
Plasticity in Caste-Fate Determination During the Adult Stage in Temperate Polistes Wasps



{kind=link}
Simple Summary
Abstract
1. Introduction
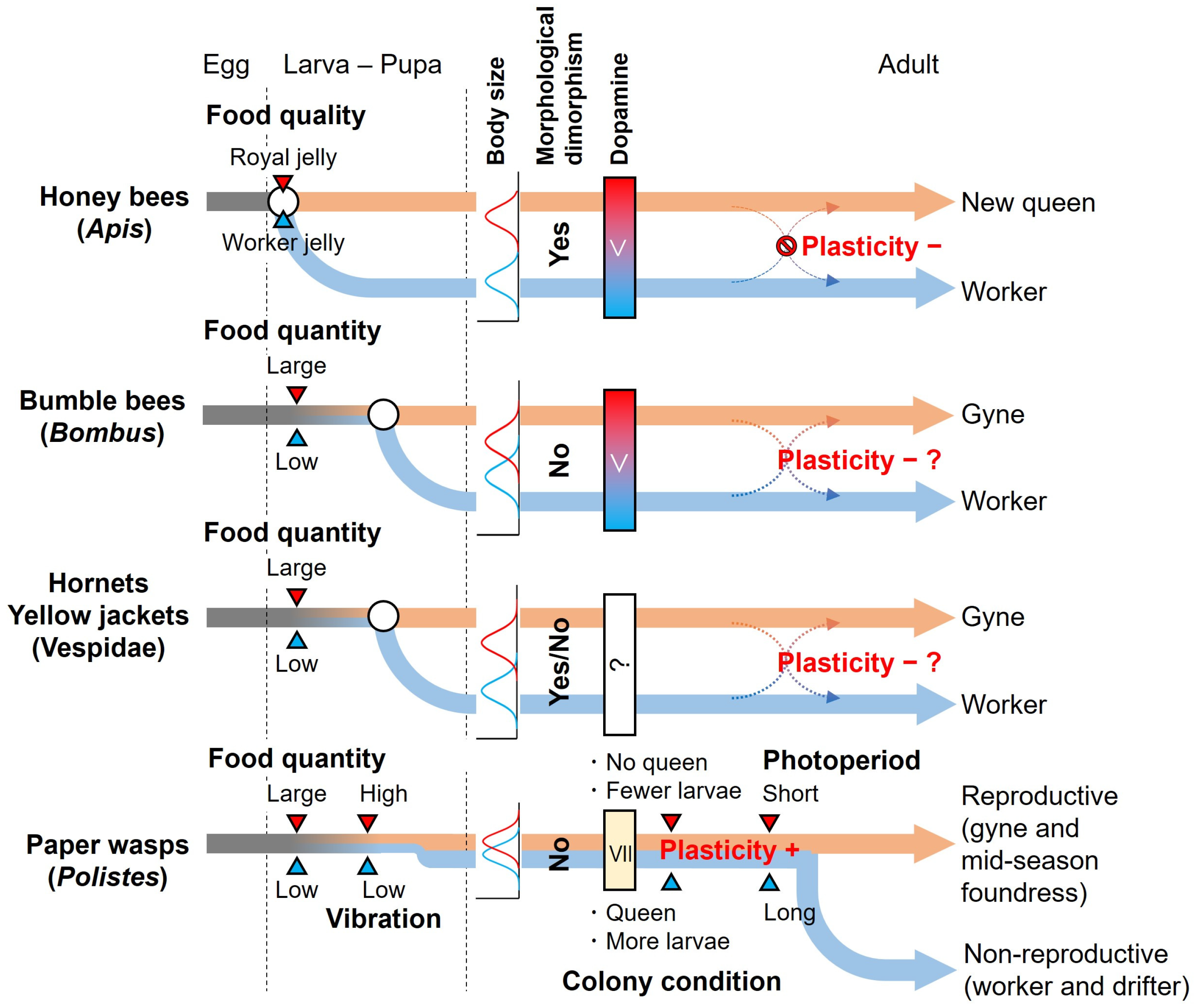
2. Caste Determination System in Temperate Polistes Wasps
2.1. Preimaginal Caste-Fate Biases
2.2. Imaginal Caste Determination Factors
3. Comparison of Caste Determination Systems Between Polistes Wasps and Other Eusocial Hymenoptera
4. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michener, C.D. Comparative social behavior of bees. Ann. Rev. Entomol. 1969, 14, 299–342. [Google Scholar]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Michener, C.D. The Social Behavior of the Bees; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Jeanne, R.L. Social complexity in the Hymenoptera, with special attention to the wasps. In Genes, Behaviors and Evolution of Social Insects; Kikuchi, T., Azuma, N., Higashi, S., Eds.; Hokkaido University Press: Sapporo, Japan, 2003; pp. 81–131. [Google Scholar]
- Jandt, J.M.; Toth, A.L. Physiological and genomic mechanisms of social organization in wasps (Family: Vespidae). Adv. Insect Physiol. 2015, 48, 95–130. [Google Scholar] [CrossRef]
- Gobin, B.; Ito, F.; Peeters, C.; Billen, J. Queen-worker differences in spermatheca reservoir of phylogenetically basal ants. Cell Tissue Res. 2006, 326, 169–178. [Google Scholar] [CrossRef]
- Khila, A.; Abouheif, E. Reproductive constraint is a developmental mechanism that maintains social harmony in advanced ant societies. Proc. Natl. Acad. Sci. USA 2008, 105, 17884–17889. [Google Scholar] [CrossRef]
- Kugler, J.; Orion, T.; Ishay, J. The number of ovarioles in the Vespinae (Hymenoptera). Insectes Sociaux 1976, 23, 525–533. [Google Scholar] [CrossRef]
- Ito, F.; Ohkawara, K. Spermatheca size differentiation between queens and workers in primitive ants. Naturwissenschaften 1994, 81, 138–140. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Groh, C.; Rössler, W. Caste-specific postembryonic development of primary and secondary olfactory centers in the female honeybee brain. Arthropod Struct. Dev. 2008, 37, 459–468. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Oldroyd, B.P.; Beekman, M.; Ratnieks, F.L.W. Ancestral monogamy shows kin selection is key to the evolution of eusociality. Science 2008, 320, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Piekarski, P.K.; Carpenter, J.M.; Lemmon, A.R.; Lemmon, E.M.; Sharanowski, B.J. Phylogenomic evidence overturns current conceptions of social evolution in wasps (Vespidae). Mol. Biol. Evol. 2018, 35, 2097–2109. [Google Scholar] [CrossRef]
- Hines, H.M. Historical biogeography, divergence times, and diversification patterns of bumble bees (Hymenoptera: Apidae: Bombus). Syst. Biol. 2008, 57, 58–75. [Google Scholar] [CrossRef]
- Dunn, R.R.; Agosti, D.; Andersen, A.N.; Arnan, X.; Bruhl, C.A.; Cerdá, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.F.; Payne, A.; Pickett, K.M.; Carpenter, J.M. Phylogeny and historical biogeography of the paper wasp genus Polistes (Hymenoptera: Vespidae): Implications for the overwintering hypothesis of social evolution. Cladistics 2015, 31, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Denlinger, D.L. Insect Diapause; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Eickwort, K. Separation of the castes of Polistes exclamans and notes on its biology (Hym.: Vespidae). Insectes Sociaux 1969, 16, 67–72. [Google Scholar] [CrossRef]
- Keeping, M.G. Reproductive and worker castes in the primitively eusocial wasp Belonogaster petiolata (DeGeer) (Hymenoptera: Vespidae): Evidence for pre-imaginal differentiation. J. Insect Physiol. 2002, 48, 867–879. [Google Scholar] [CrossRef]
- Toth, A.L.; Bilof, K.B.J.; Henshaw, M.T.; Hunt, J.H.; Robinson, G.E. Lipid stores, ovary development, and brain gene expression in Polistes metricus females. Insectes Sociaux 2009, 56, 77–84. [Google Scholar] [CrossRef]
- Judd, T.M.; Magnus, R.M.; Fasnacht, M.P. A nutritional profile of the social wasp Polistes metricus: Differences in nutrient levels between castes and changes within castes during the annual life cycle. J. Insect Physiol. 2010, 56, 42–56. [Google Scholar] [CrossRef]
- Hunt, J.H. Evolution of castes in Polistes. Ann. Zool. Fennici. 2006, 43, 407–422. [Google Scholar]
- Wilde, J.D.; Beetsma, J. The physiology of caste development in social insects. Adv. Insect Physiol. 1982, 16, 167–246. [Google Scholar] [CrossRef]
- Kamakura, M. Royalactin induces queen differentiation in honeybees. Nature 2011, 473, 478–483. [Google Scholar] [CrossRef]
- O’Donnell, S. Reproductive caste determination in eusocial wasps (Hymenoptera: Vespidae). Annu. Rev. Entomol. 1998, 43, 323–346. [Google Scholar] [CrossRef] [PubMed]
- Berens, A.J.; Hunt, J.H.; Toth, A.L. Nourishment level affects caste related gene expression in Polistes wasps. BMC Genom. 2015, 16, 235. [Google Scholar] [CrossRef]
- Judd, T.M. Effect of the presence of brood on the behavior and nutrient level of emerging individuals in field colonies of Polistes metricus. Insectes Sociaux 2018, 65, 171–182. [Google Scholar] [CrossRef]
- Hunt, J.H. Origin of an evolutionary novelty: The worker phenotype of eusocial wasps. Insectes Sociaux 2021, 68, 303–318. [Google Scholar] [CrossRef]
- Yoshimura, H.; Sasaki, K. Factor that affect reproductive states in female eusocial Hymenoptera. In Advances in Animal Science and Zoology; Jenkins, O.P., Ed.; Nova Science Publishers: New York, NY, USA, 2020; Volume 15, pp. 133–161. [Google Scholar]
- Yoshimura, H.; Yamada, Y.Y. Preimaginal caste-related bias in the paper wasp Polistes jokahamae is limited to the first brood. Insectes Sociaux 2021, 68, 133–143. [Google Scholar] [CrossRef]
- Hunt, J.H.; Amdam, G.V. Bivoltinism as an antecedent to eusociality in the paper wasp genus Polistes. Science 2005, 308, 264–267. [Google Scholar] [CrossRef]
- Jeanne, R.L.; Suryanarayanan, S. A new model for caste development in social wasps. Commun. Integr. Biol. 2011, 4, 373–377. [Google Scholar] [CrossRef]
- Judd, T.M.; Teal, P.E.A.; Hernandez, E.J.; Choudhury, T.; Hunt, J.H. Quantitative differences in nourishment affect caste-related physiology and development in the paper wasp Polistes metricus. PLoS ONE 2015, 10, e0116199. [Google Scholar] [CrossRef]
- Jandt, J.M.; Suryanarayanan, S.; Hermanson, J.C.; Jeanne, R.L.; Toth, A.L. Maternal and nourishment factors interact to influence offspring developmental trajectories in social wasps. Proc. R. Soc. B 2017, 284, 20170651. [Google Scholar] [CrossRef]
- Yoshimura, H.; Yamada, Y.Y. Caste-fate determination primarily occurs after adult emergence in a primitively eusocial paper wasp: Significance of the photoperiod during the adult stage. Sci. Nat. 2018, 105, 15. [Google Scholar] [CrossRef]
- Karsai, I.; Hunt, J.H. Food quantity affect traits of offspring in the paper wasp Polistes metricus (Hymenoptera: Vespidae). Environ. Entomol. 2002, 31, 99–106. [Google Scholar] [CrossRef]
- Rossi, A.M.; Hunt, J.H. Honey supplementation and its developmental consequences: Evidence for food limitation in a paper wasp, Polistes metricus. Ecol. Entomol. 1988, 13, 437–442. [Google Scholar] [CrossRef]
- Wheeler, D.E. Developmental and physiological determinants of caste in social Hymenoptera: Evolutionary implications. Am. Nat. 1986, 128, 13–34. [Google Scholar] [CrossRef]
- Matsuura, M.; Yamane, S. Biology of the Vespine Wasps; Springer: Berlin, Germany, 1990. [Google Scholar]
- Brian, M.V.; Brian, A.D. The wasp, Vespula sylvestris Scopoli: Feeding, foraging and colony development. Trans. R. Entomol. Soc. Lond. 1952, 103, 1–26. [Google Scholar] [CrossRef]
- Ishay, J. Caste determination by social wasps: Cell size and building behaviour. Anim. Behav. 1975, 23, 425–431. [Google Scholar] [CrossRef]
- Suryanarayanan, S.; Hantschel, A.E.; Torres, C.G.; Jeanne, R.L. Changes in the temporal pattern of antennal drumming behavior across the Polistes fuscatus colony cycle (Hymenoptera, Vespidae). Insectes Sociaux 2011, 58, 97–106. [Google Scholar] [CrossRef]
- Suryanarayanan, S.; Hermanson, J.C.; Jeanne, R.L. A mechanical signal biases caste development in a social wasp. Curr. Biol. 2011, 21, 231–235. [Google Scholar] [CrossRef]
- Solís, C.R.; Strassmann, J.E. Presence of brood affects caste differentiation in the social wasp, Polistes exclamans Viereck (Hymenoptera: Vespidae). Funct. Ecol. 1990, 4, 531–541. [Google Scholar] [CrossRef]
- Reeve, H.K.; Peters, J.M.; Nonacs, P.; Starks, P.T. Dispersal of first “workers” in social wasps: Causes and implications of an alternative reproductive strategy. Proc. Natl. Acad. Sci. USA 1998, 95, 13737–13742. [Google Scholar] [CrossRef]
- Tibbetts, E.A. Dispersal decisions and predispersal behavior in Polistes paper wasp ‘workers’. Behav. Ecol. Sociobiol. 2007, 61, 1877–1883. [Google Scholar] [CrossRef]
- Strassmann, J.E. Parasitoids, predators, and group size in the paper wasp, Polistes exclamans. Ecology 1981, 62, 1225–1233. [Google Scholar] [CrossRef]
- Furuichi, S. Field observation of predation on paper wasp nests by introduced terrestrial slugs. Insectes Sociaux 2014, 61, 95–96. [Google Scholar] [CrossRef]
- Kozyra, K.B.; Baraniak, E. Causes of mortality of Polistes nimpha colonies. Insectes Sociaux 2016, 63, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Pardi, L. Dominance order in Polistes wasp. Physiol. Zool. 1948, 21, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Strassmann, J.E.; Meyer, D.C. Gerontocracy in the social wasp, Polistes exclamans. Anim. Behav. 1983, 31, 431–438. [Google Scholar] [CrossRef]
- Reeve, H.K. Polistes. In The Social Biology of Wasps; Ross, K.G., Matthews, R.W., Eds.; Comstock Publicating Associates, A Division of Cornell University Press: London, UK, 1991; pp. 99–148. [Google Scholar]
- Yoshimura, H.; Yamada, J.; Yamada, Y.Y. The queen of the paper wasp Polistes jokahamae (Hymenoptera: Polistinae) is not aggressive but maintains her reproductive priority. Sociobiology 2019, 66, 166–178. [Google Scholar] [CrossRef]
- Jandt, J.M.; Tibbetts, E.A.; Toth, A.L. Polistes paper wasps: A model genus for the study of social dominance hierarchies. Insectes Sociaux 2014, 61, 11–27. [Google Scholar] [CrossRef]
- Gamboa, G.J.; Wacker, T.L.; Scope, J.A.; Cornell, T.J.; Shellman-Reeve, J. The mechanism of queen regulation of foraging by workers in paper wasps (Polistes fuscatus, Hymenoptera: Vespidae). Ethology 1990, 85, 335–343. [Google Scholar] [CrossRef]
- Miyano, S. Worker reproduction and related behavior in orphan colonies of a Japanese paper wasp, Polistes jadwigae (Hymenoptera, Vespidae). J. Ethol. 1991, 9, 135–146. [Google Scholar] [CrossRef]
- Mitra, A. Queen pheromone and monopoly of reproduction by the queen in the social wasp Ropalidia marginata. Proc. Natl. Acad. Sci. USA 2014, 80, 1025–1044. [Google Scholar] [CrossRef]
- Strassmann, J.E. Evolutionary implications of early male and satellite nest production in Polistes exclamans colony cycles. Behav. Ecol. Sociobiol. 1981, 8, 55–64. [Google Scholar] [CrossRef]
- Kasuya, E. Social behavior of early emerging males of a Japanese paper wasp, Polistes chinensis antennalis (Hymenoptera: Vespidae). Res. Popul. Ecol. 1983, 25, 143–149. [Google Scholar] [CrossRef]
- Suzuki, T. Mating and laying of female-producing eggs by orphaned workers of a paper wasp, Polistes snelleni (Hymenoptera: Vespidae). Ann. Entomol. Soc. Am. 1985, 78, 736–739. [Google Scholar] [CrossRef]
- Suzuki, T. Paradox of worker reproduction and worker mating in temperate paper wasps, Polistes chinensis and P. snelleni (Hymenoptera: Vespidae). Ethol. Ecol. Evol. 1998, 10, 347–359. [Google Scholar] [CrossRef]
- Yamasaki, K.; Takahashi, J.; Ono, M.; Tsuchida, K. Reproductivity of early males of the temperate paper wasp Polistes rothneyi iwatai. Entomol. Sci. 2011, 14, 383–386. [Google Scholar] [CrossRef]
- Kasuya, E. Internidal drifting of workers in the Japanese paper wasp Polistes chinensis antennalis (Vespidae; Hymenoptera). Insectes Sociaux 1981, 28, 343–346. [Google Scholar] [CrossRef]
- Sumner, S.; Lucas, E.; Barker, J.; Isaac, N. Radio-tagging technology reveals extreme nest-drifting behavior in a eusocial insect. Curr. Biol. 2007, 17, 140–145. [Google Scholar] [CrossRef]
- Nishimura, M.; Ono, M. Evidence of alternative reproduction by drifting workers in the Japanese paper wasp, Polistes rothneyi Cameron, 1900 (Hymenoptera: Vespidae). Entomol. Sci. 2021, 24, 111–115. [Google Scholar] [CrossRef]
- Bohm, M.K. Effects of environment and juvenile hormone on ovaries of the wasp, Polistes metricus. J. Insect Physiol. 1972, 18, 1875–1883. [Google Scholar] [CrossRef]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Zhuang, M.; Colgan, T.J.; Guo, Y.; Zhang, Z.; Liu, F.; Xia, Z.; Dai, X.; Zhang, Z.; Li, Y.; Wang, L.; et al. Unexpected worker mating and colony-founding in a superorganism. Nat. Commun. 2023, 14, 5499. [Google Scholar] [CrossRef]
- Röseler, P.-F.; Röseler, I. Caste specific differences in fat body glycogen metabolism of the bumblebee, Bombus terrestris. Insect Biochem. 1986, 16, 501–508. [Google Scholar] [CrossRef]
- Hunt, J.H.; Wolschin, F.; Henshaw, M.T.; Newman, T.C.; Toth, A.L.; Amdam, G.V. Differential gene expression and protein abundance evince ontogenetic bias toward castes in a primitively eusocial wasp. PLoS ONE 2010, 5, e10674. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.H.; Kensinger, B.J.; Kossuth, J.A.; Henshaw, M.T.; Norberg, K.; Wolschin, F.; Amdam, G.V. A diapause pathway underlies the gyne phenotype in Polistes wasps, revealing an evolutionary route to caste-containing insect societies. Proc. Natl. Acad. Sci. USA 2007, 104, 14020–14025. [Google Scholar] [CrossRef]
- Wheeler, D.E.; Buck, N.; Evans, J.D. Expression of insulin pathway genes during the period of caste determination in the honey bee, Apis mellifera. Insect Mol. Biol. 2006, 15, 597–602. [Google Scholar] [CrossRef]
- Wolschin, F.; Mutti, N.S.; Amdam, G.V. Insulin receptor substrate influences female caste development in honeybees. Biol. Lett. 2011, 7, 112–115. [Google Scholar] [CrossRef]
- Mutti, N.S.; Dolezal, A.G.; Wolschin, F.; Mutti, J.S.; Gill, K.S.; Amdam, G.V. IRS and TOR nutrient-signaling pathways act via juvenile hormone to influence honey bee caste fate. J. Exp. Biol. 2011, 214, 3977–3984. [Google Scholar] [CrossRef]
- Wheeler, D.E.; Buck, N.A.; Evans, J.D. Expression of insulin/insulin-like signalling and TOR pathway genes in honey bee caste determination. Insect Mol. Biol. 2014, 23, 113–121. [Google Scholar] [CrossRef]
- Patel, A.; Fondrk, M.K.; Kaftanoglu, O.; Emore, C.; Hunt, G.; Frederick, K.; Amdam, G.V. The making of a queen: TOR pathway is a key player in diphenic caste development. PLoS ONE 2007, 2, e509. [Google Scholar] [CrossRef] [PubMed]
- Hartfelder, K.; Guidugli-Lazzarini, K.R.; Cervoni, M.S.; Santos, D.E.; Humann, F.C. Old threads make new tapestry—Rewiring of signalling pathways underlies caste phenotypic plasticity in the honey bee, Apis mellifera L. Adv. Insect Physiol. 2015, 48, 1–36. [Google Scholar] [CrossRef]
- Corona, M.; Libbrecht, R.; Wheeler, D.E. Molecular mechanisms of phenotypic plasticity in social insects. Curr. Opin. Insect Sci. 2016, 13, 55–60. [Google Scholar] [CrossRef]
- Buttstedt, A.; Ihling, C.H.; Pietzsch, M.; Moritz, R.F.A. Royalactin is not a royal making of a queen. Nature 2016, 537, E10–E12. [Google Scholar] [CrossRef] [PubMed]
- Maleszka, R. Beyond royalactin and a master inducer explanation of phenotypic plasticity in honey bees. Commun. Biol. 2018, 1, 8. [Google Scholar] [CrossRef]
- Hartfelder, K.; Engels, W. Social insect polymorphism: Hormonal regulation of plasticity in development and reproduction in the honeybee. Curr. Top. Dev. Biol. 1998, 40, 45–77. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Yoshimura, H.; Yokoi, K. Brain physiology during photoperiod-related caste determination in the primitively eusocial wasp Polistes jokahamae. Sci. Rep. 2024, 14, 30399. [Google Scholar] [CrossRef]
- Formesyn, E.M.; Cardoen, D.; Ernst, U.R.; Danneels, E.L.; van Vaerenbergh, M.; de Koker, D.; Verleyen, P.; Wenseleers, T.; Schoofs, L.; de Graaf, D.C. Reproduction of honeybee workers is regulated by epidermal growth factor receptor signaling. Gen. Comp. Endocrinol. 2014, 197, 1–4. [Google Scholar] [CrossRef]
- Sasaki, K.; Ugajin, A.; Harano, K. Caste-specific development of the dopaminergic system during metamorphosis in female honey bees. PLoS ONE 2018, 13, e0206624. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Yokoi, K.; Toga, K. Bumble bee queens activate dopamine production and gene expression in nutritional signaling pathways in the brain. Sci. Rep. 2021, 11, 5526. [Google Scholar] [CrossRef]
- Harano, K.; Sasaki, M.; Nagao, T.; Sasaki, K. Dopamine influences locomotor activity in honeybee queens: Implications for a behavioural change after mating. Physiol. Entomol. 2008, 33, 395–399. [Google Scholar] [CrossRef]
- Sasaki, K.; Harada, M. Dopamine production in the brain is associated with caste-specific morphology and behavior in an artificial intermediate honey bee caste. PLoS ONE 2020, 15, e0244140. [Google Scholar] [CrossRef]
- Morigami, A.; Sasaki, K. Physiological specialization of the brain in bumble bee castes: Roles of dopamine in mating-related behaviors in female bumble bees. PLoS ONE 2024, 19, e0298682. [Google Scholar] [CrossRef]
- Sasaki, K.; Yoshimura, H.; Nishimura, M. Caste-specific storage of dopamine-related substances in the brains of four Polistes paper wasp species. PLoS ONE 2023, 18, e0280881. [Google Scholar] [CrossRef]
- Sasaki, K.; Yamasaki, K.; Tsuchida, K.; Nagao, T. Gonadotrophic effects of dopamine in isolated workers of the primitively eusocial wasp, Polistes chinensis. Naturwissenschaften 2009, 96, 625–629. [Google Scholar] [CrossRef]
- Toth, A.L.; Varala, K.; Newman, T.C.; Miguez, F.E.; Hutchison, S.K.; Willoughby, D.A.; Simons, J.F.; Egholm, M.; Hunt, J.H.; Hudson, M.E.; et al. Wasp gene expression supports an evolutionary link between material behavior and eusociality. Science 2007, 318, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Jedlička, P.; Ernst, U.R.; Votavová, A.; Hanus, R.; Valterová, I. Gene expression dynamics in major endocrine regulatory pathways along the transition from solitary to social life in a bumble bee, Bombus terrestris. Front. Physiol. 2016, 7, 574. [Google Scholar] [CrossRef]
- Chandra, V.; Fetter-Pruneda, I.; Oxley, P.R.; Ritger, A.L.; McKenzie, S.K.; Libbrecht, R.; Kronauer, D.J.C. Social regulation of insulin signaling and the evolution of eusociality in ants. Science 2018, 361, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Gruntenko, N.E.; Rauschenbach, I.Y. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A mini-review. Gen. Comp. Endocrinol. 2018, 258, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.H. The Evolution of Social Wasps; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Yoshimura, H.; Yamada, Y.Y. The first brood emerges smaller, lighter, and with lower lipid stores in the paper wasp Polistes jokahamae (Hymenoptera: Vespidae). Insectes Sociaux 2018, 65, 473–481. [Google Scholar] [CrossRef]
- Mead, F.; Habersetzer, C.; Gabouriaut, D.; Gervet, J. Dynamics of colony development in the paper wasp Polistes dominulus Christ (Hymenoptera, Vespidae): The influence of prey availability. J. Ethol. 1994, 12, 43–51. [Google Scholar] [CrossRef]
- Kudô, K.; Shirai, A. Effect of food availability on larval cannibalism by foundresses of the paper wasp Polistes chinensis antennalis. Insectes Sociaux 2012, 59, 279–284. [Google Scholar] [CrossRef]
- LeBoeuf, A.C.; Waridel, P.; Brent, C.S.; Gonçalves, A.N.; Menin, L.; Ortiz, D.; Riba-Grognuz, O.; Koto, A.; Soares, Z.G.; Privman, E.; et al. Oral transfer of chemical cues, growth proteins and hormones in social insects. eLife 2016, 5, e20375. [Google Scholar] [CrossRef]
- Jeanne, R.L. Evolution of social behavior in the Vespidae. Ann. Rev. Entomol. 1980, 25, 371–396. [Google Scholar] [CrossRef]
- Jone, J.C.; Oldroyd, B.P. Nest thermoregulation in social insects. Adv. Insect Physiol. 2007, 33, 153–191. [Google Scholar] [CrossRef]
- Kovac, H.; Nagy, J.M.; Käfer, H.; Stabentheiner, A. Relationship between nest and body temperature and microclimate in the paper wasp Polistes dominula. Insects 2023, 14, 886. [Google Scholar] [CrossRef] [PubMed]
- Gradišek, A.; Bizjak, J.; Popovski, A.; Grad, J. Bumble bee nest thermoregulation: A field study. J. Apic. Res. 2023, 62, 634–642. [Google Scholar] [CrossRef]
- Darrouzet, E.; Gévar, J.; Guignard, Q.; Aron, S. Production of early diploid males by European colonies of the invasive hornet Vespa velutina nigrithorax. PLoS ONE 2015, 10, e0136680. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshimura, H.; Sasaki, K. Plasticity in Caste-Fate Determination During the Adult Stage in Temperate Polistes Wasps. Insects 2025, 16, 326. https://doi.org/10.3390/insects16030326
Yoshimura H, Sasaki K. Plasticity in Caste-Fate Determination During the Adult Stage in Temperate Polistes Wasps. Insects. 2025; 16(3):326. https://doi.org/10.3390/insects16030326
Chicago/Turabian StyleYoshimura, Hideto, and Ken Sasaki. 2025. "Plasticity in Caste-Fate Determination During the Adult Stage in Temperate Polistes Wasps" Insects 16, no. 3: 326. https://doi.org/10.3390/insects16030326
APA StyleYoshimura, H., & Sasaki, K. (2025). Plasticity in Caste-Fate Determination During the Adult Stage in Temperate Polistes Wasps. Insects, 16(3), 326. https://doi.org/10.3390/insects16030326