Simple Summary
The noctuid species Spodoptera frugiperda (J. E. Smith), Spodoptera litura (Fabricius), and Mythimna separata (Walker) are among the most destructive agricultural pests globally. Currently, the management of these pests largely relies on chemical control methods, which are often inefficient, contribute to the development of insecticide resistance, and pose risks to environmental and food safety. In contrast, biological control using natural enemies, such as predators and parasitoids, has proven to be an effective, environmentally safe, and sustainable alternative. This approach has been successfully implemented in various regions of Africa, Asia, and the Americas. The assassin bug Rhynocoris fuscipes (Fabricius) is a key generalist predator that effectively controls a wide range of pests, including caterpillars, aphids, and planthoppers. However, its predation efficiency against the fall armyworm, common armyworm, and oriental armyworm remains poorly understood. In this study, we assessed the potential of R. fuscipes as a candidate biocontrol agent for managing these armyworm pests by examining its functional response, intraspecific interference competition, and prey preference. Our findings suggest that R. fuscipes shows strong potential as a biocontrol agent against these noctuid pests and provide valuable insights for optimizing its use in integrated pest management strategies.
Abstract
Understanding predator–prey and predator–predator interactions is essential for evaluating the effectiveness of biocontrol agents and developing efficient pest management strategies. This study investigates the effects of prey species, predator life stage, and predator density on the predatory efficiency of the generalist predator Rhynocoris fuscipes (Fabricius) under semi-field conditions. Both the nymphs and adults of R. fuscipes consumed significantly more second-instar larvae of S. frugiperda than larvae of S. litura or M. separata. Notably, fifth-instar nymphs and adults exhibited higher predation capacity (11.75 ± 0.37 and 10.90 ± 0.40 larvae) than fourth-instar nymphs (9.05 ± 0.29 larvae) on S. frugiperda. R. fuscipes demonstrated a Type II functional response at all developmental stages toward each prey species, and fifth-instar nymphs revealed higher attack rates (a = 1.5205 ± 0.0544) on S. frugiperda; additionally, the handling time did not significantly differ among three prey species. It suggests that this predator may be more effective at controlling noctuid populations at low prey densities. As predator age and density increased, intraspecific competition among R. fuscipes also intensified. In a multi-prey system, R. fuscipes showed a marked preference for S. frugiperda over the other two prey species. These findings indicate that the fifth-instar nymphs and adults of R. fuscipes are particularly effective in suppressing early-instar larvae of the three noctuid pests, especially S. frugiperda, at low densities. This study enhances our understanding of the predation capacity and prey preference of R. fuscipes towards the three noctuid pests and provides a foundation for the development of more targeted and efficient pest management strategies using this predator.
1. Introduction
Armyworms are pests that attack a wide range of crops, especially cereals [1,2,3]. Most armyworm species cause severe damage by feeding gregariously and voraciously on leaves and seedlings [4]. Common armyworm species, including the fall armyworm Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae), the common armyworm Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae), and the oriental armyworm Mythimna separata (Walker) (Lepidoptera: Noctuidae), are all highly polyphagous agricultural pests with a global distribution [5,6,7]. Due to their rapid growth, large food intake, high reproductive rate, and strong dispersal capacity, these pests have become highly destructive, causing significant economic losses in various crops [8,9,10]. The notorious migratory pest S. frugiperda, native to the tropical and subtropical regions of the Americas, was first detected in Central and Western Africa in early 2016. These pests caused annual maize losses of up to 17.7 million tons in 12 African countries, a loss large enough to feed tens of millions of people in 2018 [11,12,13]. Subsequently, it invaded China in December 2018, then spread through 26 provinces (autonomous regions and municipalities) in 2019 and 27 in 2020, damaging 1.125 and 1.278 million hectares of crops, respectively [14]. The common cutworm S. litura is widely distributed across temperate and subtropical regions, with its larvae capable of feeding on over a hundred economically important crops, including maize, rice, soybean, and cotton [15,16]. In pepper fields, large infestations of S. litura can damage the bare stalks, leading to significant losses [17]. The oriental armyworm M. separata is a migratory and polyphagous pest species found throughout Asia, Oceania, and several Pacific islands. It causes severe damage to rice, maize, wheat, vegetables, and other crops [18]. Between 2012 and 2013, an outbreak of M. separata in China threatened approximately 1743.7 million hectares of farmland, and this threat has continued in recent years [18]. The frequent or sudden appearances of these armyworms have led to extensive insecticide usage, resulting in increased pesticide resistance among pests and numerous adverse effects on non-target organisms [19,20,21,22]. Therefore, an integrated and sustainable pest management strategy is essential for the prevention and control of these lepidopteran pests, which is crucial for agricultural production and global food security [23,24,25].
Augmenting and conserving natural enemies to mitigate the impacts of pests is a fundamental strategy in biological control, which is both environmentally sustainable and cost-effective strategy [26]. This approach plays a crucial role in integrated pest management (IPM) programs. Among the biological control agents, Heteropteran predatory bugs are known for their broad prey range, with several species demonstrating significant predation capacity against key agricultural pests such as S. frugiperda, S. litura, and M. separata [27,28,29,30,31,32,33]. Notable examples include Arma chinensis (Fallou) (Hemiptera: Pentatomidae) [27,28,29], Picromerus lewisi (Scott) (Hemiptera: Pentatomidae) [30,31,32], and Rhynocoris marginatus (Fabricius, 1794) (Hemiptera: Reduviidae) [33]. However, only a few species of predatory bugs, such as Podisus maculiventris (Say) (Hemiptera: Pentatomidae), Cantheconidea furcellata (Wolff) (Hemiptera: Asopinae), Geocoris spp. (Hemiptera: Lygaeidae), and Orius sauteri (Poppius) (Hemiptera: Anthocoridae), have been successfully mass-reared for commercial use in field or greenhouse applications [34]. The selection and evaluation of additional potential predators are critical for advancing biological control strategies against major agricultural pests.
The assassin bug Rhynocoris fuscipes (Fabricius) (Heteroptera: Reduviidae) is a generalist predator that is widely distributed across East Asia, including China, India, Japan, and Vietnam [35]. Both the nymphs and adults of this predator prey on a variety of agricultural pests, such as the brown planthopper Nilaparvata lugens (Stal) (Hemiptera: Delphacidae), the mealybug Phenacoccus solenopsis (Tinsley) (Hemiptera: Pseudococcidae), the cotton stainer Dysdercus cingulatus (Fab.) (Hemiptera: Pyrrhocoridae), Dysdercus koenigii (Fab.) (Hemiptera: Pyrrhocoridae), and several aphid species [36,37,38,39]. As an effective biological control agent for agriculture pests, R. fuscipes is commonly found in agroecosystems throughout southern China [39]. Given its polyphagous nature, this predator may also play significant role in controlling noctuid pests. Understanding its prey range, prey preference, and predation capacity is essential for optimizing its use in biological control programs.
Functional response describes the number of prey consumed per unit time by a predator as prey density changes [40]. It is a crucial tool for evaluating the potential of biological control agents, as it quantifies key aspects of predation behavior, foraging ecology, digestion, and fitness-related factors [19,29,41,42,43]. Holling (1959) identified three types of functional response (Type I, Type II, and Type III) [44]. The most commonly observed and widely used types in predator–prey studies are Type II and Type III, where per capita predation increases curvilinearly and sigmoidally with prey density, respectively [44,45]. In contrast, Type I represents a linear relationship between prey density and the maximum number of prey killed. In previous research, functional responses are typically measured in Petri dishes under laboratory conditions [46]. In addition to prey density, the functional response can also be influenced by various factors, including predator density, competitor densities, temperature, predator age, spatial factors, host plants, and the use of insecticides [47,48,49,50,51]. To enhance the efficacy of predators in pest suppression, it is crucial to study their consumption ability through functional response assessments under field-simulated conditions. Furthermore, analyzing intraspecific competition and prey preference is vital for determining a predator’s predation capacity and evaluating its potential as a candidate for biological control programs [52,53].
In this study, we evaluated the potential of R. fuscipes as a biological control agent for managing three major noctuid pests (S. frugiperda, S. litura, and M. separata) by conducting functional response and intraspecific interference competition experiments under semi-field conditions. The functional response of the fourth- and fifth-instar nymphs, as well as adult R. fuscipes, to second-instar larvae of each prey species was assessed on maize plants within cages. In addition, we examined intraspecific interference competition among R. fuscipes at different developmental stages. This study also compared the consumption rates and prey preference of R. fuscipes for S. frugiperda, S. litura, and M. separata. The findings will offer valuable insights into the predator–prey and predator–predator interactions between R. fuscipes and its prey, contributing to the development of more effective strategies for controlling these noctuid pests using R. fuscipes.
2. Materials and Methods
2.1. Insect Colonies
The colony of the assassin bug R. fuscipes was originally collected from a field in Guangdong, China, and has been maintained in our laboratory in Beijing for over three generations. Both nymphs and adults of R. fuscipes were reared in a cage (6 cm length, 6 cm width, and 1.5 cm height) with Tenebrio molitor (Coleoptera: Tenebrionidae) under controlled conditions of 28 ± 1 °C, 16 h of light per day (16L:8D), and 70 ± 5% relative humidity (RH) in an artificial climate chamber. For experimental purposes, fourth- and fifth-instar nymphs and adults of R. fuscipes, selected 2 to 3 days after molting or eclosion, were used. These insects were starved for 24 h before being placed into experimental arenas to assess their functional response, intraspecific interference competition, and prey preference.
Colonies of S. frugiperda, S. litura, and M. separata have been maintained in our laboratory for at least two years. The larvae of these species were reared on fresh maize leaves and improved artificial diets as described for S. frugiperda [54,55], S. litura [55], and M. separata [55], respectively. Adults were provided with a 10% sucrose solution. All insect colonies were reared in climate chambers under conditions of 26 ± 1 °C, 16L:8D, and 70 ± 5% RH. Second-instar larvae (molted within 1 to 2 days) of each prey species were used in all subsequent experiments.
2.2. Experimental Conditions
To create experimental arenas that closely mimic natural conditions, we assessed the predation rate and prey preference of R. fuscipes towards armyworms on maize plants within mesh cages (100 mesh, 20 cm in length, 20 cm in width, and 35 cm in height). Each cage contained a flowerpot (15.5 cm top diameter, 11.5 cm bottom diameter, and 14 cm height) in which fifteen maize seedlings were planted, growing until they reached the two-leaf stage (after 14 days). Prey individuals were randomly distributed across all maize plants. All experiments were conducted in an artificial climate chamber at 28 ± 1 °C, with a photoperiod of 16L:8D and 70 ± 5% RH. Prey individuals were not replaced during the experiments.
2.3. Functional Response
To examine the functional response of R. fuscipes to different prey densities, we tested the consumption rates of fourth- and fifth-instar nymphs, as well as male and female adults, on armyworms at prey densities of 5, 10, 15, 20, and 30 larvae per predator (24 h starved). After 24 h, the number of armyworms consumed by the predator was recorded. Control treatments, without predators, were also used to assess the natural mortality of the prey species, enabling the correction of prey consumption rates for natural mortality. Each treatment included 20 predators as replicates at each prey density.
2.4. Intraspecific Interference Competition
The effects of intraspecific interference competition on the predation rate and foraging behavior of R. fuscipes were also investigated. This was carried out using fourth- and fifth-instar nymphs, as well as male and female adults, exposed to prey densities of 50, 100, 150, 200, and 250 armyworms, with varying numbers of R. fuscipes predators (1, 2, 3, 4, or 5 individuals) in the experimental arenas. The prey-to-predator ratio was maintained at 50 for each group of predators placed in a cage. As the number of predators increased, so did competition for space. After 24 h, the number of surviving prey was counted. The daily predation rate by R. fuscipes was calculated, and the coefficients of intraspecific interference were analyzed. Each treatment was replicated 10 times.
2.5. Prey Preference
To assess the predation preferences of R. fuscipes for different prey species, including S. frugiperda, S. litura, and M. separata, experiments were conducted using 24 h starved R. fuscipes nymphs (fourth and fifth instars) and adults (both females and males). In each trial, 10 instar larvae from each prey species (total of 30 prey larvae) and 1 predator were introduced into an experimental arena. The number of prey consumed by each stage of R. fuscipes was recorded, and the total predation rate for each developmental stage was calculated. For each predator life stage, 10 replicates were performed, with one predator per repetition.
2.6. Statistical Analyses
In the control groups, no mortality of S. frugiperda, S. litura, or M. separata larvae was observed, and therefore, prey mortality data were not included in the analysis. A one-way analysis of variance (ANOVA) was used to compare prey consumption and predation rates across different developmental stages of R. fuscipes at varying prey densities. Significant differences between treatments were identified using Duncan’s multiple range test (p < 0.05). Additionally, comparisons of prey consumption and preference among different developmental stages of R. fuscipes for the three prey species were performed using one-way ANOVA, followed by Duncan’s multiple range test (p < 0.05).
To determine the type of functional response, cubic logistic regression analysis was conducted to examine the relationship between the proportion of prey consumed and initial prey density as follows:
where Ne represents the number of prey consumed, and N0 denotes the prey density. The parameters P0, P1, P2, and P3 correspond to the intercept, linear, quadratic, and cubic coefficients, respectively [45]. A significant negative linear coefficient (P1 < 0) indicates a Type II functional response. Conversely, when the linear coefficient is significantly positive (P1 > 0) and the quadratic coefficient is negative (P2 < 0), the predator exhibits a Type III functional response [45]. The logistic regression analysis confirmed that the data for each case followed a Type II functional response. Therefore, the random predator equation (Equation (2)) was applied to model the relationship between the number of prey consumed (Ne) and the initial prey density (N0) [56]:
where Ne represents the number of prey consumed, N0 is the initial prey density, a denotes the attack rate, T is the total searching time (one day), and Th is the handling time (in days). The ratio T/Th is used to calculate the theoretical maximum prey consumption, while the functional response rate (FRR = a/Th) [42] provides an estimate of the predation efficiency of R. fuscipes. To compare differences in the attack rate and handling time across the developmental stages of R. fuscipes in response to the three prey species, the extra sum-of-squares F-test was applied. Parameter estimates were obtained using the non-linear least squares regression procedure.
To determine the effect of predator densities on the prey consumption of R. fuscipes, the parameters for intraspecific competition were estimated using non-linear regression analysis by fitting the model described in Equation (3) [57]:
In this model, E (where represents the predation efficency, p denotes predator density, Q is the search constant, and m is the mutual interference constant. A chi-square goodness-of-fit test was performed to evaluate whether the intraspecific competition model appropriately fits the data.
Prey preference was evaluated using Ivlev’s selection index [58], and the selectivity index Ci was calculated according to Equation (4):
In this equation, Ci represents the predator’s preference for prey type i, Fi refers to the proportion of prey type i in the experimental system, and Qi denotes the proportion of prey type i consumed by the predator. A Ci value between 0 and 1 indicates a positive preference, a Ci value between −1 and 0 indicates a negative preference, and Ci = 0 suggests no preference.
All data were checked for normality and homoscedasticity. Statistical analyses were performed using Prism 9.0 (GraphPad), while regression analyses were conducted using SigmaPlot (version 14.0, Systat Software, Inc., San Jose, CA, USA).
3. Results
3.1. Prey Consumption
The nymphs and adults of R. fuscipes had high consumption of the second-instar larvae of S. frugiperda, S. litura, and M. separata, respectively. In all experimental treatments, the number of prey consumed by R. fuscipes over a 24 h period increased significantly with prey density and then plateaued once the prey density reached an upper asymptote (Figure 1). Regardless of prey type, the fourth-instar nymphs of R. fuscipes showed the lowest mean daily prey consumption (i.e., consumed 8~9.5 armyworms per day) at the highest prey densities, with no significant differences observed among the fifth-instar nymphs, female and male adults of the predator (Table 1). For all life stages of R. fuscipes, predation was highest on S. frugiperda, followed by S. litura, and lowest on M. separata (Table 1). Notably, both the fifth-instar nymphs and male adults of R. fuscipes consumed significantly more S. frugiperda than S. litura and M. separata.
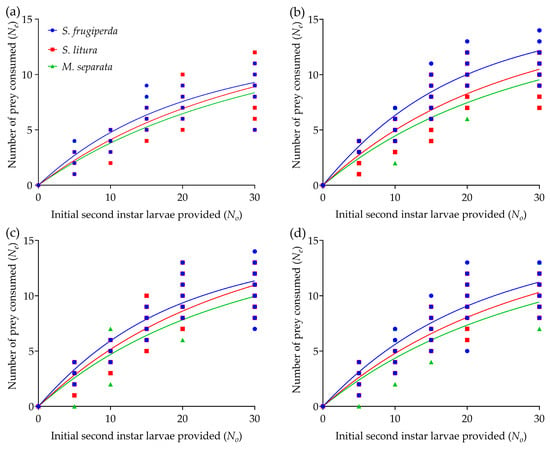
Figure 1.
Predation functional responses of the fourth-instar nymph (a), fifth-instar nymph (b), female adult (c), and male adult (d) of R. fuscipes to the second-instar larvae of S. frugiperda, S. litura, and M. separata at different prey densities. The data points represent the number of prey consumed by R. fuscipes over 24 h. The curves represent the predicted values based on Rogers’ random predator equation.

Table 1.
Prey consumption (±SE) per day for nymphs and adults of R. fuscipes of the second-instar larvae of S. frugiperda, S. litura, and M. separata at the highest prey density, respectively.
3.2. Functional Response
The maximum likelihood estimate of the linear parameter P1 was negative for all life stages of R. fuscipes (i.e., P1 < 0) (Table S1), indicating a Type II functional response of each developmental stage of the predator to each prey species, as revealed by the logistic model analysis (Figure 1). The functional response data for R. fuscipes feeding on S. frugiperda, S. litura, or M. separata over a 24 h period closely matched the random predation equation, further supporting the conclusion of a Type II response across all life stages of R. fuscipes (Table 2).
Table 2.
Parameter estimates from the Holling II functional response of different life stages of R. fuscipes preying on the second-instar larvae of S. frugiperda, S. litura, and M. separata, respectively.
With the development of the predator, the attack rate coefficient (a) of R. fuscipes increases, while the handling time (Th) decreases (Table 2). For each prey species, both adults and fifth-instar nymphs of R. fuscipes generally showed a higher attack rate but shorter handling time compared to earlier developmental stages (Table 2). The functional response parameters of the predator were also influenced by the prey species. For each developmental stage of R. fuscipes, the highest attack rate was observed in predators feeding on S. frugiperda, with no significant differences observed between predators feeding on the S. litura and M. separata (F= 60.829, df = 57, p < 0.0001). Although the female adults of R. fuscipes feeding on S. litura displayed the shortest handling time, there were no significant differences among the three prey species (Table 2). Based on the theoretical maximum consumption parameter (T/Th), the consumption rate by R. fuscipes increased with developmental stage, with female adults showing greater predation on S. litura than on other prey species. The predatory efficiency of R. fuscipes can be more objectively and accurately reflected by the functional response ratio (FRR = a/Th) [19,59], where a higher FRR value indicates greater predation efficiency. Among the various life stages, R. fuscipes exhibited the highest predation efficiency on S. frugiperda and the lowest on M. separata. Fifth-instar nymphs and adults generally had higher FRR values than fourth-instar nymphs. The highest predation efficiency was observed in the fifth-instar nymph of R. fuscipes feeding on S. frugiperda (FRR = 28.0718), followed by the female adults (FRR = 23.7099) and male adults (FRR = 21.9453) (Table 2).
3.3. Intraspecific Interference Competition
All models converged on non-zero estimates for the predator interference parameter. When the prey-to-predator ratio was kept at 50, an increase in both predator and prey numbers led to a gradual rise in the total consumption of the three prey species by R. fuscipes in the cages. Across all conditions, the average daily consumption of each prey species by R. fuscipes at different life stages significantly decreased as predator density increased (Table 3, Figure 2). For all predator–prey density combinations tested, the intraspecific interference competition model fit the empirical data well, as indicated by the asymptotic 95% confidence intervals excluding zero and relatively high R-squared values (Table 4).
Table 3.
Average daily predation rate by different life stages of R. fuscipes on second-instar larvae of S. frugiperda, S. litura and M. separata at different predator densities.
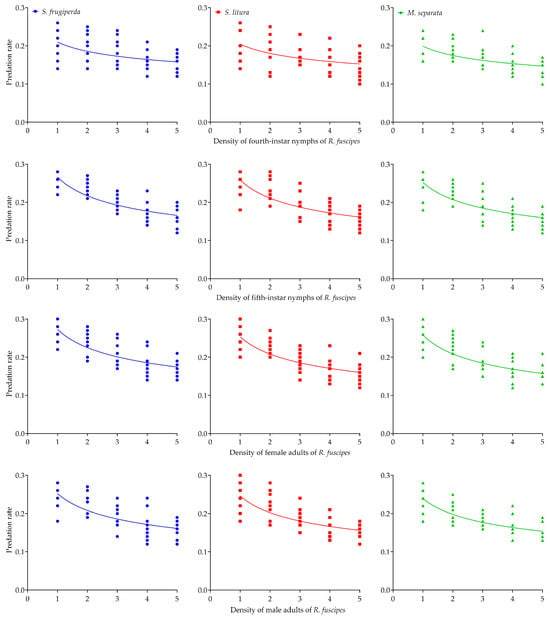
Figure 2.
The intraspecific interference competition of the fourth-instar and fifth-instar nymphs as well as the female adults and male adults of R. fuscipes preying on the second-instar larvae of S. frugiperda, S. litura, and M. separata, respectively.
Table 4.
Parameters estimated from the intraspecific competition interference equation of different life stages of R. fuscipes preying on the second-instar larvae of S. frugiperda, S. litura, and M. separata, respectively.
The intraspecific competition model effectively captured the predation rates at varying densities of R. fuscipes, regardless of prey species (Table 4). The asymptotic 95% confidence intervals, which did not include zero, were evident in the quest search constant (Q) and mutual interference constant (m) for all life stages of R. fuscipes. In nearly every case, R. fuscipes showed the highest predation rate on S. frugiperda and the lowest on M. separata. The exception was observed in the female adults of R. fuscipes, where the predation rate on M. separata exceeded that on S. litura. As R. fuscipes developed, intraspecific interference competition among the predators progressively intensified (Table 4, Figure 2).
3.4. Prey Preference
When all three prey species were offered simultaneously, R. fuscipes showed higher predation on S. frugiperda than on S. litura or M. separata at every life stage, although the differences were not statistically significant (Figure 3a). All life stages of R. fuscipes demonstrated a positive preference for the second-instar larvae of S. frugiperda and S. litura, while showing a negative preference for M. separata, except for the fifth-instar nymphs of the predator (Figure 3b).
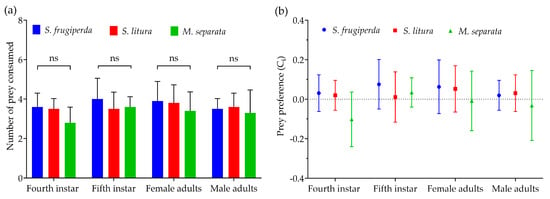
Figure 3.
The mean consumption rate (a) and prey preference (b) of the fourth-instar and fifth-instar nymphs as well as the female adults and male adults of R. fuscipes consuming second-instar larvae of S. frugiperda, S. litura, and M. separata, respectively.
4. Discussion
Our results suggest that the reduviid predator R. fuscipes has potential as an effective biocontrol agent against the major agricultural pests S. frugiperda, S. litura, and M. separata. This is primarily due to the greater predatory efficiency exhibited by older nymphs and adults of R. fuscipes, which were more effective at preying on the second-instar larvae of each prey species. Notably, R. fuscipes nymphs and adults, particularly the fifth-instar nymphs, demonstrated a higher consumption rate and preference for S. frugiperda over S. litura and M. separata. A similar pattern was observed in the fifth-instar nymphs of the closely related reduviid predator R. marginatus, which also consumed more S. frugiperda larvae than other prey life stages [33]. However, the female adults of R. fuscipes consistently exhibited the highest predation rates on S. litura and M. separata larvae compared to other developmental stages. This finding is in line with the work of George and Seenivasagan, who found that the female adults of R. marginatus had the highest predation rates on S. litura [60]. These results indicate that both the adult and fifth-instar nymph stages of R. fuscipes are particularly suitable for potential use in biological control programs.
Type II functional responses are commonly observed in insect predators [19,61,62], including predatory stinkbugs such as O. sauteri (Poppius) [63], A. chinensis [27], P. lewisi [32], Dicyphus bolivari (Lindberg), and Dicyphus errans (Wolff) [43]. In our study, R. fuscipes displayed Type II functional responses at the adult and fifth- and fourth-instar nymphal stages when preying on the larvae of S. frugiperda, S. litura, and M. separata. Previous studies have also reported Type II functional responses in R. fuscipes when feeding on other pests, such as the cotton mealybug P. solenopsis [36], the red cotton bug D. koenigii [36], the Russian wheat aphid Diuraphis noxia (Mordvilko) [47], the cucumber leaf folder Diaphania indicus (Saunders) [64], and the teak skeletonizer Eutectona machaeralis (Walker) [65]. In contrast, some predatory species, such as the minute pirate bug Anthocoris minki (Dohrn) preying on Psyllopsis repens [48] and Nabis sp. preying on both the leafhopper species Agallia constricta and Ceratagallia agricola [66], have shown a Type III functional response. In these cases, prey consumption remains low at low prey densities but increases at higher densities, a characteristic of Type III functional responses. Type II functional responses may lead to instability in predator–prey dynamics, as high predation rates at low prey densities can cause prey population collapse. Conversely, Type III functional responses are considered more stabilizing, as they allow for prey refuge at low densities or alternative prey switching, while increasing foraging efficiency at high prey densities [62,67]. Thus, predators exhibiting Type II functional response may be more effective in controlling pest populations at lower prey densities, making them ideal candidates for augmentative biological control strategies, especially when predator–prey ratios are high.
However, under complex field conditions, the functional response of predators exhibiting Type II behaviors could shift to Type III due to factors such as temperature [68], predator developmental stage [67], starvation levels [69], prey switching [70], host plants [47], space availability [71], and foraging area size [61]. A precise and reliable identification of functional response types is critical for understanding predator–prey dynamics and informing the optimal use of biocontrol agents. Experimental settings with limited prey availability may result in estimates of functional responses that represent theoretical maximum interactions under those specific conditions. Kalinkat et al. emphasized that the importance of considering appropriate prey density ranges, habitat structure, and biologically relevant designs when conducting functional response experiments [72]. Therefore, it is crucial to conduct further studies on the functional response and predatory efficiency of R. fuscipes under semi-field or field conditions targeting a range of pests across various crops. Such studies will provide more robust insights into the predator’s potential as a biocontrol agent and inform its practical application in integrated pest management systems.
The key indicators of predatory functional responses are the attack rate and handling time, which reflect the theoretical efficiency of predators in capturing, killing, and consuming prey [73]. Our results clearly demonstrate the significant influence of both prey species and predator developmental stages on these two key parameters (Table 2). At the same developmental stage, R. fuscipes exhibits distinct species-specific differences in attack rates and prey handling times when preying on three noctuid pests with similar host ranges and body sizes. These variations underscore the complexity of foraging strategies employed by R. fuscipes. Moreover, as predators mature, their body size, strength, and predatory skills improve, potentially leading to increased attack rates and enhanced handling efficiency [70]. For instance, when preying on S. litura and M. separata, both the attack rates and handling efficiency of R. fuscipes increased progressively with its development. Interestingly, when preying on S. frugiperda, the fifth-instar nymphs of R. fuscipes exhibited a significantly higher attack rate than adults, while prey handling efficiency did not differ substantially between the two stages. This finding is similar to the observations of Sahayaraj et al., where the fifth-instar nymphs of Rhynocoris longifrons (Stål) had higher attack rates than adults when preying on Aphis gossypii (Glover) [74]. However, in Rhynocoris kumarii Ambrose, the attack rate of fifth-instar nymphs towards the prey P. solenopsis Tinsley was higher than that of male adults but lower than that of female adults, with significant differences in prey handling efficiency [75]. Additional factors, such as temperature [76,77], host plant species [78], search area [77], hunger level [67,79] and insecticide exposure [80,81], also significantly influence attack rates and handling times. However, relying solely on attack rates and handling times may lead to contradictory predictions regarding predatory efficiency. To address this, a new metric, the parameter FRR (a/Th) has been proposed [19]. Our results indicate that the FRR (a/Th) of R. fuscipes is also influenced by both prey species and predator developmental stage. When S. litura was the prey, R. fuscipes reached maximum predatory efficiency at the adult stage. These results suggest that predators at different developmental stages may encounter different ecological challenges and predation pressures, leading to hierarchical differences in their functional responses.
Intraspecific competition occurs when predators of the same species compete for resources, such as food or territory, and can significantly impact behavior, population dynamics, and ecosystem functioning [82]. Our results indicate that when the prey-to-predator ratio was maintained at 50, the average daily consumption of the three noctuid pests by R. fuscipes decreased as the predator density increased (Table 3 and Table 4). This decline in predation efficiency is likely due to intensified spatial competition and resource contention at higher predator densities. Additionally, the interaction between prey species and predator developmental stage appears to modulate intraspecific interference. Regardless of prey type, the fifth-instar nymphs and adults of R. fuscipes experienced significantly stronger intraspecific interference than the fourth-instar nymphs. This was particularly evident when S. frugiperda and M. separata were the prey, with the highest levels of intraspecific interference occurring at the fifth-instar nymph and adult stages of R. fuscipes, likely due to maximum prey consumption at these stages, which intensified competition for resources (Table 1). Interestingly, although R. fuscipes adults consumed the most S. litura, intraspecific interference was more pronounced during the fifth-instar nymph stage. This suggests that other factors, such as the hunger level of predators, may also play a significant role in shaping interference dynamics [83]. Studies have shown that the hunger level of predators can influence intraspecific competition; once predators reach satiety, their activity levels increase, and they shift from foraging to mating behaviors, which can exacerbate interference, especially among same-sex adults [81,82,83]. This may partially explain the stronger interference effects observed in the adult stage compared to other developmental stages of R. fuscipes. Understanding intraspecific competition is crucial for optimizing predator population management and enhancing pest control efficiency, allowing for reduced interference while maintaining high predation performance. However, the optimal release ratio of natural enemies remains to be validated through field trials.
In field conditions, the presence of alternative prey (non-target pest) may alter the predation efficiency and functional response of generalist predators to target pests [84,85]. Given the variety of available prey, understanding the predator’s feeding preferences is critical for enhancing its application in biological control programs [19]. In our study, the generalist predator R. fuscipes at different developmental stages showed a preference for consuming S. frugiperda and S. litura over M. separata in a multi-prey system. This aligns with findings by Nagarajan et. al., who used a Y-shaped olfactometer to analyze the prey preferences of adult R. fuscipes. They reported that R. fuscipes exhibited the highest preference for S. litura compared to other prey species such as Helicoverpa armigera Hubner, Achaea janata Linnaeus, and D. cingulatus and Mylabris indica Thunberg [86]. However, the fifth-instar nymphs of R. fuscipes showed a positive selectivity towards M. separata. The feeding preferences of predators may be influenced by factors such as prey size, nutritional quality, behavioral traits, and chemical cues [85,87]. For example, R. fuscipes prefers feeding on soft-bodied prey such as termites (Odontotermes obesus Rambur) over more chitinized prey such as grasshoppers (Oxya sp.) [88]. Similarly, when offered a choice between D. cingulatus, S. litura, and castor pest A. janata, R. fuscipes responded most strongly to a chloroform–methanol extract of A. janata, with the least response to S. litura extract [89]. Given that the three noctuids (S. frugiperda, S. litura, and M. separata) used in this study share similar host plant ranges [8,9,10], we hypothesize that chemical pheromones released by S. frugiperda or S. litura may attract R. fuscipes to these prey. In natural field settings, these three armyworms often occur simultaneously, highlighting the need for further studies on the predatory behavior and consumption rates of R. fuscipes in complex multi-prey environments.
It is important to note that S. frugiperda, S. litura, and M. separata enter a highly destructive feeding phase following the third-instar, which significantly exacerbates their damage to crops [90,91,92]. Effective population control prior to the end of the third-instar can substantially reduce crop loss and economic losses. This study focuses primarily on evaluating the biological control efficacy of various developmental stages of R. fuscipes against the second-instar larvae of these pest species. Since eggs and first-instar larvae are more vulnerable to natural enemies and third-instar larvae represent a critical stage in the pest’s voracious feeding cycle, further exploration of R. fuscipes’ predation capabilities against their earlier and later pest stages is warranted. This will enhance our understanding of its functional response and provide a basis for developing more targeted and efficient pest management strategies, ultimately improving the application of R. fuscipes in integrated pest management.
5. Conclusions
In conclusion, we provide evidence that R. fuscipes is a promising biological control agent for controlling early larval stages of S. frugiperda, S. litura, and M. separata. The most effective predatory stages are the fifth-instar nymphs and adults, which have higher foraging efficiency compared to earlier developmental stages. R. fuscipes showed a Type II functional response to all prey species, suggesting that it is likely to be more effective in suppressing noctuid populations at lower prey densities. Intraspecific competition among R. fuscipes individuals intensifies with both predator development and increasing population density. Notably, R. fuscipes appears to be more efficient in controlling S. frugiperda than the other two noctuid pests, as evidenced by its superior predation capacity and prey preference. Considering factors such as predatory effectiveness and intraspecific interference, our findings suggest that releasing fourth-instar nymphs of R. fuscipes during periods of low noctuid larval density would optimize the efficacy of biological control. These results contribute valuable insights into predator–prey dynamics, predator–predator interactions, and the foraging behavior of R. fuscipes. However, further studies are needed to assess its utility in managing armyworms across various crops and in complex agroecosystems. Future research should focus on evaluating the influence of prey stage, prey switching, predator–prey ratio, and environmental factors such as temperature on the predation efficiency of R. fuscipes under field conditions. This will be essential for developing a comprehensive and effective integrated pest management strategy.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/insects16020224/s1, Table S1: Logistic models describing the proportion of second-instar larvae of S. frugiperda, S. litura, and M. separata consumed by different developmental stages of R. fuscipes as a function of initial prey density.
Author Contributions
Conceptualization, C.X., Y.L., and M.W.; methodology, C.X. and Y.L.; software, C.X., J.M. (Jiaying Mao) and B.X.; validation, C.X., L.Z. (Lei Zhou), and H.Z.; formal analysis, C.X., J.M. (Jianjun Mao) and Z.S.; investigation, L.Z. (Lisheng Zhang), Y.L. and M.W.; resources, L.Z. (Lisheng Zhang); data curation, C.X. and L.Z. (Lei Zhou); writing—original draft preparation, C.X.; writing—review and editing, Y.L. and M.W.; visualization, Y.L. and M.W.; supervision, L.Z. (Lisheng Zhang); funding acquisition, Y.L., M.W. and L.Z. (Lisheng Zhang). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key R&D Program of China (2023YFE0123000; 2021YFD1400703).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank our funders and the anonymous reviewers, who have helped improve our manuscript substantially.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chimweta, M.; Nyakudya, I.W.; Jimu, L.; Mashingaidze, A.B. Fall armyworm [Spodoptera frugiperda (J.E. Smith)] damage in maize: Management options for flood-recession cropping smallholder farmers. Int. J. Pest Manag. 2020, 66, 142–154. [Google Scholar] [CrossRef]
- Li, Y.Y.; Qu, C.; Zhang, Q.H.; Zhang, L.P.; Luo, C.; Wang, R. Baseline Susceptibility, Cross-Resistance, and Sublethal Effects of Broflanilide, a Novel Meta-Diamide Pesticide, in Spodoptera litura. Int. J. Mol. Sci. 2023, 24, 5351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.B.; Liu, H.W.; Liu, Y.P.; Wang, C.; Ma, B.W.; Zhang, M.J.; Zhang, Y.; Liu, Y.; Yang, B.; Wang, S.; et al. Chromosome-level genomes of two armyworms, Mythimna separata and Mythimna loreyi, provide insights into the biosynthesis and reception of sex pheromones. Mol. Ecol. Resour. 2023, 23, 1423–1441. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.P.; Guo, J.F.; Jiang, Y.Y.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2020, 27, 780–790. [Google Scholar] [CrossRef]
- Lu, Y.H.; Tian, J.; Ullah, F.; Desneux, N.; Guo, J.W.; Wang, S.S.; Xu, H.X. Sublethal and transgenerational effects of lufenuron on biological characteristics and expression of reproductive related genes in the fall armyworm, Spodoptera frugiperda. Pestic. Biochem. Physiol. 2023, 196, 105593. [Google Scholar] [CrossRef]
- Li, K.; Chen, J.; Jin, P.; Li, J.; Wang, J.; Shu, Y. Effects of Cd accumulation on cutworm Spodoptera Litura larvae via Cd treated Chinese flowering cabbage Brassica campestris and artificial diets. Chemosphere 2018, 200, 151–163. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.H.; Wang, J.; Liu, J.; Tang, Q.B.; Xiang-Rui, L.; Cheng, D.F.; Zhu, X. Analysis on the migration of first-generation Mythimna separata (Walker) in China in 2013. J. Integr. Agric. 2018, 17, 1527–1537. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Hoesain, M.; Suharto; Prastowo, S.; Pradana, A.P.; Alfarisy, F.K.; Adiwena, M. Investigating the plant metabolite potential as botanical insecticides against Spodoptera litura with different application methods. Cogent Food Agric. 2023, 9, 2229580. [Google Scholar] [CrossRef]
- Wang, G.P.; Zhang, Q.W.; Ye, Z.H.; Luo, L.Z. The role of nectar plants in severe outbreaks of armyworm Mythimna separata (Lepidoptera: Noctuidae) in China. Bull. Entomol. Res. 2006, 96, 445–455. [Google Scholar] [CrossRef]
- Global Action for Fall Armyworm Control. Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/fall-armyworm/background/zh/ (accessed on 5 April 2024).
- Zhou, L.; Meng, J.Y.; Ruan, H.Y.; Yang, C.L.; Zhang, C.Y. Expression stability of candidate RT-qPCR housekeeping genes in Spodoptera frugiperda (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2021, 108, e21831. [Google Scholar] [CrossRef] [PubMed]
- Mallapur, C.P.; Naik, A.K.; Hagari, S.; Prabhu, S.T.; Patil, P.K. Status of alien pest fall armyworm, Spodoptera frugiperda (J.E. Smith) on maize in Northern Karnataka. J. Entomol. Zool. Stud. 2018, 6, 432–436. [Google Scholar]
- Zhou, Y.; Wu, Q.L.; Zhang, H.W.; Wu, K.M. Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Gong, J.; Cheng, T.C.; Wu, Y.Q.; Yang, X.; Feng, Q.L.; Mita, K. Genome-wide patterns of copy number variations in Spodoptera litura. Genomics 2019, 111, 1231–1238. [Google Scholar] [CrossRef]
- Takatsuka, J.; Okuno, S.; Nakai, M.; Kunimi, Y. Genetic and phenotypic comparisons of viral genotypes from two nucleopolyhedroviruses interacting with a common host species, Spodoptera litura (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2016, 139, 42–49. [Google Scholar] [CrossRef]
- Ming, L.; Du, Y.W.; Yuan, G.G.; Su, Q.; Shi, X.B.; Yu, H.; Chen, G. Spodoptera litura larvae are attracted by HvAV-3h-infected S. litura larvae-damaged pepper leaves. Pest Manag. Sci. 2023, 79, 2713–2724. [Google Scholar] [CrossRef]
- Xu, C.; Ji, J.C.; Zhu, X.Z.; Huangfu, N.; Xue, H.; Wang, L.; Zhang, K.X.; Li, D.Y.; Niu, L.; Chen, R.; et al. Chromosome level genome assembly of oriental armyworm Mythimna separata. Sci. Data 2023, 10, 597–608. [Google Scholar] [CrossRef]
- Li, Y.Y.; Wang, Y.N.; Zhang, H.Z.; Zhang, M.S.; Wang, M.Q.; Mao, J.J.; Zhang, L.S. The green lacewing Chrysopa formosa as a potential biocontrol agent for managing Spodoptera frugiperda and Spodoptera litura. Bull. Entomol. Res. 2023, 113, 49–62. [Google Scholar] [CrossRef]
- Tian, T.A.; Yu, L.C.; Sun, G.J.; Xiao, F.Y.; Li, L.T.; Wu, C.X.; Chen, Y.C.; Yang, M.F.; Liu, J.F. Biological control efficiency of an ectoparasitic mite Pyemotes zhonghuajia on oriental armyworm Mythimna separata. Syst. Appl. Acarol. 2020, 25, 1683–1692. [Google Scholar]
- Zettler, J.L.; Arthur, F.H. Chemical control of stored product insects with fumigants and residual treatments. Crop Prot. 2000, 19, 577–582. [Google Scholar] [CrossRef]
- Phillips, T.W.; Throne, J.E. Biorational approaches to managing stored-product insects. Annu. Rev. Entomol. 2010, 55, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Di, X.Y.; Yan, B.; Ren, P.; Wu, H.Z.; Yang, M.F. Parasitism success of Microplitis manilae (Hymenoptera: Braconidae) on different diet-fed Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae. J. Appl. Entomol. 2023, 148, 199–204. [Google Scholar] [CrossRef]
- Sarkhandia, S.; Sharma, G.; Mahajan, R.; Koundal, S.; Kumar, M.; Chadha, P.; Saini, H.S.; Kaur, S. Synergistic and additive interactions of Shewanella sp., Pseudomonas sp. and Thauera sp. with chlorantraniliprole and emamectin benzoate for controlling Spodoptera litura (Fabricius). Sci. Rep. 2023, 13, 14648. [Google Scholar] [CrossRef] [PubMed]
- Azhar, M.; Freed, S.; Sabir, H.; Rafique, S.; Naeem, A.; Ahmed, R. Effect of sub-lethal and lethal concentrations of the entomopathogenic fungus Metarhizium anisopliae Sorokin on detoxification enzymes and demographic parameters of Mythimna separata (Walker). Crop Prot. 2023, 172, 106323. [Google Scholar] [CrossRef]
- Lenteren, J.C.V.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Tang, Y.T.; Li, Y.Y.; Liu, C.X.; Mao, J.J.; Chen, H.Y.; Zhang, L.S.; Wang, M.Q. Predation and behavior of Arma chinensis to Spodoptera frugiperda. Plant Prot. 2019, 45, 65–68. [Google Scholar]
- Tang, Y.T.; Wang, M.Q.; Li, Y.Y.; Liu, C.X.; Mao, J.J.; Chen, H.Y.; Zhang, L.S. Predation of Arma chinensis on Spodoptera litura larvae. Chin. Tob. Sci. 2020, 41, 62–66. [Google Scholar]
- Pan, M.Z.; Zhang, H.P.; Zhang, L.S.; Chen, H.Y. Effects of starvation and prey availability on predation and dispersal of an omnivorous predator Arma chinensis Fallou. J. Insect Behav. 2019, 32, 134–144. [Google Scholar] [CrossRef]
- Tang, Y.T.; Wang, M.Q.; Chen, H.Y.; Wang, Y.; Zhang, H.M.; Chen, F.S.; Zhao, X.Q.; Zhang, L.S. Predatory capacity and behavior of Picromerus lewisi Scott against Spodoptera frugiperda higher instar larve. Chin. J. Biol. Control 2019, 35, 698–703. [Google Scholar]
- Yang, Q.Q.; Chen, L.; Fang, L.; Wu, Z.Y.; Ding, Y.S.; Hu, X.D.; Li, W.B.; Kong, C.X. Evaluation of predation ability of Picromerus lewisi to Spodoptera litura larvae. J. Anhui Agric. Sci. 2022, 50, 140–142. [Google Scholar]
- Tang, Y.T.; Guo, Y.; He, G.W.; Liu, C.X.; Chen, H.Y.; Zhang, L.S.; Wang, M.Q. Functional responses of Picromerus lewisi Scott (Hemiptera: Pentatomidae) attacking Mythimna separata (Walker)(Lepidoptera: Noctuidae). Chin. J. Biol. Control 2018, 34, 825–830. [Google Scholar]
- Pradeep, P.; Deshmukh, S.S.; Kalleshwaraswamy, C.M.; Rajan, S.J. Biology and predation potential of the hemipteran predator, Rhynocoris marginatus (Fab., 1794) on the fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Egypt. J. Biol. Pest Control 2022, 32, 64. [Google Scholar] [CrossRef]
- Zhang, L.S.; Chen, H.Y.; Li, B.P. Mass-Rearing and Utilization of Insect Natural Enemies, 1st ed.; Chinese Agriculture Press: Beijing, China, 2014; pp. 119–122. [Google Scholar]
- Zhao, P.; Yuan, J.L. The insect list and faunal analysis of Harpactorinae in Guizhou Province. Guizhou Agric. Sci. 2011, 39, 99–102. [Google Scholar]
- Tomson, M. Functional response, host stage preference and development of Rhynocoris fuscipes (Fab.) (Heteroptera: Reduviidae) for two cotton pests. J. Biopestic. 2021, 14, 12–21. [Google Scholar] [CrossRef]
- Sunil, V.; Sampathkumar, M.; Lydia, C.; Chiranjeevi, K.; Shanker, C. Biology, predatory potential and functional response of Rhynocoris fuscipes (Fabricius) (Hemiptera: Reduviidae) on rice brown planthopper, Nilaparvata lugens (Stal) (Homoptera: Delphacidae). J. Exp. Zool. India 2018, 21, 259–263. [Google Scholar]
- Tomson, M.; Sahayaraj, K.; Kumar, V.; Avery, P.B.; McKenzie, C.L.; Osborne, L.S. Mass rearing and augmentative biological control evaluation of Rhynocoris fuscipes (Hemiptera: Reduviidae) against multiple pests of cotton. Pest Manag. Sci. 2017, 73, 1743–1752. [Google Scholar] [CrossRef]
- Deng, H.B.; Wang, Z.; Chen, Y.M.; Wu, W.B.; Peng, W.S. Predation of Harpactor fuscipes on Helicoverpa assulta and Spodoptera litura. Guangdong Agric. Sci. 2012, 39, 107–109. [Google Scholar]
- Solomon, M.E. The natural control of animal populations. J. Anim. Ecol. 1949, 18, 1–35. [Google Scholar] [CrossRef]
- Poncio, S.; Montoya, P.; Cancino, J.; Nava, D.E. Determining the Functional Response and Mutual Interference of Utetes anastrephae (Hymenoptera: Braconidae) on Anastrepha obliqua (Diptera: Tephritidae) Larvae for Mass Rearing Purposes. Ann. Entomol. Soc. Am. 2016, 109, 518–525. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Dick, J.T.A.; Callaghan, A.; Dickey, J.W.E. Biological control agent selection under environmental change using functional responses, abundances and fecundities; the relative control potential (RCP) metric. Biol. Control 2018, 121, 50–57. [Google Scholar] [CrossRef]
- Ingegno, B.L.; Messelink, G.J.; Bodino, N.; Lliadou, A.; Driss, L.; Woelke, J.B.; Leman, A.; Tavella, L. Functional response of the mirid predators Dicyphus bolivari and Dicyphus errans and their efficacy as biological control agents of Tuta absoluta on tomato. J. Pest Sci. 2019, 92, 1457–1466. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Juliano, S.A. Nonlinear Curve Fitting: Predation and Functional Response Curves, 2nd ed.; Chapman and Hall: London, UK, 2001; pp. 178–196. [Google Scholar]
- Uiterwaal, S.F.; Lagerstrom, I.T.; Lyon, S.R.; Delong, J.P. Data paper: FoRAGE (Functional Responses from Around the Globe in all Ecosystems) database: A compilation of functional responses for consumers and parasitoids. BioRxiv 2018. [Google Scholar] [CrossRef]
- Messina, F.J.; Hanks, J.B. Host plant alters the shape of the functional response of an aphid predator (Coleoptera: Coccinellidae). Environ. Entomol. 1998, 27, 1196–1202. [Google Scholar] [CrossRef]
- Hassanzadeh-Avval, M.; Sadeghi-Namaghi, H.; Fekrat, L. Factors influencing functional response, handling time and searching efficiency of Anthocoris minki Dohrn (Hem.: Anthocoridae) as predator of Psyllopsis repens Loginova (Hem.: Psyllidae). Phytoparasitica 2019, 47, 341–350. [Google Scholar] [CrossRef]
- Hassanpour, M.; Mohaghegh, J.; Iranipour, S.; Nouri-Ganbalani, G.; Enkegaard, A. Functional response of Chrysoperla carnea (Neuroptera: Chrysopidae) to Helicoverpa armigera (Lepidoptera: Noctuidae): Effect of prey and predator stages. Insect Sci. 2011, 18, 217–224. [Google Scholar] [CrossRef]
- Ziaei Madbouni, M.A.; Samih, M.A.; Namvar, P.; Biondi, A. Temperature-dependent functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to different densities of pupae of cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Eur. J. Entomol. 2017, 114, 325–331. [Google Scholar] [CrossRef]
- Pakyari, H.; Kasirloo, F.; Arbab, A. Effect of sublethal doses of Abamectin and fenpropathrin on functional response of Cryptolaemus Montrouzieri (Coleoptera: Coccinellidae) predator of Planococcus citri (Hemiptera: Pseudococcidae). J. Entomol. Zool. Stud. 2016, 4, 469–473. [Google Scholar]
- Hassell, M.P. A population model for the interaction between Cyzenisal bicans (Fall.) (Techinidae) and Opero phterabrumata (L.) (Geometridae) at Wytham, Berkshire. J. Anim. Ecol. 1969, 38, 567–576. [Google Scholar] [CrossRef]
- Hassanzadeh-Avval, M.; Sadeghi-Namaghi, H.; Fekrat, L. Prey preference and prey switching in Anthocoris minki Dohrn (Hemiptera: Anthocoridae). J. Asia-Pac. Entomol. 2018, 21, 1116–1121. [Google Scholar] [CrossRef]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Chen, W.B.; Li, Y.Y.; Wang, M.Q.; Mao, J.J.; Zhang, L.S. Evaluating the potential of using Spodoptera litura eggs for mass-rearing Telenomus remus, a promising egg parasitoid of Spodoptera frugiperda. Insects 2021, 12, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D. Random search and insect population models. J. Anim. Ecol. 1972, 41, 369–383. [Google Scholar] [CrossRef]
- Hassell, M.P.; Varley, G.C. New inductive population model for insect parasites and its bearing on biological control. Nature 1969, 223, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Lechowicz, M.J. The sampling characteristics of electivity indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Dickey, J.W.E.; Coughlan, N.E.; Joyce, P.W.S.; Dick, J.T.A. The functional response ratio (FRR): Advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biol. Invasions 2019, 21, 2543–2547. [Google Scholar] [CrossRef]
- George, P.J.E.; Seenivasagan, R. Predatory Efficiency of Rhynocoris marginatus (Fabricius)(Heteroptera: Reduviidae) on Helicoverpa armigera (Hiibner) and Spodoptera litura (Fabricius). J. Biol. Control 1998, 12, 25–29. [Google Scholar]
- Feng, Y.; Zhou, Z.X.; An, M.R.; Yu, X.L.; Liu, T.X. The effects of prey distribution and digestion on functional response of Harmonia axyridis (Coleoptera: Coccinellidae). Biol. Control 2018, 124, 74–81. [Google Scholar] [CrossRef]
- Varshney, R.; Budhlakoti, N.; Ballal, C.R. Functional response of Geocoris ochropterus Fieber (Hemiptera: Geocoridae) to different egg densities of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Phytoparasitica 2018, 46, 451–458. [Google Scholar] [CrossRef]
- Liu, P.; Jia, W.; Zheng, X.; Zhang, L.; Sangbaramou, R.; Tan, S.Q.; Liu, Y.Q.; Shi, W.P. Predation functional response and life table parameters of Orius sauteri (Hemiptera: Anthocoridae) feeding on Megalurothrips usitatus (Thysanoptera: Thripidae). Fla. Entomol. 2018, 101, 254–259. [Google Scholar] [CrossRef]
- Nagarajan, K.; Rajan, K.; Ambrose, D.P. Functional response of assassin bug, Rhynocoris fuscipes (Fabricius) (Hemiptera: Reduviidae) to cucumber leaf folder, Diaphania indicus Saunders (Lepidoptera: Pyraustidae). Entomon 2010, 35, 1–7. [Google Scholar]
- Ambrose, D.P.; Nagarajan, K. Functional response of Rhynocoris fuscipes (Fabricius) (Hemiptera: Reduviidae) to teak skeletonizer Eutectona machaeralis Walker (Lepidoptera: Pyralidae). J. Biol. Control 2010, 24, 175–178. [Google Scholar]
- Stasek, D.J.; Radl, J.N.; Crist, T.O. The functional response and prey preference of generalist Nabis (Hemiptera: Nabidae) predators to leafhopper prey (Hemiptera: Cicadellidae). Can. Entomol. 2018, 150, 190–200. [Google Scholar] [CrossRef]
- Sajjad, S.; Sultan, A.; Khan, M.F.; Keerio, I.D.; Channa, M.S.; Akbar, M.F. Biology, life table parameters, and functional response of Chrysoperla carnea (Neuroptera: Chrysopidae) on different stages of invasive Paracoccus marginatus (Hemiptera: Pseudococcidae). J. Asia-Pac. Biodivers. 2021, 14, 174–182. [Google Scholar] [CrossRef]
- Mohaghegh, J.; Clercq, P.D.; Tirry, L. Functional response of the predators Podisus maculiventris (Say) and Podisus nigrispinus (Dallas) (Het., Pentatomidae) to the beet armyworm, Spodoptera exigua (Hübner) (Lep., Noctuidae): Ef fect of temperature. J. Appl. Entomol. 2001, 125, 131–134. [Google Scholar] [CrossRef]
- Hassanpour, M.; Maghami, R.; Rafiee-Dastjerdi, H.; Golizadeh, A.; Yazdanian, M.; Enkegaard, A. Predation activity of Chrysoperla carnea (Neuroptera: Chrysopidae) upon Aphis fabae (Hemiptera: Aphididae): Effect of different hunger levels. J. Asia-Pac. Entomol. 2015, 18, 297–302. [Google Scholar] [CrossRef]
- Schenk, D.; Bacher, S. Functional response of a generalist insect predator to one of its prey species in the field. J. Anim. Ecol. 2002, 71, 524–531. [Google Scholar] [CrossRef]
- Khan, M.A.Z.; Liang, Q.; Liu, T.X. Effect of cage size on functional response of the parasitoid Aphidius Gifuensis (Ashmead) (Hymenoptera: Braconidae) against Myzus Persicae (Sulzer) (Hemiptera: Aphididae). Egypt. J. Biol. Pest Control 2016, 26, 373. [Google Scholar]
- Kalinkat, G.; Rall, B.C.; Uiterwaal, S.F.; Uszko, W. Empirical evidence of type III functional responses and why it remains rare. Front. Ecol. Evol. 2023, 11, 1033818. [Google Scholar] [CrossRef]
- Viteri Jumbo, L.O.; Teodoro, A.V.; Rego, A.S.; Haddi, K.; Galvao, A.S.; de Oliveira, E.E. The lacewing Ceraeochrysa caligata as a potential biological agent for controlling the red palm mite Raoiella indica. PeerJ 2019, 7, e7123. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Kalidas, S.; Tomson, M. Stage preference and functional response of Rhynocoris longifrons (Stål) (Hemiptera: Reduviidae) on three hemipteran cotton pests. Braz. Arch. Biol. Technol. 2012, 55, 733–740. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Kumar, V.; Avery, P.B. Functional response of Rhynocoris kumarii (Hemiptera: Reduviidae) to different population densities of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) recorded in the laboratory. Eur. J. Entomol. 2015, 112, 69–74. [Google Scholar] [CrossRef]
- Atrchian, H.; Mahdian, K.; Shahidi Noghabi, S. Effect of temperature on predation of Chilocorus bipustulatus L. (Col.: Coccinellidae) feeding on Agonoscena pistaciae (Hem.: Psyllidae). Phytoparasitica 2023, 51, 215–226. [Google Scholar]
- Mondal, R.P.; Chandra, G.; Bandyopadhyay, S.; Ghosh, A. Effect of temperature and search area on the functional response of Anisops sardea (Hemiptera: Notonectidae) against Anopheles stephensi in laboratory bioassay. Acta Trop. 2017, 166, 262–267. [Google Scholar] [CrossRef]
- Gholami, N.; Aleosfoor, M.; Hosseini, M.; Fekrat, L. Predation functional response and demographic parameters of Orius albidipennis (Hemiptera: Anthocoridae) on Schizaphis graminum (Hemiptera: Aphididae): Effect of host plant morphological attributes. Biocontrol Sci. Technol. 2022, 32, 362–380. [Google Scholar] [CrossRef]
- Maselou, D.; Perdikis, D.; Fantinou, A. Effect of hunger level on prey consumption and functional response of the predator Macrolophus pygmaeus. Bull. Insectology 2015, 68, 211–218. [Google Scholar]
- Ambrose, D.P.; Rajan, S.J.; Raja, J.M. Impacts of Synergy-505 on the functional response and behavior of the reduviid bug, Rhynocoris marginatus. J. Insect Sci. 2010, 10, 187. [Google Scholar] [CrossRef]
- Zhang, D.J.; Maiga, H.; Li, Y.J.; Bakhoum, M.T.; Wang, G.; Sun, Y.; Damiens, D.; Mamai, W.; Somda, N.S.B.; Wallner, T.; et al. Mating harassment may boost the effectiveness of the sterile insect technique for Aedes mosquitoes. Nat. Commun. 2024, 15, 1980. [Google Scholar] [CrossRef]
- Wu, P.X.; He, J.; Dong, H.; Zhang, R.Z. Functional response and intraspecific competition of three ladybird species feeding on aphids on goji berry plants in laboratory and semi-field conditions. Insects 2023, 14, 853. [Google Scholar] [CrossRef]
- Wu, P.X.; Zhang, J.; Haseeb, M.; Yan, S.; Kanga, L.; Zhang, R.Z. Functional responses and intraspecific competition in the ladybird Harmonia axyridis (Coleoptera: Coccinellidae) provided with Melanaphis sacchari (Homoptera: Aphididae) as prey. Eur. J. Entomol. 2018, 115, 232–241. [Google Scholar] [CrossRef]
- Nachappa, P.; Braman, S.K.; Guillebeau, L.P.; All, J.N. Functional response of the tiger beetle Megacephala carolina carolina (Coleoptera: Carabidae) on twolined spittlebug (Hemiptera: Cercopidae) and fall armyworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 2006, 99, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Messelink, G.J.; Vijverberg, R.; Leman, A.; Janssen, A. Biological control of mealybugs with lacewing larvae is affected by the presence and type of supplemental prey. BioControl 2016, 61, 555–565. [Google Scholar] [CrossRef][Green Version]
- Nagarajan, K.; Ambrose, D.P. Chemically mediated prey-approaching behaviour of the reduviid predator Rhynocoris fuscipes (Fabricius) (Insecta: Heteroptera: Reduviidae) by Y-arm olfactometer. Pak. J. Biol. Sci. 2013, 16, 1363–1367. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.X.; Enkegaard, A. Predation capacity and prey preference of Chrysoperla carnea on Pieris brassicae. BioControl 2010, 55, 379–385. [Google Scholar] [CrossRef]
- Daniel, S.; Kumar, S.P. Impact of prey types on the predatory behaviour of Rhynocoris fuscipes (Insecta: Heteroptera). J. Ecobiol. 2003, 15, 121–126. [Google Scholar]
- Sujatha, S.; Vidya, L.S.; Sumi, G.S. Prey-predator interaction and info-chemical behavior of Rhynocoris fuscipes (Fab.) on three agricultural pests (Heteroptera: Reduviidae). J. Entomol. 2012, 9, 130–136. [Google Scholar] [CrossRef]
- Kumar, R.M.; Gadratagi, B.G.; Paramesh, V.; Kumar, P.; Madivalar, Y.; Narayanappa, N.; Ullah, F. Sustainable management of invasive fall armyworm, Spodoptera frugiperda. Agronomy 2022, 12, 2150. [Google Scholar] [CrossRef]
- Thakur, N.; Sharma, A.; Kaur, S.; Ahluwalia, K.K.; Sidhu, A.K.; Kumar, S.; Rustagi, S.; Singh, S.; Rai, A.K.; Sheikh, S. Insect pest Spodoptera litura (Fabricius) and its resistance against the chemical insecticides: A review. Plant Sci. Today 2024, 11, 192–203. [Google Scholar] [CrossRef]
- Xu, Y.C.; Yang, C.H.; Li, Y.H.; Shi, C.J.; He, Y.G.; Yang, M.H.; Zhang, Y.; Tang, G.W. Larvae head morphology and ultrastructure of sensilla in tyrannophagous Mythimna separata and Spodoptera litura. Plant Prot. 2024, 50, 196–204. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).