
Age and Density of Mated Females Affect Dispersal Strategies in Spider Mite Tetranychus ludeni Zacher



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mite Colony and Experimental Conditions
2.2. Preparation of Mated Females for Experiments
2.3. Dispersal and Reproduction of Females of Different Ages
2.4. Dispersal of Females at Different Densities
2.5. Statistical Analysis
3. Results
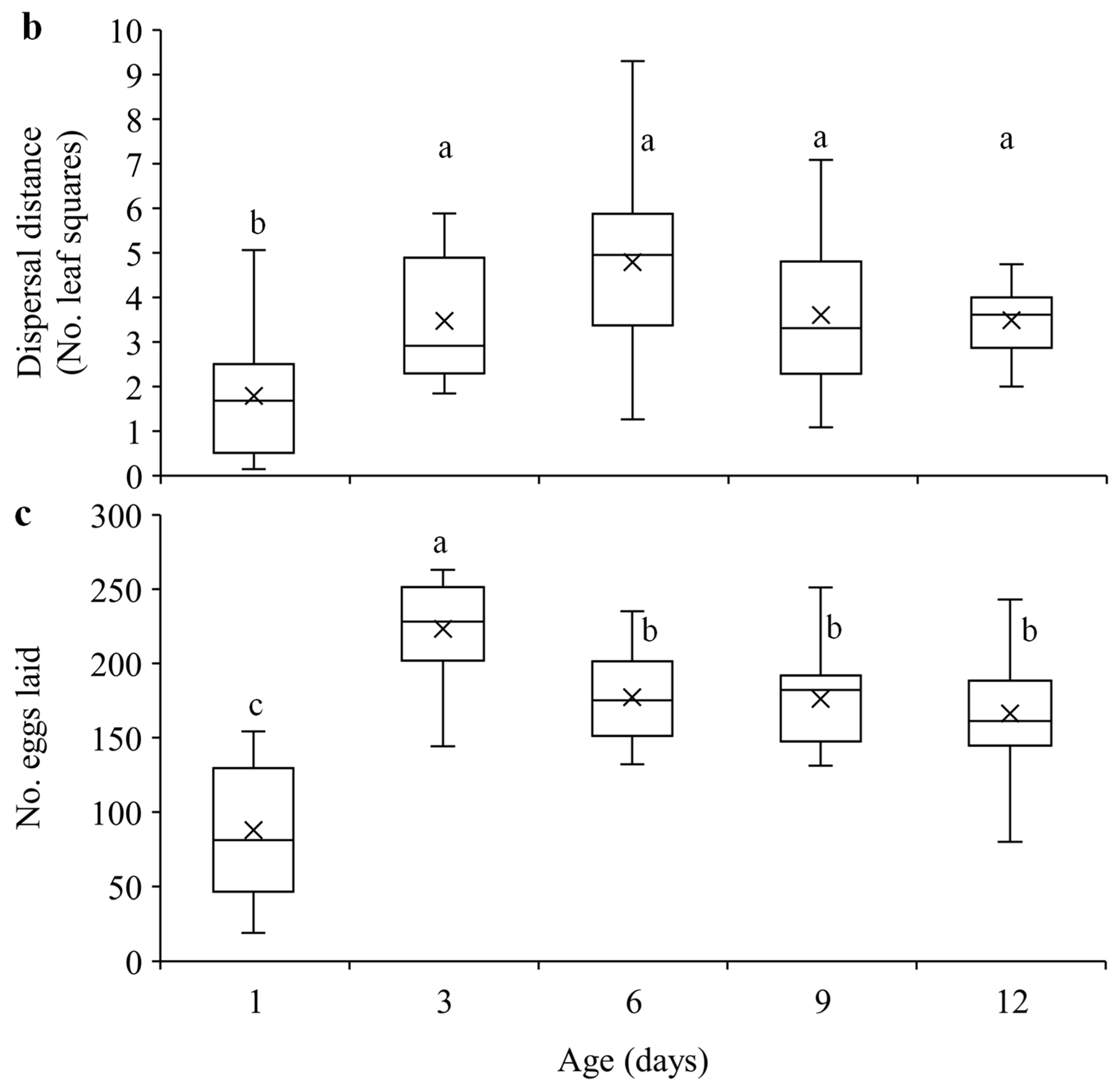
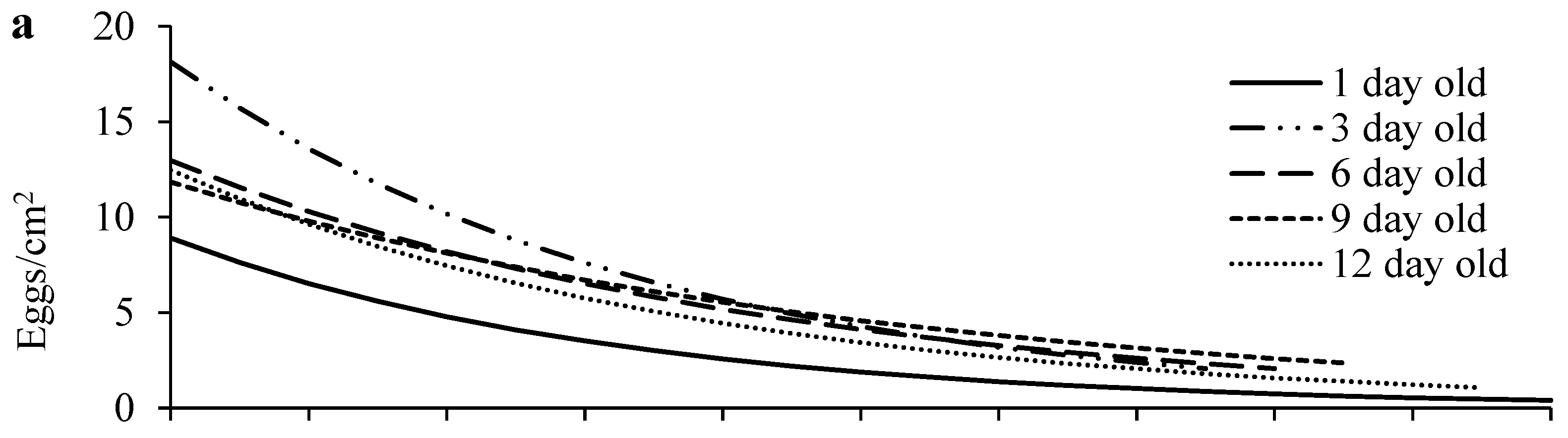
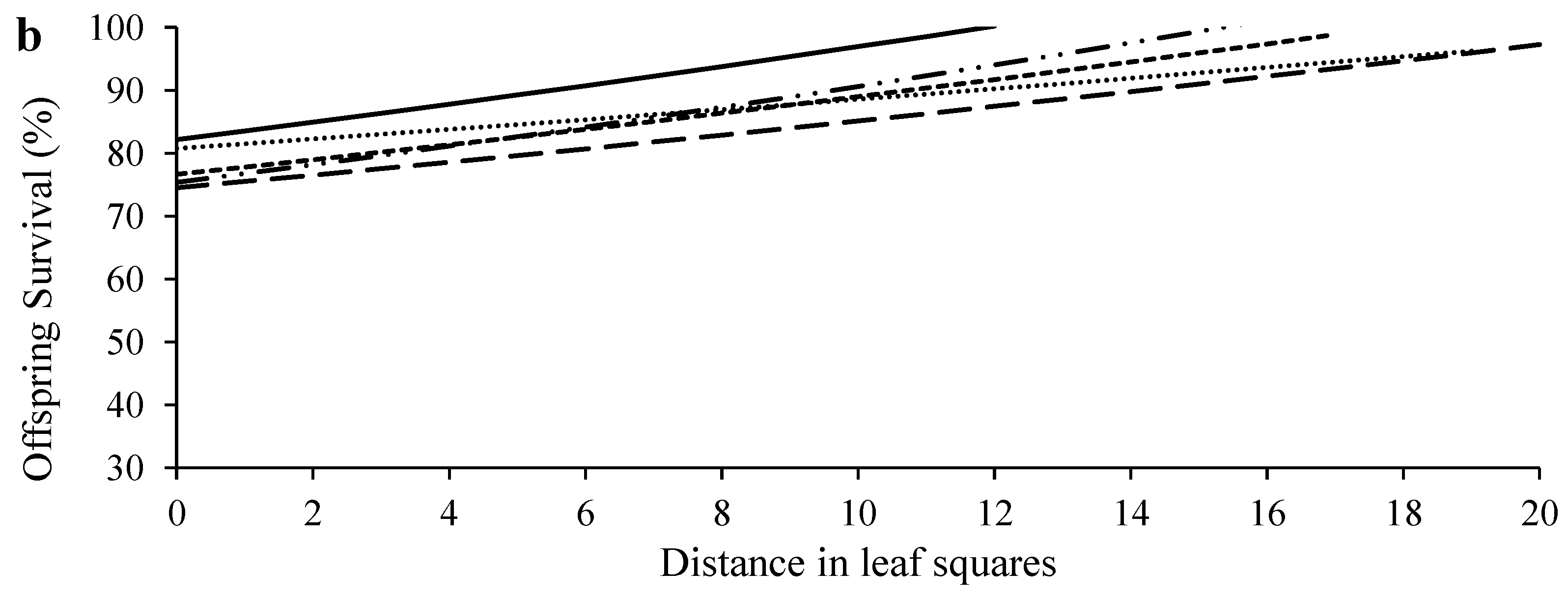
3.1. Dispersal and Reproduction of Females of Different Ages
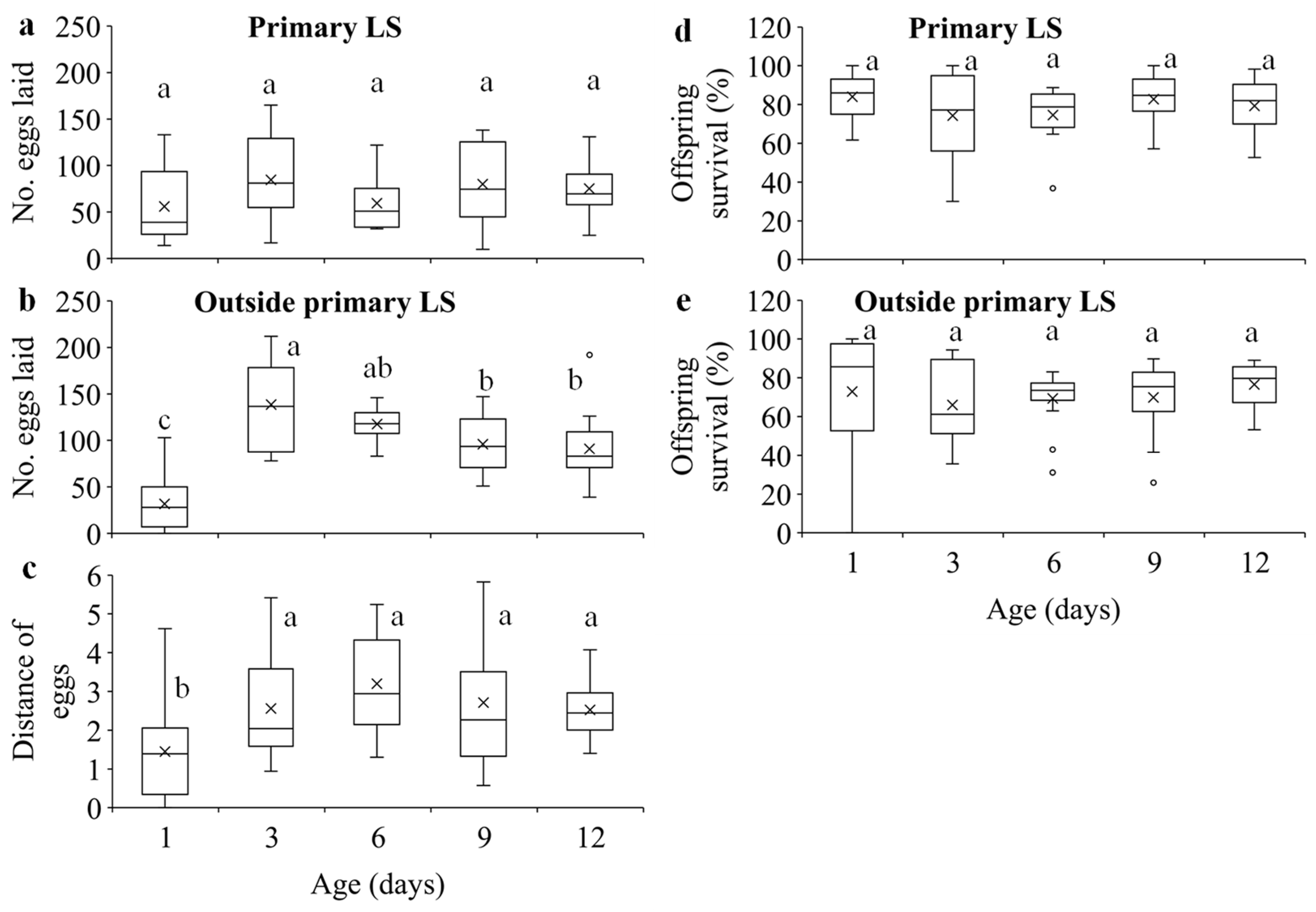
3.2. Dispersal of Females at Different Densities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthysen, E. Multicausality of dispersal: A review. In Dispersal Ecology and Evolution; Clobert, J., Baguette, M., Benton, T.G., Bullock, J.M., Eds.; Oxford University Press: London, UK, 2012; Volume 27, pp. 3–18. [Google Scholar]
- Azandémè-Hounmalon, G.Y.; Fellous, S.; Kreiter, S.; Fiaboe, K.K.; Subramanian, S.; Kungu, M.; Martin, T. Dispersal behavior of Tetranychus evansi and T. urticae on tomato at several spatial scales and densities: Implications for integrated pest management. PLoS ONE 2014, 9, e95071. [Google Scholar] [CrossRef] [PubMed]
- Bowler, D.E.; Benton, T.G. Causes and consequences of animal dispersal strategies: Relating individual behaviour to spatial dynamics. Biol. Rev. Camb. Philos. Soc. 2005, 80, 205–225. [Google Scholar] [CrossRef]
- Clobert, J.; Le Galliard, J.F.; Cote, J.; Meylan, S.; Massot, M. Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecol. Lett. 2009, 12, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Regniere, J.; Nealis, V.G. Density dependence of egg recruitment and moth dispersal in spruce budworms. Forests 2019, 10, 706. [Google Scholar] [CrossRef]
- Travis, J.M.J.; Mustin, K.; Benton, T.G.; Dytham, C. Accelerating invasion rates result from the evolution of density-dependent dispersal. J. Theor. Biol. 2009, 259, 151–158. [Google Scholar] [CrossRef]
- Hovestadt, T.; Mitesser, O.; Poethke, H.-J. Gender-specific emigration decisions sensitive to local male and female density. Am. Nat. 2014, 184, 38–51. [Google Scholar] [CrossRef]
- Hovestadt, T.; Poethke, H.J. The control of emigration and its consequences for the survival of populations. Ecol. Modell. 2006, 190, 443–453. [Google Scholar] [CrossRef]
- Li, J.; Margolies, D.C. Effects of mite age, mite density, and host quality on aerial dispersal behavior in the twospotted spider mite. Entomol. Exp. Appl. 1993, 68, 79–86. [Google Scholar] [CrossRef]
- Mishra, A.; Tung, S.; Sruti, V.R.S.; Sadiq, M.A.; Srivathsa, S.; Dey, S. Pre-dispersal context and presence of opposite sex modulate density dependence and sex bias of dispersal. Oikos 2018, 127, 1596–1604. [Google Scholar] [CrossRef]
- Plazio, E.; Margol, T.; Nowicki, P. Intersexual differences in density-dependent dispersal and their evolutionary drivers. J. Evol. Biol. 2020, 33, 1495–1506. [Google Scholar] [CrossRef]
- Suiter, K.A.; Gould, F. Effects of mating status and age on dispersal behavior in the twospotted spider mite, Tetranychus urticae in response to fenvalerate-treated leaf surfaces. Entomol. Exp. Appl. 1992, 62, 1–8. [Google Scholar] [CrossRef]
- Fadamiro, H.Y. Free flight capacity determination in a sustained flight tunnel: Effects of age and sexual state on the flight duration of Prostephanus truncatus. Physiol. Entomol. 1997, 22, 29–36. [Google Scholar] [CrossRef]
- Gu, H.; Danthanarayana, W. Age-related flight and reproductive performance of the light brown apple moth, Epiphyas postvittana. Entomol. Exp. Appl. 1990, 54, 109–115. [Google Scholar] [CrossRef]
- Ishizaki, M.; Watanabe, T.; Moriya, S.; Tabuchi, K. Diurnal locomotion activity of adult rice bug, Leptocorisa chinensis (Hemiptera: Alydidae), at different ages, measured by actograph and video camera. Appl. Entomol. Zool. 2011, 46, 135–142. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Campbell, J.F.; Throne, J.E. Influence of age, mating status, sex, quantity of food, and long-term food deprivation on red flour beetle (Coleoptera: Tenebrionidae) flight initiation. J. Econ. Entomol. 2011, 104, 2078–2086. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Bowler, D.E. Dispersal in invertebrates: Influences on individual decisions. In Dispersal Ecology and Evolution; Clobert, J., Baguette, M., Benton, T.G., Bullock, J.M., Eds.; Oxford University Press: London, UK, 2012; pp. 41–49. [Google Scholar]
- Lakovic, M.; Poethke, H.-J.; Hovestadt, T. Dispersal timing: Emigration of insects living in patchy environments. PLoS ONE 2015, 10, e0128672. [Google Scholar] [CrossRef]
- Nowicki, P.; Vrabec, V. Evidence for positive density-dependent emigration in butterfly metapopulations. Oecologia 2011, 167, 657–665. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Wilson, M.L. Culex restuans (Diptera: Culicidae) oviposition behavior determined by larval habitat quality and quantity in southeastern Michigan. J. Med. Entomol. 2004, 41, 179–186. [Google Scholar] [CrossRef]
- Salomão, R.P.; Arellano, L.; Huerta, C.; León-Cortés, J.L. Do sexual gonadic maturity and age determine habitat occupancy of Canthon cyanellus LeConte, 1859 (Coleoptera: Scarabaeidae)? Can. Entomol. 2021, 153, 412–427. [Google Scholar] [CrossRef]
- Dingle, H. Relation between age and flight activity in milkweed bug, Oncopeltus. J. Exp. Biol. 1965, 42, 269–283. [Google Scholar] [CrossRef]
- Järemo, J.; Bengtsson, G. On the importance of life history and age structure in biological invasions. Ecol. Modell. 2011, 222, 485–492. [Google Scholar] [CrossRef]
- Schumacher, P.; Weyeneth, A.; Weber, D.C.; Dorn, S. Long flights in Cydia pomonella L. (Lepidoptera: Tortricidae) measured by a flight mill: Influence of sex, mated status and age. Physiol. Entomol. 1997, 22, 149–160. [Google Scholar] [CrossRef]
- Stewart, S.D.; Gaylor, M.J. Age, sex, and reproductive status of the tarnished plant bug (Heteroptera: Miridae) colonizing mustard. Environ. Entomol. 1991, 20, 1387–1392. [Google Scholar] [CrossRef]
- Williamson, J.A.; Charlesworth, B. The effect of age of founder on the probability of survival of a colony. J. Theor. Biol. 1976, 56, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Bonte, D.; De Roissart, A.; Wybouw, N.; Van Leeuwen, T. Fitness maximization by dispersal: Evidence from an invasion experiment. Ecology 2014, 95, 3104–3111. [Google Scholar] [CrossRef]
- Burgess, S.C.; Powell, J.; Bueno, M. Dispersal, kin aggregation, and the fitness consequences of not spreading sibling larvae. Ecology 2022, 104, e3858. [Google Scholar] [CrossRef] [PubMed]
- De Meester, N.; Bonte, D. Information use and density-dependent emigration in an agrobiont spider. Behav. Ecol. 2010, 21, 992–998. [Google Scholar] [CrossRef]
- Bitume, E.V.; Bonte, D.; Ronce, O.; Bach, F.; Flaven, E.; Olivieri, I.; Nieberding, C.M. Density and genetic relatedness increase dispersal distance in a subsocial organism. Ecol. Lett. 2013, 16, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Poethke, H.J.; Gros, A.; Hovestadt, T. The ability of individuals to assess population density influences the evolution of emigration propensity and dispersal distance. J. Theor. Biol. 2011, 282, 93–99. [Google Scholar] [CrossRef]
- Ventura, L.; Smith, D.R.; Lubin, Y. Crowding leads to fitness benefits and reduced dispersal in a colonial spider. Behav. Ecol. 2017, 28, 1384–1392. [Google Scholar] [CrossRef]
- Bolland, H.R.; Gutierrez, J.; Flechtmann, C.H. World Catalogue of the Spider Mite Family (Acari: Tetranychidae); Brill: Leiden, The Netherlands, 1998; p. 392. [Google Scholar]
- CABI. Tetranychus ludeni (Red Spider Mite). Available online: https://www.cabi.org/isc/datasheet/53351 (accessed on 6 April 2024).
- Zhou, P.; He, X.Z.; Wang, Q. Sons from virgin mothers produce more daughters in a haplodiploid mite. Syst. Appl. Acarol. 2018, 23, 1869–1878. [Google Scholar] [CrossRef]
- Zhou, P.; He, X.Z.; Chen, C.; Wang, Q. Resource relocations in relation to dispersal in Tetranychus ludeni Zacher. Syst. Appl. Acarol. 2021, 26, 2018–2026. [Google Scholar] [CrossRef]
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider mites web: A comprehensive database for the Tetranychidae. In Trends in Acarology; Sabelis, M.W., Bruin, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 557–560. [Google Scholar]
- Ragusa, E.; Sinacori, M.; Tsolakis, H. First record of Tetranychus ludeni Zacher (Acariformes: Tetranychidae) in Italy. Int. J. Acarol. 2019, 45, 26–28. [Google Scholar] [CrossRef]
- Zhang, Z.-Q. Mites of Greenhouses: Identification, Biology and Control; CABI Publishing: Wallingford, UK, 2003; p. 244. [Google Scholar]
- Mcenroe, W.D. Spreading and inbreeding in the spider mite. J. Hered. 1969, 60, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Smitley, D.R.; Kennedy, G.G. Aerial dispersal of the two-spotted spider mite (Tetranychus urticae) from field corn. Exp. Appl. Acarol. 1988, 5, 33–46. [Google Scholar] [CrossRef]
- Weerawansha, N.; Wang, Q.; He, X.Z. Effect of foundress population density and size on reproduction and population growth of a haplodiploid mite. Syst. Appl. Acarol. 2020, 25, 2063–2076. [Google Scholar]
- Weerawansha, N.; Wang, Q.; He, X.Z. A haplodiploid mite adjusts fecundity and sex ratio in response to density changes during the reproductive period. Exp. Appl. Acarol. 2022, 88, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Weerawansha, N.; Wang, Q.; He, X.Z. Adjustment of fecundity and sex ratio in response to social environments in a haplodiploid mite. Syst. Appl. Acarol. 2022, 27, 61–70. [Google Scholar] [CrossRef]
- Mitchell, R. Growth and population dynamics of a spider mite (Tetranychus urticae K., Acarina: Tetranychidae). Ecology 1973, 54, 1349–1355. [Google Scholar] [CrossRef]
- Geroh, M.; Gulati, R.; Sharma, S.; Kaushik, H.; Aneja, D. Effect of abiotic stresses on population build up of Tetranychus urticae Koch and its predator, Stethorus punctillum Weise on okra. Ann. Plant Prot. Sci. 2010, 18, 108–113. [Google Scholar]
- Helle, W.; Sabelis, M.W. Spider Mites: Their Biology, Natural Enemies and Control; Elsevier: Amsterdam, The Netherlands, 1985; p. 458. [Google Scholar]
- Adango, E.; Onzo, A.; Hanna, R.; Atachi, P.; James, B. Comparative demography of the spider mite, Tetranychus ludeni, on two host plants in West Africa. J. Insect Sci. 2006, 6, 49. [Google Scholar] [CrossRef]
- Zhou, P.; He, X.Z.; Chen, C.; Wang, Q. Reproductive strategies that may facilitate invasion success: Evidence from a spider mite. J. Econ. Entomol. 2021, 114, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.D.; Gaylor, M.J. Effects of age, sex, and reproductive status on flight by the tarnished plant bug (Heteroptera: Miridae). Environ. Entomol. 1994, 23, 80–84. [Google Scholar] [CrossRef]
- Bonte, D.; Dahirel, M. Dispersal: A central and independent trait in life history. Oikos 2017, 126, 472–479. [Google Scholar] [CrossRef]
- Buoro, M.; Carlson, S.M. Life-history syndromes: Integrating dispersal through space and time. Ecol. Lett. 2014, 17, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Kokko, H. Intersexual resource competition and the evolution of sex-biased dispersal. Front. Ecol. Evol. 2019, 7, 111. [Google Scholar] [CrossRef]
- Brandenburg, R.L.; Kennedy, G.G. Intercrop relationships and spider mite dispersal in a corn/peanut agro-ecosystem. Entomol. Exp. Appl. 1982, 32, 269–276. [Google Scholar] [CrossRef]
- Nadeali, T.; Golpayegani, A.Z.; Saboori, A. When do the predators leave their patch? Leaving tendency in Phytoseiulus persimilis and Neoseiulus californicus (Phytoseiidae). Syst. Appl. Acarol. 2014, 19, 263–274. [Google Scholar]
- Vanas, V.; Enigl, M.; Walzer, A.; Schausberger, P. The predatory mite Phytoseiulus persimilis adjusts patch-leaving to own and progeny prey needs. Exp. Appl. Acarol. 2006, 39, 1–11. [Google Scholar] [CrossRef]
- Schausberger, P.; Yano, S.; Sato, Y. Cooperative behaviors in group-living spider mites. Front. Ecol. Evol. 2021, 9, 745036. [Google Scholar] [CrossRef]
- Denno, R.F.; Roderick, G.K. Density-related dispersal in planthoppers-effects of interspecific crowding. Ecology 1992, 73, 1323–1334. [Google Scholar] [CrossRef]
- Li, G.-Y.; Zhang, Z.-Q. The costs of social interaction on survival and reproduction of arrhenotokous spider mite Tetranychus urticae. Entomol. Gen. 2021, 41, 49–57. [Google Scholar] [CrossRef]
- Dahirel, M.; Vardakis, M.; Ansart, A.; Madec, L. Density-dependence across dispersal stages in a hermaphrodite land snail: Insights from discrete choice models. Oecologia 2016, 181, 1117–1128. [Google Scholar] [CrossRef]
- Sæther, B.-E.; Engen, S.; Lande, R. Finite metapopulation models with density-dependent migration and stochastic local dynamics. Proc. R. Soc. Lond. B 1999, 266, 113–118. [Google Scholar] [CrossRef]
- Zhou, P.; He, X.Z.; Chen, C.; Wang, Q. Effect of age and density on dispersal probability and distance in Tetranychus ludeni Zacher. Zoosymposia 2022, 22, 122. [Google Scholar] [CrossRef]
- Bowler, D.E.; Benton, T.G. Variation in dispersal mortality and dispersal propensity among individuals: The effects of age, sex and resource availability. J. Anim. Ecol. 2009, 78, 1234–1241. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, P.; He, X.Z.; Chen, C.; Wang, Q. Age and Density of Mated Females Affect Dispersal Strategies in Spider Mite Tetranychus ludeni Zacher. Insects 2024, 15, 387. https://doi.org/10.3390/insects15060387
Zhou P, He XZ, Chen C, Wang Q. Age and Density of Mated Females Affect Dispersal Strategies in Spider Mite Tetranychus ludeni Zacher. Insects. 2024; 15(6):387. https://doi.org/10.3390/insects15060387
Chicago/Turabian StyleZhou, Peng, Xiong Z. He, Chen Chen, and Qiao Wang. 2024. "Age and Density of Mated Females Affect Dispersal Strategies in Spider Mite Tetranychus ludeni Zacher" Insects 15, no. 6: 387. https://doi.org/10.3390/insects15060387
APA StyleZhou, P., He, X. Z., Chen, C., & Wang, Q. (2024). Age and Density of Mated Females Affect Dispersal Strategies in Spider Mite Tetranychus ludeni Zacher. Insects, 15(6), 387. https://doi.org/10.3390/insects15060387