citrus Orchards and Its Field Control in Sharkia Governorate, Egypt. J. Product. Dev. 2024, 29, 57–79. [Google Scholar]
- Ullah, F.; Zhang, Y.; Gul, H.; Hafeez, M.; Desneux, N.; Qin, Y.; Li, Z. Estimation of the Potential Geographical Distribution of Invasive Peach Fruit Fly under Climate Change by Integrated Ecological Niche Models. CABI Agric. Biosci. 2023, 4, 46. [Google Scholar] [CrossRef]
- Elnagar, S.; El-Sheikh, M.A.; Hashem, A.G.; Afia, Y.E. Recent Invasion by Bactrocera zonata (Saunders) as a New Pest Competing with Ceratitis capitata (Wiedemann) in Attacking Fruits in Egypt. Available online: https://api.semanticscholar.org/CorpusID:82635385 (accessed on 15 November 2024).
- Darwish, A.A.-F.E.-S. Relative Susceptibility of Some Fruits to the Mediterranean Fruit Fly, Ceratitis capitata (Wiedemann) and Peach Fruit Fly, Bactrocera zonata (Saunders) (Diptera: Tephritidae) in Egypt. J. Entomol. Zool. Stud. 2016, 4, 42–48. [Google Scholar]
- Elmahdy, S.; Afia, Y. Population Fluctuation of the Peach Fruit Fly, Bactrocera zonata (Saunders) and the Mediterranean Fruit Fly Ceratitis capitata (Wiedemann) (Diptera, Tephritidae) on Three Grape Varieties in Nubaria, Egypt. J. Plant Prot. Pathol. 2018, 9, 547–555. [Google Scholar] [CrossRef]
- Duyck, P.-F.; David, P.; Quilici, S. A Review of Relationships between Interspecific Competition and Invasions in Fruit Flies (Diptera: Tephritidae). Ecol. Entomol. 2004, 29, 511–520. [Google Scholar] [CrossRef]
- Stange, E.E.; Ayres, M.P. Climate Change Impacts: Insects. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010; ISBN 0470016175. [Google Scholar]
- Eickermann, M.; Junk, J.; Rapisarda, C. Climate Change and Insects. Insects 2023, 14, 678. [Google Scholar] [CrossRef]
- El-Gendy, I.; Nassar, A.; Abdel-Hafeez, T. Response of Peach Fruit Fly, Bactrocera zonata (Saunders) to the Essential Oil of Cubeb Pepper, Piper Cubeba Bojer. Egypt. Acad. J. Biol. Sci. A Entomol. 2020, 13, 283–293. [Google Scholar] [CrossRef]
- Amin, R.; Nancy, N.P.; Miah, R.U.; Miah, M.G.; Kwon, O.; Suh, S.H. Fluctuations in Fruit Fly Abundance and Infestation in Sweet Gourd Fields in Relation to Varied Meteorological Factors. Entomol. Res. 2019, 49, 223–228. [Google Scholar] [CrossRef]
- El-Gepaly, H.; Sallam, G.; Mohamed, A.; Abdel-Aziz, S. Occurrence and Abundance of Spiders in Various Agricultural Formations at Sohag Governorate, Egypt. Acarines J. Egypt. Soc. Acarol. 2018, 12, 45–55. [Google Scholar] [CrossRef]
- Bous, M.E. Hoda Species Distribution Patterns of the Weed Flora in Mango Orchards of Ismailia Governorate, Egypt:implications for Conservation. Taeckholmia 2018, 38, 184–200. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Hancock, D.L. The Bactrocera Dorsalis Complex of Fruit Flies (Diptera: Tephritidae: Dacinae) in Asia. Bull. Entomol. Res. Suppl. Ser. 1994, 2, 1–68. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Tsuruta, K.; White, I.M. A New Species of Pest Fruit Fly (Diptera: Tephritidae: Dacinae) from Sri Lanka and Africa. Afr. Entomol. 2005, 13, 149–154. [Google Scholar]
- Soliman, M.M.; Malash, A.A.; El-Hawagry, M.S. Seasonal Abundance of Heterotrioza chenopodii (Reuter, 1876) and Distribution of the Known Psylloid Species (Hemiptera: Psylloidea) in Egypt. Afr. Entomol. 2021, 29, 201–211. [Google Scholar] [CrossRef]
- Child, R.E. Insect Damage as a Function of Climate. In Museum Microclimates; National Museum of Denmark: Copenhagen, Denmark, 2007. [Google Scholar]
- Arif, M.; Aasi, M.S.; Farooq, M.; Jian, W.Z. Spatio-Temporal Distribution of the Peach Fruit Fly, Bactrocera zonata (Diptera: Tephritidae) Infesting Citrus Orchards at Sargodha, Pakistan. Acta Entomol. Sin. 2017, 60, 1457–1466. [Google Scholar] [CrossRef]
- Harris, E.J.; Lee, C.Y.L. Seasonal and Annual Distribution of the Mediterranean Fruit Fly (Diptera: Tephritidae) in Honolulu and Suburban Areas of Oahu, Hawaii. Environ. Entomol. 1987, 16, 1273–1282. [Google Scholar] [CrossRef]
- Alberti, S.; Bogus, G.M.; Garcia, F.R.M. Flutuação Populacional de Moscas-Das-Frutas (Diptera, Tephritidae) Em Pomares de Pessegueiro E Maracujazeiro Em Iraceminha, Santa Catarina. Biotemas 2012, 25, 2. [Google Scholar] [CrossRef]
- Martínez-Ferrer, M.T.; Navarro, C.; Campos, J.M.; Marzal, C.; Fibla, J.M.; Bargues, L.; Garcia-Marí, F. Seasonal and Annual Trends in Field Populations of Mediterranean Fruit fly, Ceratitis capitata, in Mediterranean Citrus Groves: Comparison of Two Geographic Areas in Eastern Spain. Span. J. Agric. Res. 2010, 8, 757. [Google Scholar] [CrossRef]
- Aluja, M.; Ordano, M.; Guillén, L.; Rull, J. Understanding Long-Term Fruit Fly (Diptera: Tephritidae) Population Dynamics: Implications for Areawide Management. J. Econ. Entomol. 2012, 105, 823–836. [Google Scholar] [CrossRef]
- Dejene, B.T.; Wondimu, T.W.; Herrera, S.L.; Larsson, M.; Mafra-Neto, A.; Gessese, Y.W.; Dekker, T. Dispersal and Competitive Release Affect the Management of Native and Invasive Tephritid Fruit Flies in Large and Smallholder Farms in Ethiopia. Sci. Rep. 2021, 11, 2690. [Google Scholar] [CrossRef]
- Sarwar, M.; Hamed, M.; Yousaf, M.; Hussain, M. Surveillance on Population Dynamics and Fruits Infestation of Tephritid Fruit Flies (Diptera: Tephritidae) in Mango (Mangifera indica L.) Orchards of Faisalabad, Pakistan. Int. J. Sci. Res. Environ. Sci. 2014, 2, 113–119. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | χ2 | df | p | B | 95% CI | p |
|---|---|---|---|---|---|---|
| Month | 298.1 | 11 | <0.001 *** | |||
| Jan | −1.312 | −2.0–−0.60 | <0.001 *** | |||
| Feb | −2.024 | −2.7–−1.3 | <0.001 *** | |||
| Mar | −1.904 | −2.6–−1.1 | <0.001 *** | |||
| Apr | −2.158 | −3.1–−1.1 | <0.001 *** | |||
| May | −2.356 | −3.8–−0.87 | 0.002 ** | |||
| Jun | −2.520 | −4.2–−0.86 | 0.003 ** | |||
| Jul | −2.166 | −3.8–−0.46 | 0.012 * | |||
| Aug | −1.780 | −3.4–−0.10 | 0.037 * | |||
| Sep | −0.228 | −1.7–1.3 | 0.765 | |||
| Oct | 0.431 | −0.71–1.6 | 0.459 | |||
| Nov | 0.588 | −0.10–1.3 | 0.096 | |||
| Dec | Ref | – | – | |||
| Year | 41.4 | 10 | <0.001 *** | |||
| 2013 | 0.927 | 0.32–1.5 | 0.003 ** | |||
| 2014 | −0.522 | −1.2–0.14 | 0.127 | |||
| 2015 | −0.048 | −0.65–0.55 | 0.877 | |||
| 2016 | −0.453 | −1.0–0.11 | 0.120 | |||
| 2017 | −0.790 | −1.4–−0.19 | 0.009 ** | |||
| 2018 | −0.142 | −0.70–0.41 | 0.619 | |||
| 2019 | −0.022 | −0.63–0.59 | 0.944 | |||
| 2020 | 0.025 | −0.59–0.64 | 0.938 | |||
| 2021 | 0.118 | −0.48–0.72 | 0.702 | |||
| 2022 | 0.140 | −0.47–0.75 | 0.656 | |||
| 2023 | Ref | – | – | |||
| Zone | 24.6 | 1 | <0.001 *** | |||
| Sohag | −1.110 | −1.5–−0.67 | <0.001 *** | |||
| Ismailia | Ref | – | – | |||
| Temperature | 5.17 | 1 | 0.023 * | 0.124 | 0.01–0.23 | 0.023 * |
| Precipitation | 0.199 | 1 | 0.655 | 0.017 | −0.06–0.09 | 0.655 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, M.M.; EL-Solimany, E.A.; Hesselberg, T.; Negm, A.A.K.H. Seasonal and Long-Term Population Dynamics of the Peach Fruit Fly in Egypt. Insects 2025, 16, 332. https://doi.org/10.3390/insects16040332
Soliman MM, EL-Solimany EA, Hesselberg T, Negm AAKH. Seasonal and Long-Term Population Dynamics of the Peach Fruit Fly in Egypt. Insects. 2025; 16(4):332. https://doi.org/10.3390/insects16040332
Chicago/Turabian StyleSoliman, Mustafa M., Esmat A. EL-Solimany, Thomas Hesselberg, and Amira A. K. H. Negm. 2025. "Seasonal and Long-Term Population Dynamics of the Peach Fruit Fly in Egypt" Insects 16, no. 4: 332. https://doi.org/10.3390/insects16040332
APA StyleSoliman, M. M., EL-Solimany, E. A., Hesselberg, T., & Negm, A. A. K. H. (2025). Seasonal and Long-Term Population Dynamics of the Peach Fruit Fly in Egypt. Insects, 16(4), 332. https://doi.org/10.3390/insects16040332