

Subfossil Insects of the Kebezen Site (Altai Mountains): New Data on the Last Deglaciation Environment

 , , , ,
, , , ,  , ,
, , 
Simple Summary
Abstract
1. Introduction
2. Regional Setting
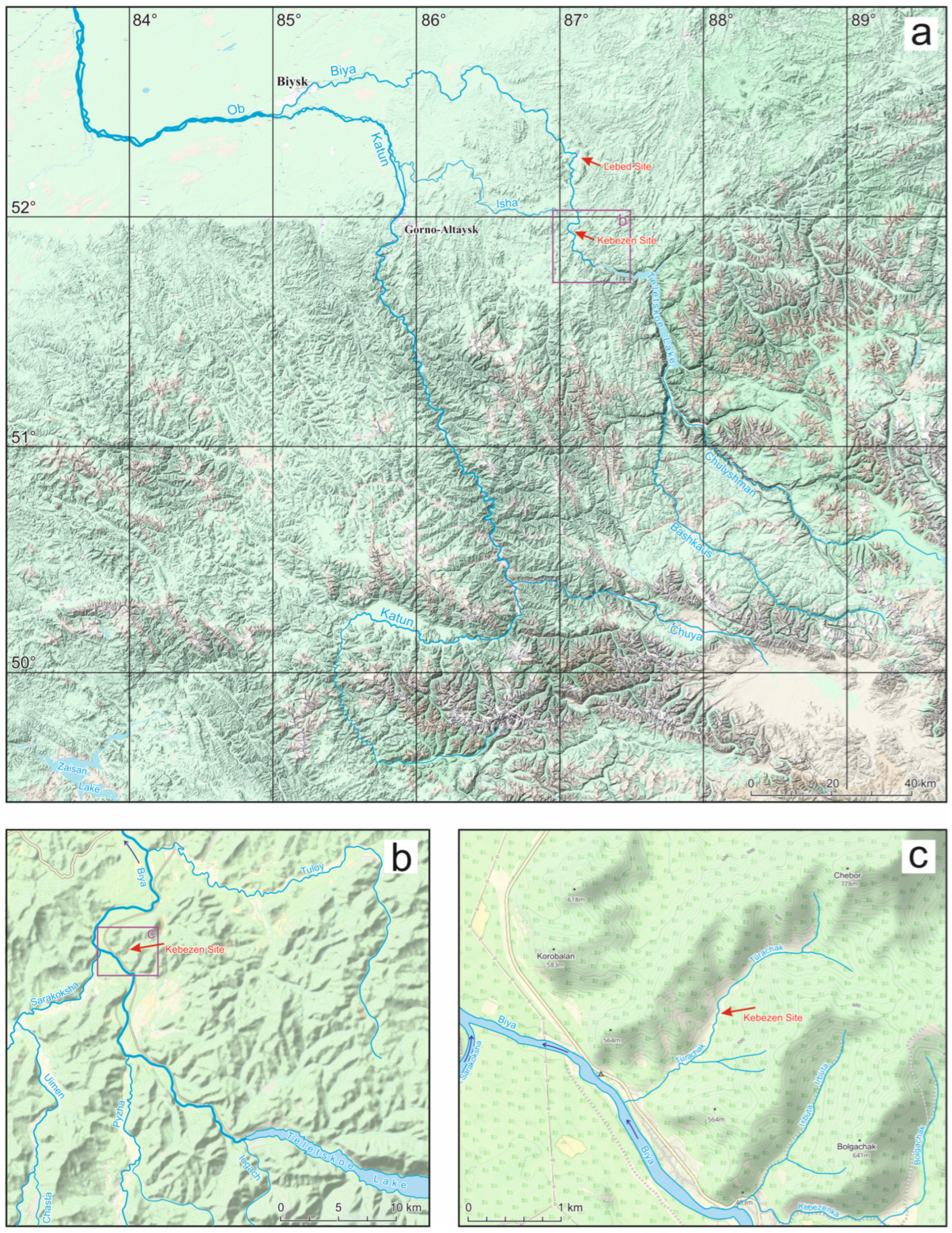
2.1. Study Area
2.2. Geological Setting
2.3. Description of the Sections
3. Materials and Methods
3.1. Sampling
3.2. Treatment
3.3. Radiocarbon Dating
4. Results
4.1. Entomological Data
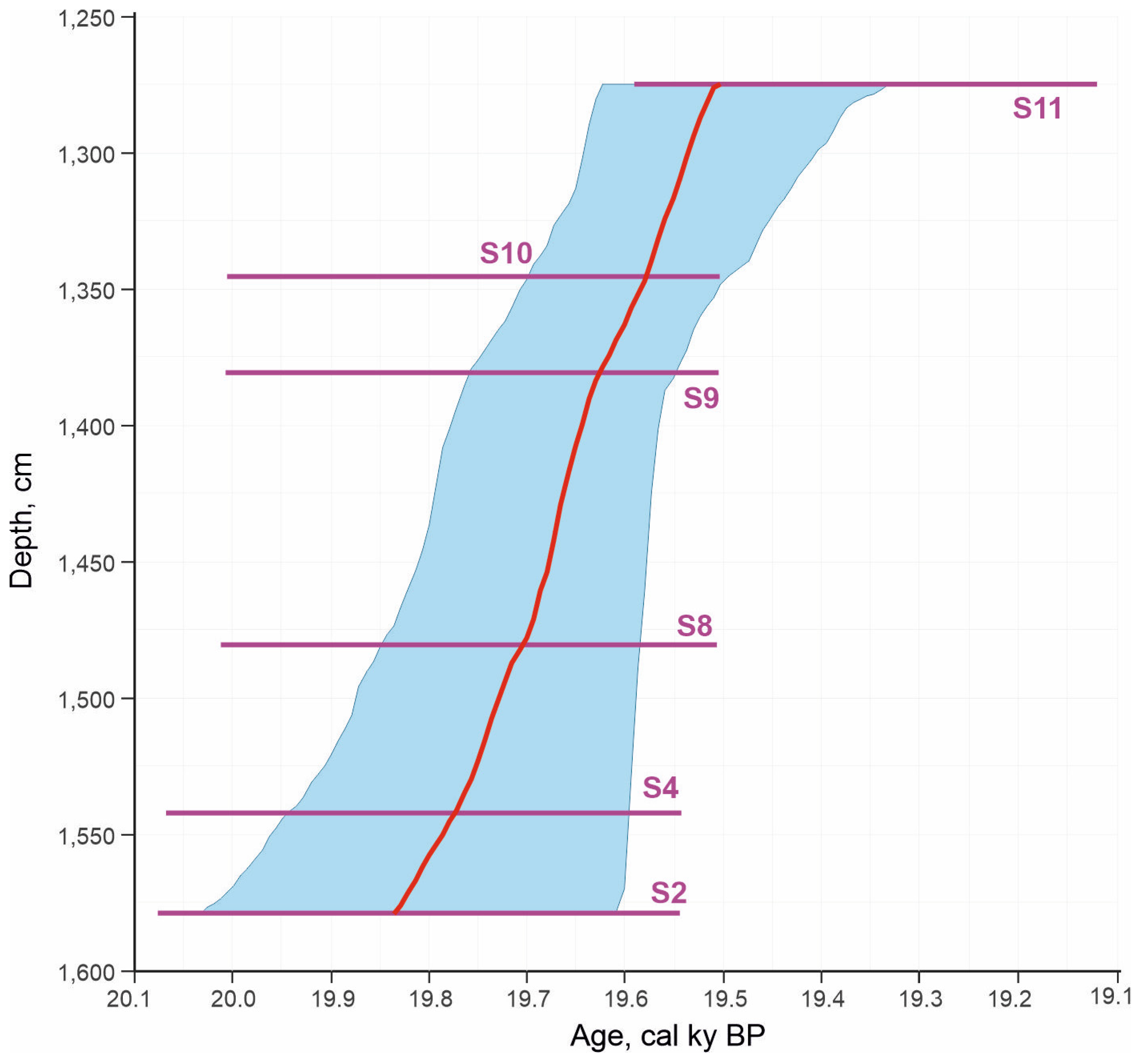
4.2. Radiocarbon Dates
5. Discussion
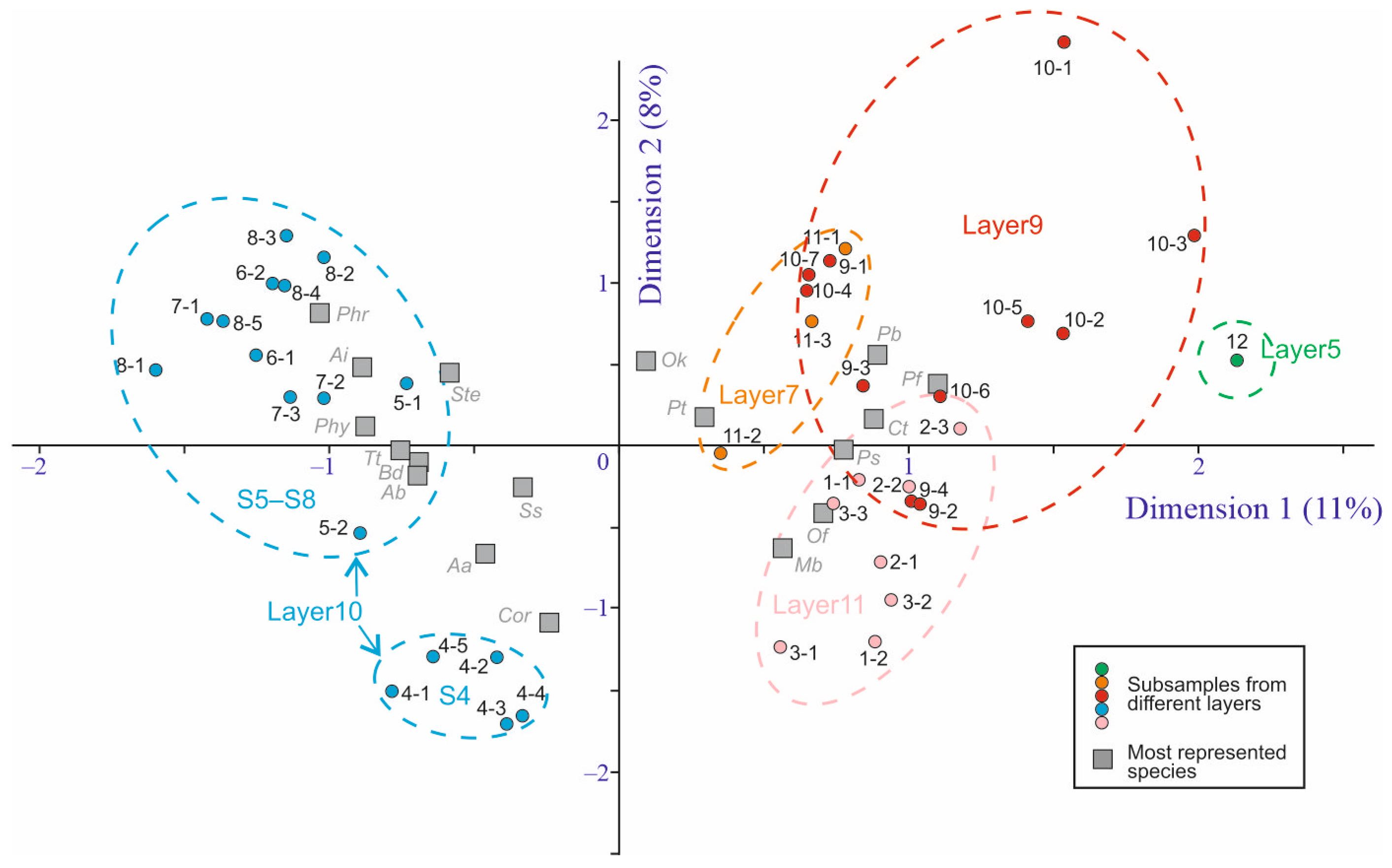
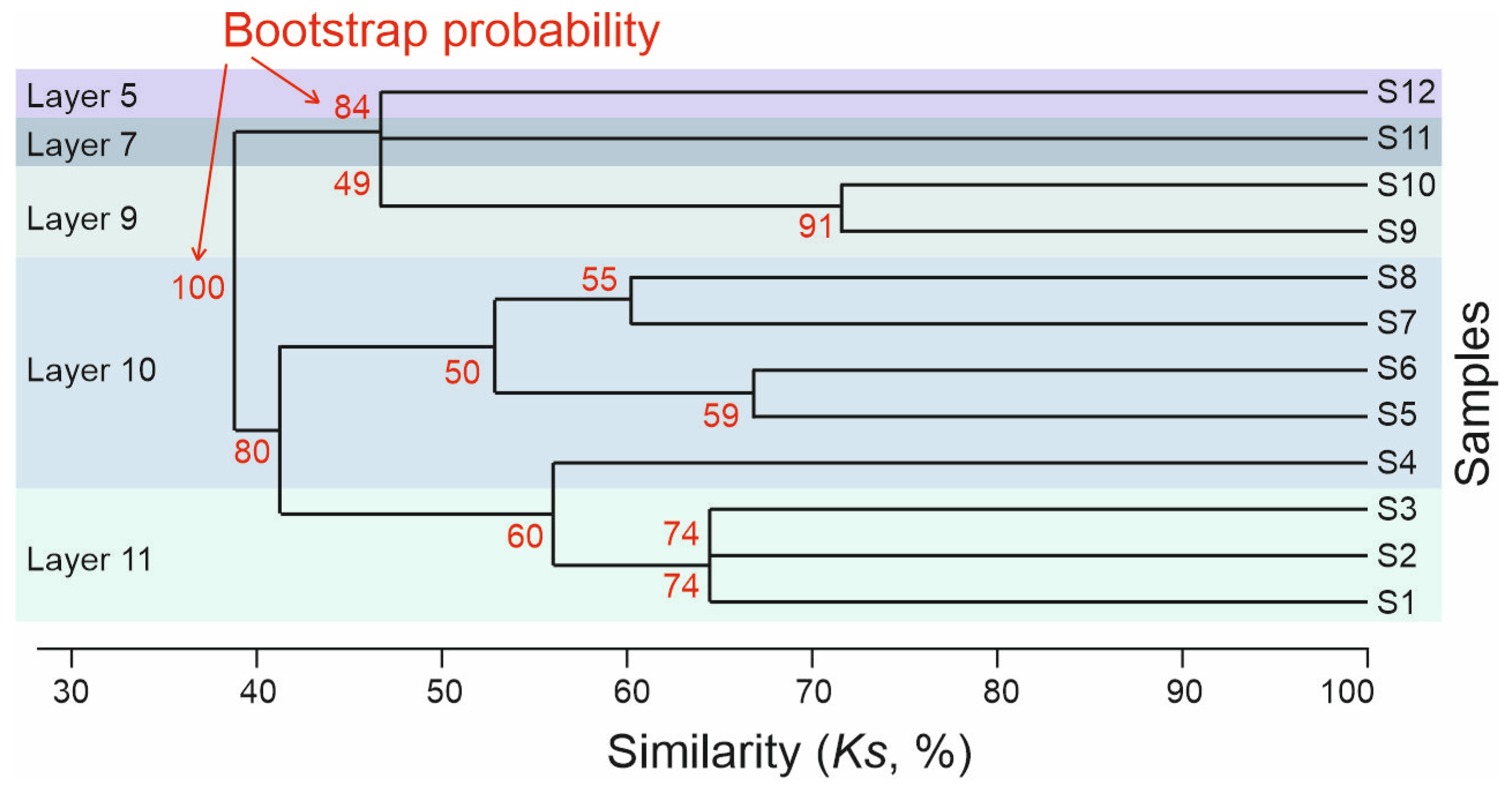
5.1. Taxonomical Composition and Diversity
5.2. Faunal Comparisons
5.2.1. Comparison with Late Pleistocene fauna of Western Siberia
5.2.2. Comparison with Modern Beetle Fauna
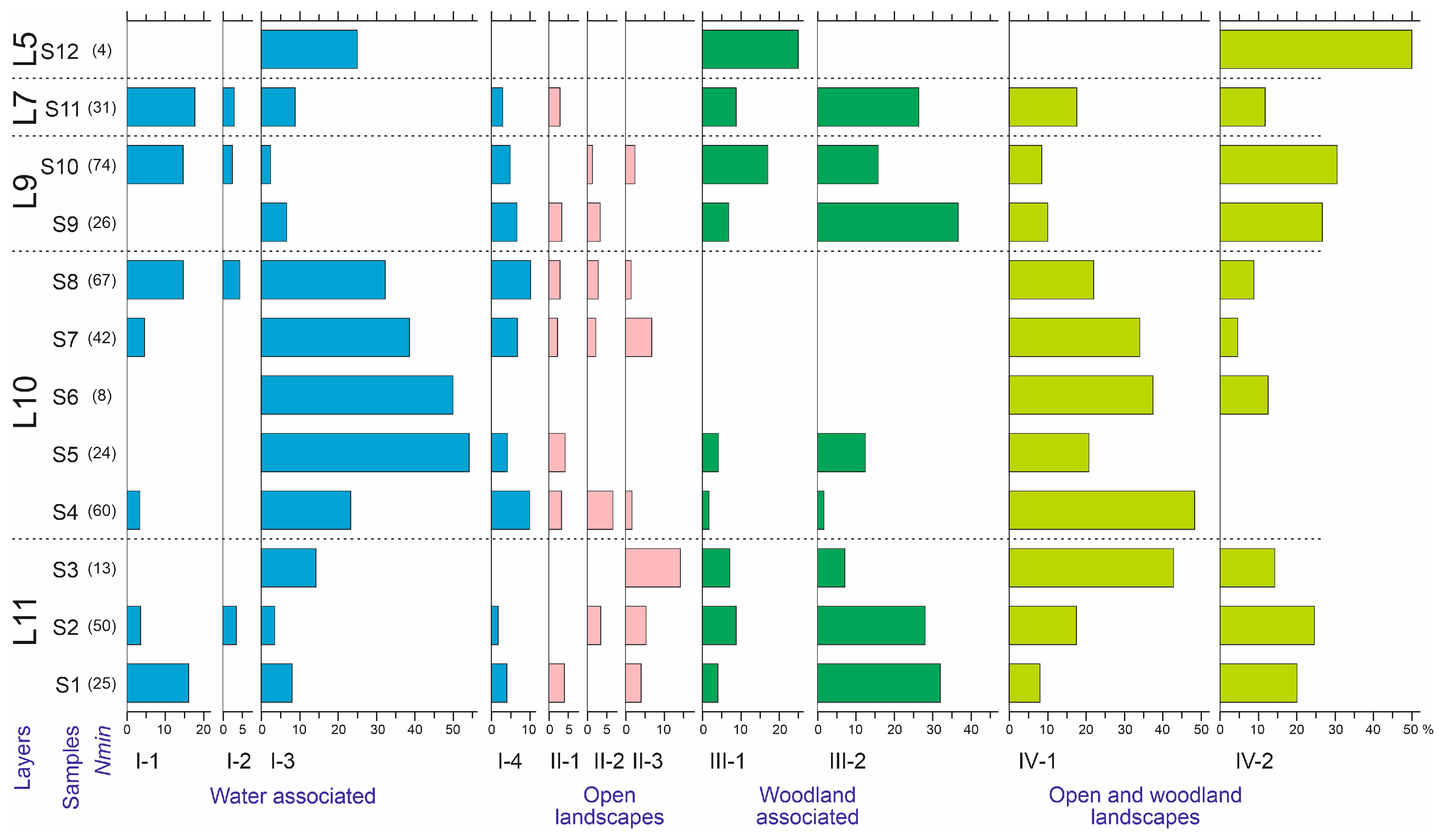
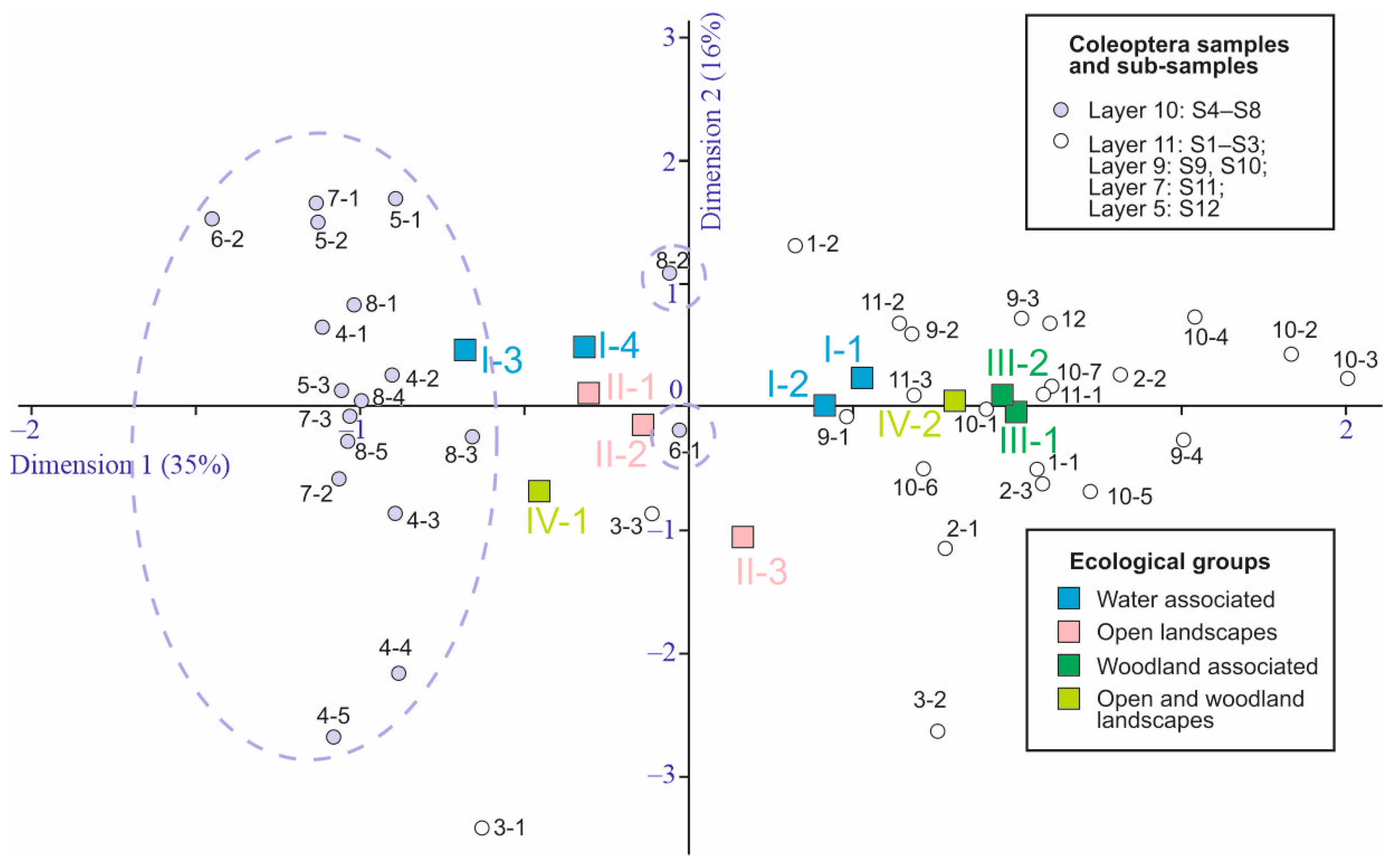
5.3. Ecological Analysis
5.3.1. Altitudinal Distribution
5.3.2. Trophic Groups
- Species developing on conifers, predominantly Picea, less frequently Pinus: Pissodes gyllenhali, Xylechinus pilosus, Carphoborus teplouchovi, Polygraphus, and P. subopacus. No species obligate to other species of coniferous and deciduous trees were identified.
- Species associated with near-water herbaceous vegetation (Phaedon armoraciae, Tournotaris bimaculata, Notaris acridulus, Ceutorhynchus cochleariae) and shrubby vegetation (Lochmaea caprea, Phratora sp., Gonioctena spp., Dorytomus taeniatus). These species indicate the presence of willow (possibly Salix aurita, S. dasyclados, S. myrsinifolia, S. caprea), Brassicaceae (Cardamine sp.), Ranunculaceae (Caltha sp.), Plantaginaceae (Veronica sp.), and Poaceae (Glyceria sp., Phalaris arundinacea sp.) in the area.
- Species associated with meadow vegetation (Sternoplatys fulvipes, Hemitrichapion tschernovi, Eutrichapion rhomboidale, Sitona lineellus, Boreohypera diversipunctata, Phyllobius pomaceus, Trichalophus maklini, and Otiorhynchus grandineus). These species indicate the presence of Fabaceae (Lathyrus gmelini, Oxytropis sp., Vicia sp., Trifolium sp., Medicago sp.), Caryophyllaceae (Cerastium sp., Stellaria sp., Silene sp.), Grossulariaceae (Ribes sp.), Urticaceae (Urtica sp.), and Rosaceae (Filipendula sp.) in the area.
- Species trophically associated with algae or moist detritus, such as Helophorus praenanus, H. sibiricus, Ochtebius flavipes, and O. kaninensis.
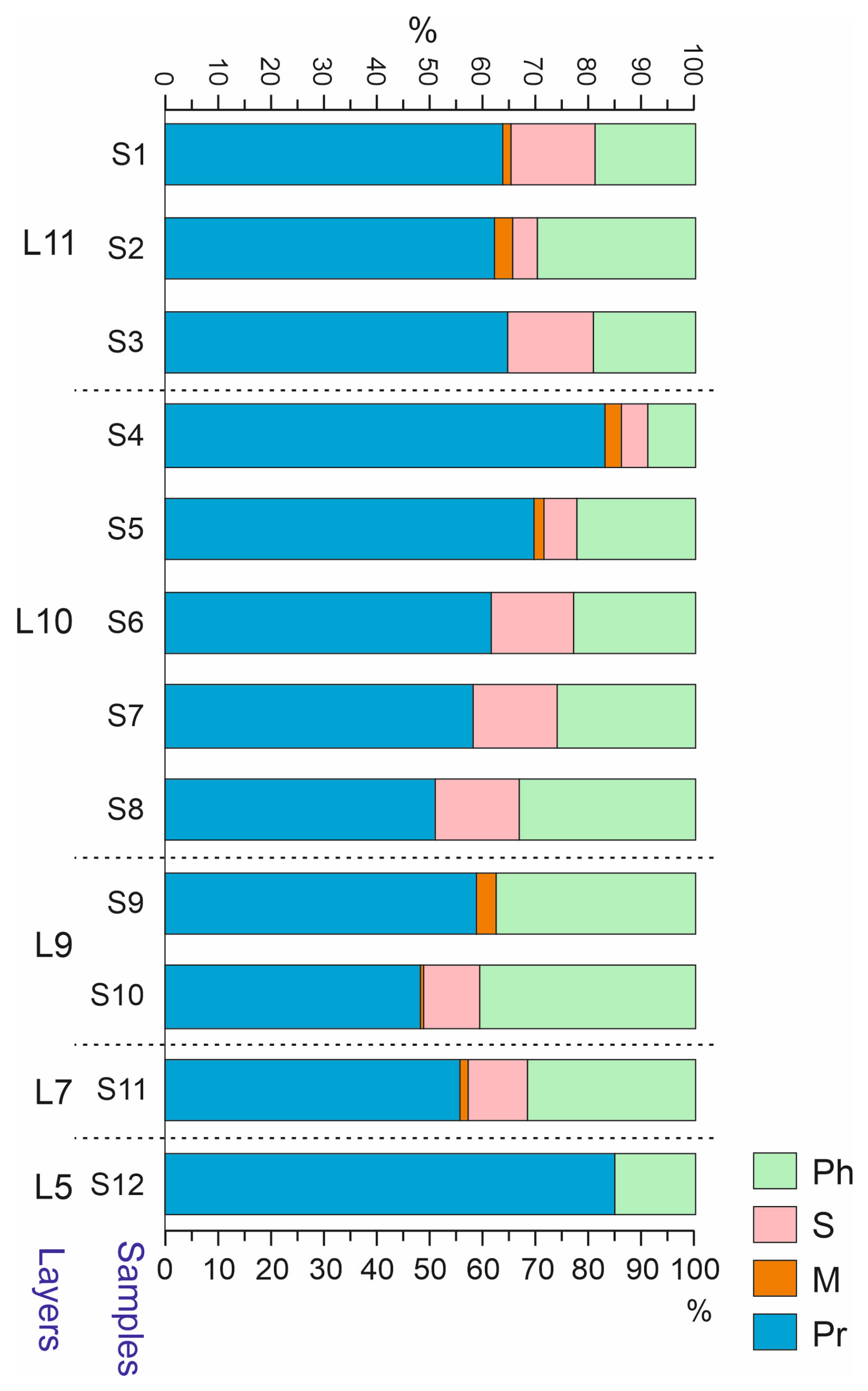
5.3.3. Biotopic Groups
5.4. Conditions for the Formation of the Kebezen Section
5.4.1. The Lower Strata of Lacustrine and Floodplain Sediments (Layers 11, 10)
5.4.2. The Middle Strata (Layers 9–7)
5.4.3. The Upper Strata (Layers 5 and 4)
5.5. Environment Reconstruction and Age Correlation
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BP | before present |
| cal | calibrated |
| GLOF | glacial lake outburst floods |
| ky | thousand years |
| MIS | marine isotope stage |
| Nmin | minimum number of individuals |
| UPGMA | unweighted pair group method with arithmetic mean |
| V | volume |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COLEOPTERA | |||||||||||||
| Dytiscidae | |||||||||||||
| 1 | Hydroporus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Carabidae | |||||||||||||
| 2 | Notiophilus fasciatus Mäklin, 1855 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 1 | 0 |
| 3 | Nebria gyllenhali (Schönherr, 1806) | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4 | Carabus (Carabus) arvensis Herbst, 1784 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 |
| 5 | C. (Morphocarabus) cf. aeruginosus Fischer von Waldheim, 1820 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 0 |
| 6 | *C. (Carabulus) obovatus Fischer von Waldheim, 1828 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| – | Carabus sp. | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 7 | Loricera pilicornis (Fabricius, 1775) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 8 | Diacheila polita (Faldermann, 1835) | 1 | 2 | 2 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 |
| 9 | Elaphrus (Elaphrus) cf. riparius (Linnaeus, 1758) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 10 | E. (Elaphroterus) angusticollis R.F. Sahlberg, 1844 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 11 | *Dyschiriodes nigricornis (Motschulsky, 1844) | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 12 | *Trechus cf. lomakini Belousov et Kabak, 1994 | 0 | 1 | 0 | 2 | 0 | 0 | 1 | 2 | 1 | 1 | 0 | 0 |
| 13 | *T. cf. toroticus Belousov et Kabak, 1996 | 1 | 1 | 0 | 9 | 0 | 0 | 8 | 10 | 0 | 1 | 0 | 0 |
| – | Trechus spp. | 1 | 3 | 1 | 4 | 2 | 1 | 2 | 2 | 0 | 0 | 0 | 0 |
| 14 | *Bembidion (Testedium) bipunctatum (Linnaeus, 1760) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 0 |
| 15 | B. (Notaphus) sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 16 | B. (Plataphus) difficile (Motschulsky, 1844) | 2 | 1 | 2 | 11 | 11 | 2 | 6 | 10 | 0 | 0 | 1 | 0 |
| 17 | *B. (Asioperyphus) sajanum Shilenkov, 1995 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 18 | B. (Peryphus) dauricum (Motschulsky, 1844) | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 1 | 0 | 0 |
| – | Bembidion sp. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 19 | Pterostichus (Cryobius) cf. brevicornis (Kirby, 1837) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 20 | *P. (Cryobius) cf. burjaticus (Poppius, 1906) | 4 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 8 | 0 | 0 |
| 21 | *P. (C.) cf. fulvescens (Motschulsky, 1844) | 1 | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 8 | 1 | 1 |
| – | P. (Cryobius) spp. | 0 | 8 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 8 | 3 | 0 |
| 22 | P. (Eosteropus) maurusiacus (Mannerheim, 1825) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| 23 | *P. (Petrophilus) cf. ehnbergi Poppius, 1908 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 24 | P. (P.) monticoloides Shilenkov, 1995/P. subaeneus (Chaudoir, 1850) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 25 | *P. (P.) triseriatus (Gebler, 1847) | 1 | 2 | 4 | 1 | 1 | 1 | 2 | 2 | 3 | 3 | 2 | 0 |
| – | P. (Petrophilus) sp. | 1 | 6 | 0 | 2 | 0 | 0 | 0 | 2 | 2 | 2 | 2 | 0 |
| – | Pterostichus spp. | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 |
| 26 | Amara sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 27 | Agonum alpinum Motschulsky, 1844 | 0 | 3 | 0 | 7 | 3 | 1 | 1 | 2 | 0 | 0 | 0 | 0 |
| – | Agonum sp. | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 28 | Harpalus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| – | Carabidae indet. | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 2 | 1 | 4 | 1 | 0 |
| Helophoridae | |||||||||||||
| 29 | Helophorus (Gephelophorus) sibiricus (Motschulsky, 1960) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 30 | *H. (Rhopalohelophorus) praenanus Łomnicki, 1894 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Hydrophilidae | |||||||||||||
| 31 | Hydrophilidae indet. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| Hydraenidae | |||||||||||||
| 32 | *Ochtebius (Asiobates) cf. flavipes Dalla Torre, 1877 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 4 | 1 | 0 |
| 33 | O. (A.) kaninensis Poppius, 1909 | 2 | 2 | 0 | 2 | 0 | 0 | 2 | 5 | 0 | 8 | 1 | 0 |
| Leiodidae | |||||||||||||
| 34 | *Catops cf. alpinus Gyllenhal, 1827 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 35 | *Colon cf. bidentatum (C.R. Sahlberg, 1822) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| Staphylinidae | |||||||||||||
| 36 | *Arpedium cf. brachypterum (Gravenhorst, 1802) | 1 | 0 | 1 | 6 | 0 | 0 | 6 | 6 | 0 | 0 | 2 | 0 |
| 37 | *Mannerheimia cf. brevipennis (Motschulsky, 1860) | 2 | 3 | 3 | 5 | 1 | 0 | 0 | 0 | 2 | 2 | 0 | 1 |
| 38 | Olophrum cf. fuscum (Gravenhorst, 1806) | 14 | 8 | 3 | 5 | 0 | 0 | 1 | 0 | 0 | 1 | 6 | 2 |
| 39 | Olophrum sp.1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 40 | Coryphium sp.1 | 0 | 2 | 0 | 12 | 1 | 0 | 2 | 0 | 0 | 1 | 0 | 0 |
| 41 | Omalium sp.1 | 0 | 0 | 0 | 2 | 1 | 0 | 4 | 1 | 0 | 0 | 0 | 0 |
| 42 | Phyllodrepa sp.1 | 1 | 0 | 0 | 3 | 3 | 0 | 0 | 8 | 0 | 0 | 0 | 0 |
| 43 | Omaliinae indet. sp.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 44 | Omaliinae indet. sp.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 45 | Omaliinae indet. sp.3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| – | Omaliinae indet. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 2 |
| 46 | Mycetoporus sp.1 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 47 | Mycetoporus sp.2 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 48 | Tachyporus sp.1 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 49 | Aleocharinae indet. sp.1 | 1 | 0 | 0 | 12 | 3 | 1 | 0 | 2 | 8 | 4 | 1 | 0 |
| 50 | Aleocharinae indet. sp.2 | 3 | 1 | 1 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 51 | Aleocharinae indet. sp.3 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 52 | Aleocharinae indet. sp.4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| – | Aleocharinae indet. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 |
| 53 | Bledius sp.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 2 |
| 54 | Stenus sp.1 | 1 | 0 | 0 | 2 | 1 | 0 | 3 | 7 | 1 | 1 | 1 | 0 |
| 55 | Stenus sp.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 56 | Rugilus sp.1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 0 |
| 57 | Quedius sp.1 | 0 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 58 | Quedius sp.2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 0 |
| 59 | Staphylininae sp.1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Staphylinidae indet. | 2 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | |
| Scarabaeidae | |||||||||||||
| 60 | Psammoporus cf. matalini (Gusakov, 2003) | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 61 | *Aphodius ater (De Geer, 1774) | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 62 | Aphodius sp.1 | 2 | 0 | 2 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| 63 | Aphodius sp.2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 64 | Aphodius sp.3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 65 | Aphodius sp.4 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| – | Aphodius spp. | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| 66 | Onthophagus sp. | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Byrrhidae | |||||||||||||
| 67 | Simplocaria semistriata (Fabricius, 1794) | 0 | 1 | 0 | 4 | 0 | 0 | 1 | 2 | 0 | 3 | 0 | 0 |
| 68 | Byrrhus cf. mordkovitshi Tshernyshev et Dudko, 1997 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 |
| Heteroceridae | |||||||||||||
| 69 | *Augyles intermedius (Kiesenwetter, 1843) | 0 | 1 | 0 | 1 | 2 | 1 | 10 | 8 | 0 | 1 | 2 | 0 |
| Elateridae | |||||||||||||
| 70 | Hypnoidus rivularius-group | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 2 | 1 | 0 | 0 |
| 71 | *Denticollis acuticollis (Motschulsky, 1860) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cantharidae | |||||||||||||
| 72 | Podabrus alpinus (Paykull, 1798)/P. annulatus (Mannerheim, 1825) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 73 | *Rhagonycha cf. nigriventris Motschulsky, 1860 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Dermestidae | |||||||||||||
| 74 | *Megatoma cf. undata (Linnaeus, 1758) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Laemophloeidae | |||||||||||||
| 75 | Leptophloeus alternans (Erichson, 1846) | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 |
| Coccinelidae | |||||||||||||
| 76 | Coccinella nivicola Mulsant, 1850 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ciidae | |||||||||||||
| 77 | *Sulcacis nitidus (Fabricius, 1792) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Pythidae | |||||||||||||
| 78 | *Pytho depressus (Linnaeus, 1767) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Chrysomelidae | |||||||||||||
| 79 | *Sternoplatys cf. fulvipes Motschulsky, 1860 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| 80 | Phratora sp. | 0 | 0 | 0 | 0 | 2 | 2 | 3 | 8 | 0 | 0 | 1 | 0 |
| 81 | *Phaedon cf. armoraciae (Linnaeus, 1758) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 0 | 0 |
| 82 | Gonioctena sp.1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 83 | Gonioctena sp.2 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 2 | 0 | 0 |
| 84 | *Lochmaea caprea (Linnaeus, 1758) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| 85 | Chaetocnema sp. | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4 | 0 | 1 | 0 | 0 |
| – | Alticinae indet. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 |
| – | Chrysomelidae indet. | 2 | 4 | 3 | 1 | 3 | 0 | 9 | 18 | 3 | 16 | 2 | 1 |
| Brentidae | |||||||||||||
| 86 | Eutrichapion rhomboidale (Desbrochers des Loges, 1871) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 87 | Hemitrichapion tschernovi (Ter-Minasian, 1973) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 88 | *Holotrichapion aethiops (Herbst, 1797) | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 5 | 0 | 0 |
| Curculionidae | |||||||||||||
| 89 | *Notaris acridulus (Linnaeus, 1758) | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 90 | Notaris sp. | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 91 | Tournotaris bimaculata (Fabricius, 1787) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 |
| 92 | *Dorytomus taeniatus (Fabricius, 1781) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| 93 | Pissodes gyllenhali (C.R. Sahlberg, 1834) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| 94 | *Ceutorhynchus cochleariae (Gyllenhal, 1813) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 |
| 95 | Tachyerges stigma (Germar, 1821)/T. pseudostigma (Tempere, 1982) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 96 | Trichalophus maklini (Faust, 1890) | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| 97 | Boreohypera diversipunctata (Schrank, 1798) | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 98 | Phyllobius pomaceus Gyllenhal, 1834 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| 99 | Otiorhynchus grandineus Germar, 1823 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 |
| 100 | Sitona lineellus (Bonsdorff, 1785) | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 | 1 | 0 | 1 | 0 |
| – | Curculionidae indet. | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 2 | 3 | 1 | 0 |
| Scolytidae | |||||||||||||
| 101 | *Xylechinus pilosus (Ratzeburg, 1837) | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | 1 | 0 |
| 102 | Phloeotribus spinulosus (Rey, 1883) | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 0 |
| 103 | Carphoborus teplouchovi Spessivtsev, 1916 | 5 | 5 | 1 | 0 | 1 | 0 | 0 | 0 | 4 | 5 | 5 | 0 |
| 104 | Polygraphus poligraphus (Linnaeus, 1758) | 1 | 3 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
| 105 | P. subopacus Thomson, 1871 | 1 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 0 |
| – | Scolytidae indet. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| – | Coleoptera indet. | 0 | 4 | 4 | 2 | 3 | 2 | 0 | 4 | 8 | 7 | 6 | 1 |
| HEMIPTERA | |||||||||||||
| – | Heteroptera indet. | 0 | 0 | 0 | 1 | 0 | 0 | 3 | 1 | 0 | 0 | 0 | 0 |
| – | Homoptera indet. | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 3 | 0 | 0 | 0 | 0 |
| HYMENOPTERA | |||||||||||||
| – | Hymenoptera indet. | 2 | 0 | 0 | 5 | 4 | 0 | 5 | 2 | 4 | 12 | 6 | 1 |
| DIPTERA | |||||||||||||
| – | Diptera indet. | 3 | 3 | 9 | 18 | 7 | 0 | 13 | 1 | 1 | 5 | 12 | 0 |
| INSECTA | |||||||||||||
| – | Insecta indet. | 0 | 0 | 2 | 11 | 0 | 0 | 10 | 4 | 0 | 6 | 3 | 0 |
| ARANEI | |||||||||||||
| – | Linyphiidae indet. | 0 | 0 | 0 | 0 | 4 | 1 | 11 | 6 | 1 | 1 | 2 | 0 |
| In total: | 64 | 76 | 39 | 151 | 58 | 14 | 91 | 154 | 62 | 147 | 77 | 15 |
References
- Barash, M.S.; Baryshnikov, G.F.; Baulin, V.V.; Becker, B.; Belyaev, A.V.; Beug, H.-J.; Borisova, O.K.; Borzenkova, I.I.; Brunnacker, K.; Busche, D.; et al. Atlas of Paleoclimates and Paleoenvironments of the Northern Hemisphere, Late Pleistocene—Holocene; Geographical Research Institute, Hungarian Academy of Sciences: Budapest, Hungary; Gustav Fischer Verlag: Stuttgart, Germany, 1992; pp. 1–153. [Google Scholar]
- Okishev, P.A. Relief i Oledenenie Russkogo Altaya [Relief and Glaciation of Russian Altai]; Tomsk University Publishing House: Tomsk, Russia, 2011; pp. 1–381. [Google Scholar]
- Hais, M.; Komprdová, K.; Ermakov, N.; Chytrý, M. Modelling the Last Glacial Maximum environments for a refugium of Pleistocene biota in the Russian Altai Mountains, Siberia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 438, 135–145. [Google Scholar] [CrossRef]
- Rusanov, G.G. Ozera i Paleogeografia Severnogo Altaya v Pozdnem Neopleistotsene i Golotsene [Lakes and Paleogeography of Northern Altai in the Late Neopleistocene and Holocene]; Biysk Pedagogical State University: Biisk, Russia, 2007; pp. 1–164. [Google Scholar]
- Baryshnikov, G.Y. Razvitie Reliefa Perekhodnykh zon Gornykh Stran v Kainozoe (na Primere Gornogo Altaya) [Development of the Relief of Transition Zones of Mountainous Countries in the Cenozoic (Using the Example of the Altai Mountains)]; Tomsk University Publishing House: Tomsk, Russia, 1992; pp. 1–182. [Google Scholar]
- Butvilovsky, V.V. Paleogeografia Poslednego Oledenenia i Golotsena Altaya: Sobytiino-Katastroficheskaya Model’ [Paleogeography of the Last Glaciation and the Holocene of Altai: An Event-Catastrophic Model]; Tomsk University Publishing House: Tomsk, Russia, 1993; pp. 1–252. [Google Scholar]
- Coope, G.R. Interpretations of Quaternary insect fossils. Annu. Rev. Entomol. 1970, 15, 97–120. [Google Scholar] [CrossRef]
- Coope, G.R. Fossil coleopteran assemblages as sensitive indicators of climatic changes during the Devensian (last) cold stage. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 280, 313–337. [Google Scholar]
- Zinovyev, E.V. Sub-fossil beetle assemblages associated with the «mammoth fauna» in the late Pleistocene localities of the Ural Mountains and West Siberia. ZooKeys 2011, 100, 149–169. [Google Scholar] [CrossRef]
- Zinovyev, E.V. Insect Fauna of the Ural and West Siberian Plain in the Quaternary Period. Ph.D. Thesis, Institute of Plant and Animal Ecology, UB RAS, Ekaterinburg, Russia, 2020. [Google Scholar]
- Gurina, A.A.; Dudko, R.Y.; Prosvirov, A.S.; Tshernyshev, S.E.; Legalov, A.A.; Zinovyev, E.V. Coleoptera assemblages from the Quaternary deposits of Kizikha river, the southernmost late Pleistocene insects of the West Siberian Plain. Invertebr. Zool. 2019, 16, 165–182. [Google Scholar] [CrossRef]
- Gurina, A.A.; Dudko, R.Y.; Ivanov, A.V.; Kotov, A.A.; Mikhailov, Y.E.; Prokin, A.A.; Prosvirov, A.S.; Solodovnikov, A.Y.; Zinovyev, E.V.; Legalov, A.A. New Data on the Distribution of Southern Forests for the West Siberian Plain during the Late Pleistocene: A Paleoentomological Approach. Diversity 2023, 15, 56. [Google Scholar] [CrossRef]
- Angus, R.B.; Ryndevich, S.K.; Zhang, T. A new species of Helophorus Fabricius, 1775 from the Chinese Altai, with notes on the former subgenus Atractohelophorus Kuwert, 1886 and selected species (Coleoptera: Helophoridae). Koleopterol. Rundsch 2017, 87, 239–252. [Google Scholar]
- Zhang, T.; Elias, S. Holocene palaeoenvironmental reconstruction based on fossil beetle faunas from the Southern Altai region, north-west China. J. Quat. Sci. 2019, 34, 593–602. [Google Scholar] [CrossRef]
- Gurina, A.A.; Dudko, R.Y.; Mikhailov, Y.E.; Prokin, A.A.; Solodovnikov, A.Y.; Zinovyev, E.V.; Legalov, A.A. First record of insects from the oldest and older Dryas of Altai (Russia). Coleoptera assemblages from Lebed River. Palaeoentomology 2024, 7, 112–131. [Google Scholar] [CrossRef]
- Kuminova, A.V. Rastitelnyi Pokrov Altaya [The Vegetation of the Altai]; SO AN SSSR: Novosibirsk, Russia, 1960; pp. 1–449. [Google Scholar]
- Ogureeva, N.G. Botanicheskaya Geografia Altaya [Botanical Geography of Altai]; Nauka: Moscow, Russia, 1980; pp. 1–180. [Google Scholar]
- Climate Data. Available online: https://en.climate-data.org/asia/russian-federation/altai-republic/iogach-443162/ (accessed on 1 February 2025).
- Rudoy, A.N. Gigantskaya Ryab’ Techeniya (Istoria Issledovanii, Diagnostika, Paleogeografichesloe Znachenie) [Giant Current Ripples (History of Research, Diagnostics, Paleogeographic Significance)]; Tomsk Pedagogical State University Publishing House: Tomsk, Russia, 2005; pp. 1–224. [Google Scholar]
- Baker, V.R.; Benito, G.; Rudoy, A.N. Paleohidrology of Late Pleistocene superflooding, Altai, Mountains, Siberia. Science 1993, 259, 348–350. [Google Scholar] [CrossRef]
- Bohorquez, P.; Jimenez-Ruiz, P.J.; Carling, P.A. Revisiting the dynamics of catastrophic late Pleistocene glacial-lake drainage, Altai Mountains, central Asia. Earth-Sci. Rev. 2019, 197, 102892. [Google Scholar] [CrossRef]
- Borisov, B.A.; Minina, E.A. Ribbed and mesh-cellular basic moraines of the Eastern Pamirs and the Altai Mountains. Geomorfologia 1979, 10, 69–74. [Google Scholar]
- Zykin, V.S.; Zykina, V.S. Discussion problems of Pleistocene paleogeography of glacial regions of Western Siberia. Adv. Curr. Nat. Sci. 2018, 7, 121–128. [Google Scholar] [CrossRef]
- Zykin, V.S.; Zykina, V.S.; Volvakh, N.E. Conditions of the Pleistocene Inya series formation Yaloman-Katun area of the Altai Mountains. Adv. Curr. Nat. Sci. 2018, 8, 118–129. [Google Scholar] [CrossRef]
- Pozdnyakov, A.V.; Pupyshev, Y.S. Self-organization and disorganization of ultra-high-pressure ice-dam basin (Chuya-Kurai natural phenomenon, Gornyi Altai). Geogr. I Prir. Resur. 2022, 4, 103–111. [Google Scholar] [CrossRef]
- Baryshnikov, G.Y.; Panychev, V.A. Features of the formation of terrace complexes of the upper Biya. Vopr. Geogr. Sib. 1987, 17, 41–52. [Google Scholar]
- Rusanov, G.G.; Orlova, L.A. Radiouglerodnye Datirovki (SOAN) Gornogo Altaya i Predaltaiskoi Ravniny: Katalog [Radiocarbon Dating (SOAN) of the Altai Mountains and Pre-Altai Plain: Catalogue]; FBGOU VPO Altai: Biisk, Russia, 2013; pp. 1–291. [Google Scholar]
- Dudko, R.Y.; Danukalova, G.A.; Gurina, A.A.; Ivanov, A.V.; Mikhailov, Y.E.; Osipova, E.M.; Prosvirov, A.S.; Solodovnikov, A.Y.; Legalov, A.A.; Zinovyev, E.V. Insects and molluscs of the late Pleistocene at the Gornovo site (Southern Ural foreland, Russia): New data on palaeoenvironment reconstructions. Quat. Int. 2022, 632, 154–177. [Google Scholar] [CrossRef]
- Gurina, A.A.; Dudko, R.Y.; Zinovyev, E.V.; Borodin, A.V.; Tshernyshev, S.E.; Legalov, A.A. Late Pleistocene taphocoenosis of insects and small mammals from the upper reaches of the Ob River. Paleontol. J. 2018, 53, 1610–1622. [Google Scholar] [CrossRef]
- Haslett, J.; Parnell, A. A simple monotone process with application to radiocarbon-dated depth chronologies. J. R. Stat. Soc. C Appl. Stat. 2008, 57, 399–418. [Google Scholar] [CrossRef]
- Dudko, R.Y.; Gurina, A.A.; Dudko, E.R.; Zinovyev, E.V.; Tshernyshev, S.E.; Legalov, A.A. Beetles (Coleoptera) in the late Pleistocene deposits on the Isha River in the foothills of the Northern Altai Mountains. In Proceedings of the XV Congress of the Russian Entomological Society, Novosibirsk, Russia, 31 July–7 August 2017. [Google Scholar]
- Dudko, R.Y.; Lomakin, D.E. Vertical-zonal distribution of carabids (Coleoptera, Carabidae) in the north-east Altai. Sib. J. Ecol. 1996, 3, 187–194. [Google Scholar]
- Tshernyshev, S.; Dudko, R. A new species of the pill beetles (Coleoptera, Byrrhidae) from Siberian Altai Mountains. Entomol. Basil. 1997, 20, 133–142. [Google Scholar]
- Legalov, A.A.; Dudko, R.Y.; Zinovyev, E.V. Sub-fossil weevils (Coleoptera, Curculionoidea) from the central part of West Siberia provide evidence of range expansion during the last glaciations. Quat. Int. 2016, 420, 233–241. [Google Scholar] [CrossRef]
- Chernov, Y.I.; Makarov, K.V.; Eremin, P.K. Family of Ground Beetles (Coleoptera, Carabidae) in the Arctic Fauna: Communication 1. Entomol. Rev. 2000, 80, 1009–1021. [Google Scholar]
- Hågvar, S.; Gobbi, M. The role of arthropods in early colonization near melting glaciers: Contradictions between ecological assumptions and recent study results. Acta Oecol. 2022, 114, 103820. [Google Scholar] [CrossRef]
- Mordkovich, V.G. Successions of Zooedaphone: Their Peculiarities, Novelties and Significance Among Traditional Postulates of Ecology; SB RAS: Novosibirsk, Russia, 2023; pp. 1–150. [Google Scholar]
- Kuzmina, S.A. Macroentomology analysis: Methods, opportunities, and examples of reconstructions of Paleoclimatic and Paleoenvironmental conditions in the Quaternary of the Northeastern Siberia. Contemp. Probl. Ecol. 2017, 10, 336–349. [Google Scholar] [CrossRef]
- Andreev, A.A.; Pierau, R.; Kalugin, I.A.; Daryin, A.V.; Smolyaninova, L.G.; Diekmann, B. Environmental changes in the northern Altai during the last millennium documented in Lake Teletskoye pollen record. Quat. Res. 2007, 67, 394–399. [Google Scholar] [CrossRef]
- Crann, C.A.; Patterson, R.T.; Macumber, A.L.; Galloway, J.M.; Roe, H.M.; Blaauw, M.; Swindles, G.T.; Falck, H. Sediment accumulation rates in subarctic lakes: Insights into age-depth modeling from 22 dated lake records from the Northwest Territories, Canada. Quat. Geochronol. 2015, 27, 131–144. [Google Scholar] [CrossRef]
- Darin, A.V.; Chu, G.; Sun, Q.; Babich, V.V.; Kalugin, I.A.; Markovich, T.I.; Novikov, V.S.; Darin, F.A.; Rakshun, Y.V. Archive data on climate changes and seismic events in glacial clays of Lake Kucherla (Altai region, Russia). Geodyn. Tectonophys. 2020, 11, 624–631. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Webb, N.R.; Coulson, S.J. Primary community assembly on land—The missing stages: Why are the heterotrophic organisms always there first? J. Ecol. 2002, 90, 569–577. [Google Scholar] [CrossRef]
- Mordkovich, V.G.; Lyubechanskii, I.I. Zoological aspects of ecological succession on the graded-flat dump of the Nazarovo lignite open-cast mine in Krasnoyarsk Krai. Contemp. Probl. Ecol. 2019, 12, 346–359. [Google Scholar] [CrossRef]
- Makarova, O.L.; Antipova, M.D.; Babenko, A.B.; Bushueva, I.S.; Chulei, A.D.; Golovatch, S.I.; Doroshina, G.Y.; Kolesnikov, V.B.; Mazei, Y.A.; Makarov, K.V.; et al. Successions of terrestrial invertebrate communities during the Tsey Glacier retreat, Central Caucasus. Caucasiana 2024, 3, 41–87. [Google Scholar] [CrossRef]
- Petrushina, M.N. Features of landscape successions in the zones of debris flows release in the Northern Caucasus. In Debris Flows: Risks, Forecast, Protection; Plyusnin, V.M., Ed.; Institute of Geography SB RAS Publishing House: Irkutsk, Russia, 2016; pp. 199–203. [Google Scholar]
- Nikolaeva, S.A.; Belova (Dirks), M.N. Phytocoenotic assessment of ecotopes on avalanche tracks and debris flow deposits in the Aktru river upper reaches. Vestn. Tomsk. Gos. Univ. Biol. 2017, 40, 181–201. [Google Scholar] [CrossRef]
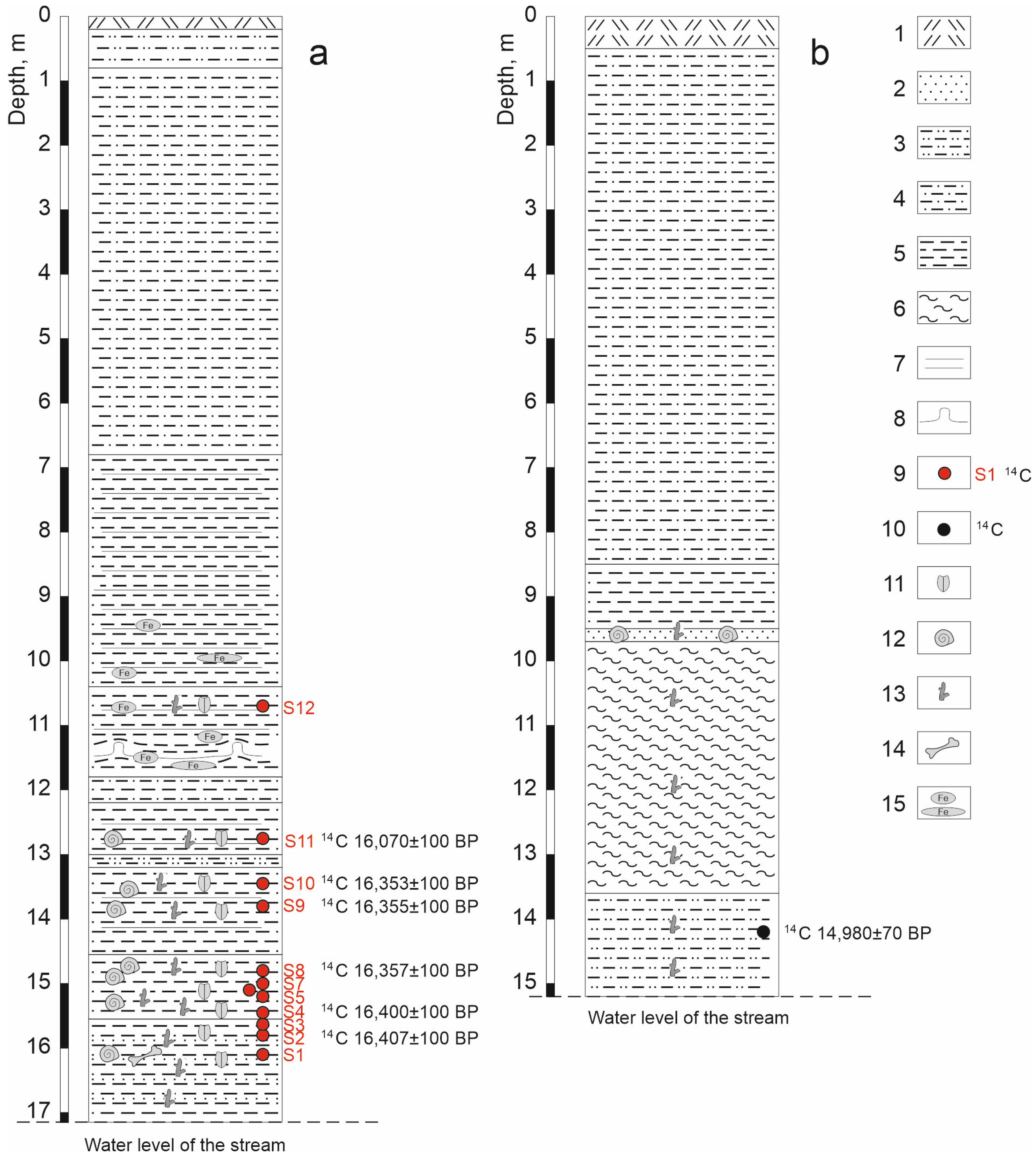




| Layer No | Depth of Bed, m | Elevation, m | Thickness, m | Description |
|---|---|---|---|---|
| 1 | 0.0–0.2 | 16.95–17.15 | 0.2 | Modern soil. |
| 2 | 0.2–0.8 | 16.35–16.95 | 0.6 | Light grey sandy loam, dense. |
| 3 | 0.8–6.8 | 10.35–16.35 | 6.0 | Brown loam, in some places of a red-brown color. |
| 4 | 6.8–10.4 | 6.75–10.35 | 3.6 | Interlayering of light grey clays and reddish grey clays with spots and interlayers of ferrugination. |
| 5 | 10.4–11.8 | 5.35–6.75 | 1.4 | Interlayering of dark grey clays with lenses of plant detritus and light grey clays with spots of ferrugination (sample S12). |
| 6 | 11.8–12.2 | 4.95–5.35 | 0.4 | Dark grey loam with a layer (0.05 m) of dense rufous clay in the middle. |
| 7 | 12.2–13.0 | 4.15–4.95 | 0.8 | Rhythmic horizontal alternation of light-grey clays and dark-grey clays, including lenses and layers of plant detritus (sample S11). |
| 8 | 13.0–13.2 | 3.95–4.15 | 0.2 | Dense rufous sandy loam. |
| 9 | 13.2–14.55 | 2.6–3.95 | 1.35 | Rhythmic horizontal alternation of light-grey clays and dark-grey clays, including lenses and layers of plant detritus (samples S9, S10). |
| 10 | 14.55–15.55 | 1.6–2.6 | 1.0 | Dense light grey non-bedding clay with inclusions of plant detritus and twigs (samples S4–S8). |
| 11 | 15.55–17.15 | 0.0–1.6 | 1.6 | Dense dark grey clay with plant detritus, which goes under the water edge in the creek. In the upper part of the layer, there are interlayers of dark coarse-grained sand (samples S1–S3). |
| Layer No | Sample No | Depth of Sample, m | Number of Subsamples | Subsample Volume, Liters | Radiocarbon Date, ky BP; (Laboratory Code) | Calibrated Date, cal ky BP |
|---|---|---|---|---|---|---|
| 5 | S12 | 10.6–10.8 | 1 | 5 | – | – |
| 7 | S11 | 12.7–12.8 | 3 | 7 | 16.070 ± 0.1 (SPb-4072) | 19.120–19.590 |
| 9 | S10 | 13.4–13.5 | 7 | 1–5 | 16.353 ± 0.1 (SPb-3761) | 19.503–20.006 |
| S9 | 13.75–13.85 | 4 | 1 | 16.355 ± 0.1 (SPb-4073) | 19.505–20.008 | |
| 10 | S8 | 14.75–14.85 | 5 | 10 | 16.357 ± 0.1 (SPb-4074) | 19.507–20.011 |
| S7 | 14.95–15.05 | 3 | 10 | – | – | |
| S6 | 15.05–15.15 | 2 | 10 | – | – | |
| S5 | 15.15–15.25 | 3 | 10 | – | – | |
| S4 | 15.4–15.45 | 5 | 5 | 16.400 ± 0.1 (SPb-4075) | 19.542–20.069 | |
| 11 | S3 | 15.55–15.65 | 3 | 10 | – | – |
| S2 | 15.75–15.85 | 3 | 10 | 16.407 ± 0.1 (SPb-4076) | 19.546–20.078 | |
| S1 | 16.05–16.15 | 2 | 10 | – | – |
| Taxa | In Total | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gastropoda | + | + | – | – | + | – | – | + | + | + | + | + | – | |
| Branchiopoda: | Cladocera | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Ostracoda | – | – | – | – | – | – | – | – | – | – | – | – | – | |
| Arachnida: | Oribatida | + | + | – | – | – | + | – | + | – | – | – | + | – |
| Aranei | 26 | 0 | 0 | 0 | 0 | 4 | 1 | 11 | 6 | 1 | 1 | 2 | 0 | |
| Insecta: | Hemiptera | 10 | 0 | 0 | 0 | 1 | 0 | 0 | 5 | 4 | 0 | 0 | 0 | 0 |
| Coleoptera | 1405 | 108 | 136 | 48 | 232 | 85 | 23 | 141 | 251 | 78 | 185 | 104 | 18 | |
| Hymenoptera | 41 | 2 | 0 | 0 | 5 | 4 | 0 | 5 | 2 | 4 | 12 | 6 | 1 | |
| Diptera | 72 | 3 | 3 | 9 | 18 | 7 | 0 | 13 | 1 | 1 | 5 | 12 | 0 | |
| Index | In Total | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | S11 | S12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
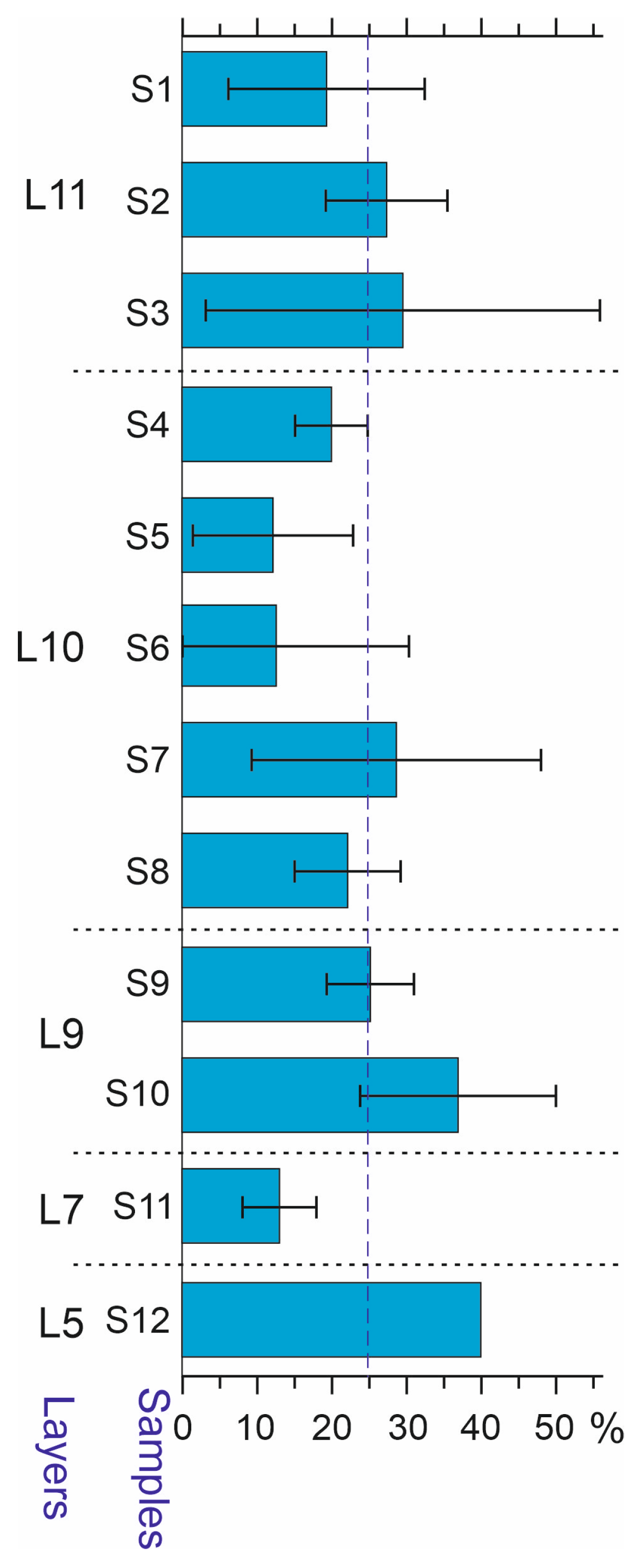
| Nmin | 779 | 61 | 76 | 30 | 130 | 45 | 13 | 70 | 125 | 45 | 113 | 58 | 13 |
| Nmin/V, ind./liter | 5.6 ± 0.9 | 3.1 | 2.5 ± 1.0 | 1.0 ± 0.1 | 5.2 ± 2.5 | 1.5 ± 0.4 | 0.65 | 2.3 ± 0.6 | 2.5 ± 0.7 | 11.3 ± 7.4 | 11.7 ± 4.0 | 2.8 ± 0.7 | 2.6 |
| Species number | 105 | 31 | 32 | 17 | 33 | 23 | 11 | 26 | 38 | 22 | 39 | 34 | 10 |
| Families number | 21 | 8 | 13 | 5 | 13 | 9 | 6 | 8 | 11 | 8 | 11 | 11 | 4 |
| Simpson (1–D) | 0.974 | 0.923 | 0.967 | 0.950 | 0.948 | 0.918 | 0.964 | 0.939 | 0.960 | 0.944 | 0.963 | 0.976 | 0.944 |
| Shannon (H) | 4.06 | 3.15 | 3.51 | 2.85 | 3.22 | 2.98 | 2.51 | 3.06 | 3.45 | 3.06 | 3.53 | 3.63 | 2.22 |
| Ecology | Definition | Species | |
|---|---|---|---|
| Type | Group | ||
| I. Water-associated | 1. Aquatic | Spend the majority of their adult life in water | Hydroporus sp., Helophorus praenanus, H. sibiricus, Ochthebius kaninensis, O. flavipes |
| 2. Peat bogs, fens, and marshland | Habitat across a variety of semi-aquatic environments, such as mires, marsh, swamp, and fens | Phaedon armoraciae, Notaris acridulus, Ceutorhynchus cochleariae | |
| 3. Water edge | Usually live at the edge of ponds, lakes, or slow streams | Nebria gyllenhali, Elaphrus angusticollis, E. riparius, Dyschiriodes nigricornis, Bembidion bipunctatum, B. difficile, B. sajanum, Augyles intermedius | |
| 4. Moist localities | Live in moist localities such as floodplains but not strictly ‘wetlands’ | Loricera pilicornis, Psammoporus matalini, Simplocaria semistriata, Hypnoidus rivularius, Lochmaea caprea, Tournotaris bimaculata, Dorytomus taeniatus, Tachyerges stigma/pseudostigma | |
| II. Open landscapes | 1. Meadow | Prefer grassland and different types of meadows | Sitona lineellus, Phyllobius pomaceus |
| 2. Alpine meadow | High-elevation meadow species | Trechus lomakini, Coccinella nivicola | |
| 3. Tundra | Live in tundra and nival belt of mountains | Diacheila polita, Bembidion dauricum, Hemitrichapion tschernovi | |
| III. Woodland-associated | 1. Woodland | Live in woodland | Notiophilus fasciatus, Carabus aeruginosus, C. obovatus, P. maurusiacus, P. monticoloides, Podabrus alpinus/annulatus, Denticollis acuticollis, Sulcacis nitidus, Sternoplatys fulvipes, Holotrichapion aethiops |
| 2. Xylophagous and xylobionts | Develop in trees | Megatoma undata, Leptophloeus alternans, Pytho depressus, Pissodes gyllenhali, Phloeotribus spinulosus, Carphoborus teplouchovi, Polygraphus polygraphus, P. subopacus, Xylechinus pilosus | |
| IV. Open and woodland landscapes | 1. Boreo-alpine | Live in boreal forests and alpine meadows | Trechus toroticus, Pterostichus fulvescens, P. triseriatus, P. ehnbergi, Agonum alpinum, Coryphium sp., Rhagonycha nigriventris, Eutrichapion rhomboidale, Notaris sp.1, Trichalophus maklini, Otiorhynchus grandineus |
| 2. Forest–tundra | Live in boreal forests and tundra | Carabus arvensis, Pterostichus brevicornis, P. burjaticus, P. subaeneus, Byrrhus mordkovitshi, Boreohypera diversipunctata | |
| 3. Eurytopic | Live in different biotopes or preferences are unclear | Arpedium brachypterum, Mannerheimia brevipennis, Olophrum fuscum, Aphodius ater |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gurina, A.A.; Agrikolyanskaya, N.I.; Dudko, R.Y.; Mikhailov, Y.E.; Prokin, A.A.; Reshetnikov, S.V.; Sazhnev, A.S.; Solodovnikov, A.Y.; Zinovyev, E.V.; Legalov, A.A. Subfossil Insects of the Kebezen Site (Altai Mountains): New Data on the Last Deglaciation Environment. Insects 2025, 16, 321. https://doi.org/10.3390/insects16030321
Gurina AA, Agrikolyanskaya NI, Dudko RY, Mikhailov YE, Prokin AA, Reshetnikov SV, Sazhnev AS, Solodovnikov AY, Zinovyev EV, Legalov AA. Subfossil Insects of the Kebezen Site (Altai Mountains): New Data on the Last Deglaciation Environment. Insects. 2025; 16(3):321. https://doi.org/10.3390/insects16030321
Chicago/Turabian StyleGurina, Anna A., Natalia I. Agrikolyanskaya, Roman Yu. Dudko, Yuri E. Mikhailov, Alexander A. Prokin, Sergei V. Reshetnikov, Alexey S. Sazhnev, Alexey Yu. Solodovnikov, Evgenii V. Zinovyev, and Andrei A. Legalov. 2025. "Subfossil Insects of the Kebezen Site (Altai Mountains): New Data on the Last Deglaciation Environment" Insects 16, no. 3: 321. https://doi.org/10.3390/insects16030321
APA StyleGurina, A. A., Agrikolyanskaya, N. I., Dudko, R. Y., Mikhailov, Y. E., Prokin, A. A., Reshetnikov, S. V., Sazhnev, A. S., Solodovnikov, A. Y., Zinovyev, E. V., & Legalov, A. A. (2025). Subfossil Insects of the Kebezen Site (Altai Mountains): New Data on the Last Deglaciation Environment. Insects, 16(3), 321. https://doi.org/10.3390/insects16030321