

Topical Toxicity and Repellency Profiles of 17 Essential Oil Components against Insecticide-Resistant and Susceptible Strains of Adult Musca domestica (Diptera: Muscidae)




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. House Flies
2.3. Topical Application
2.3.1. Resistant House Fly Strain
2.3.2. Susceptible House Fly Strain
2.4. Repellency Bioassay
2.5. Data Analysis
3. Results
3.1. Topical Application
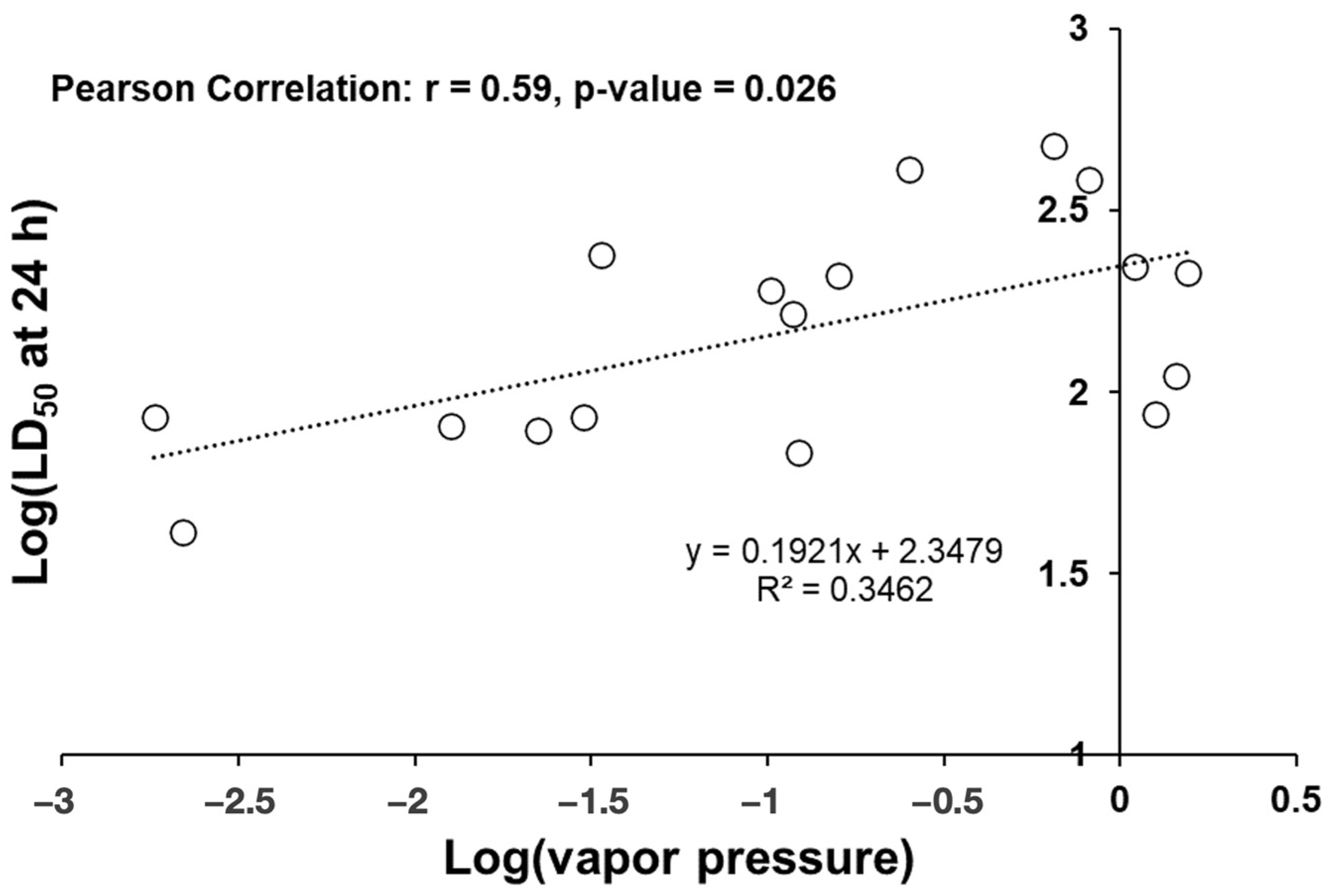
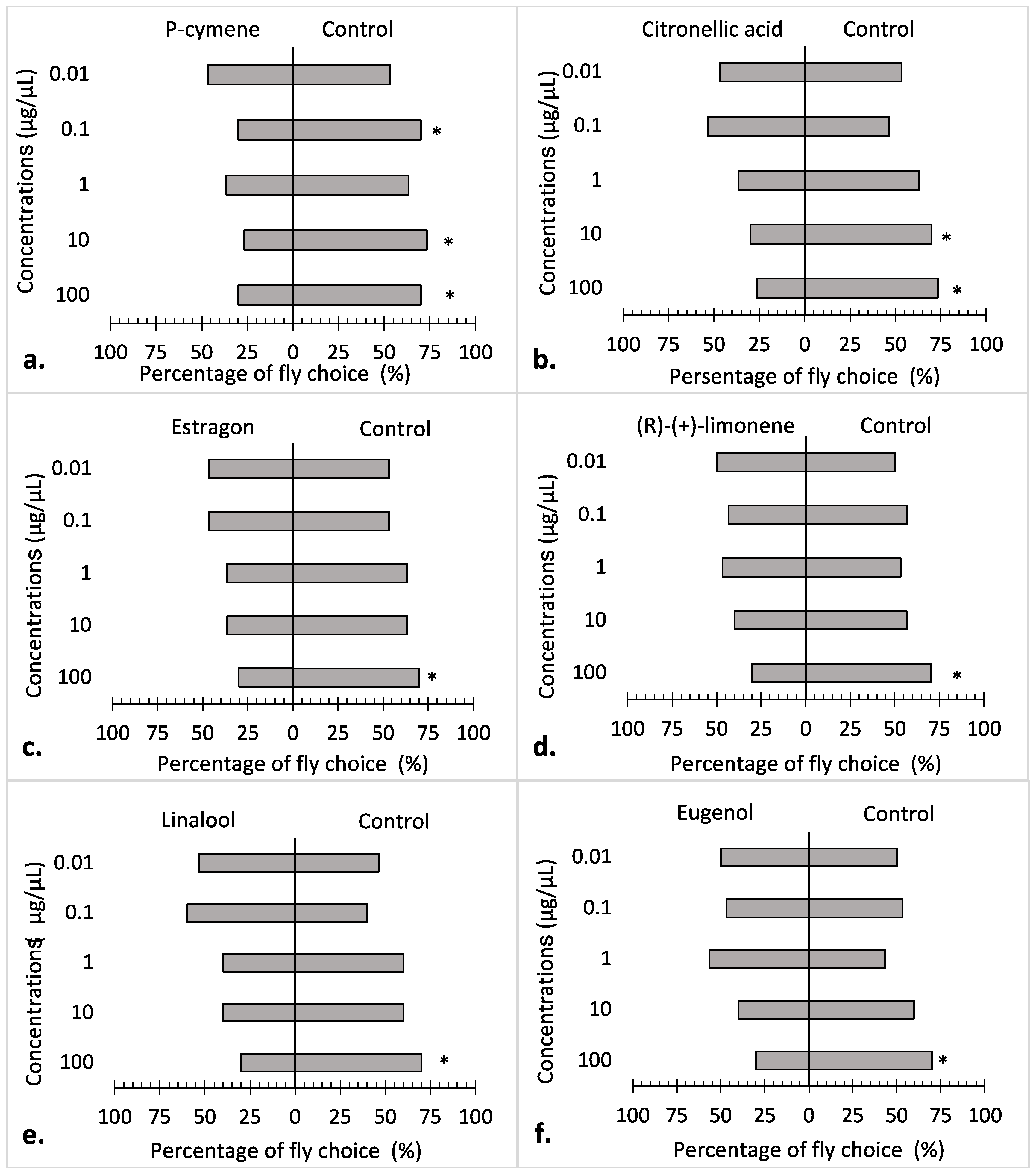
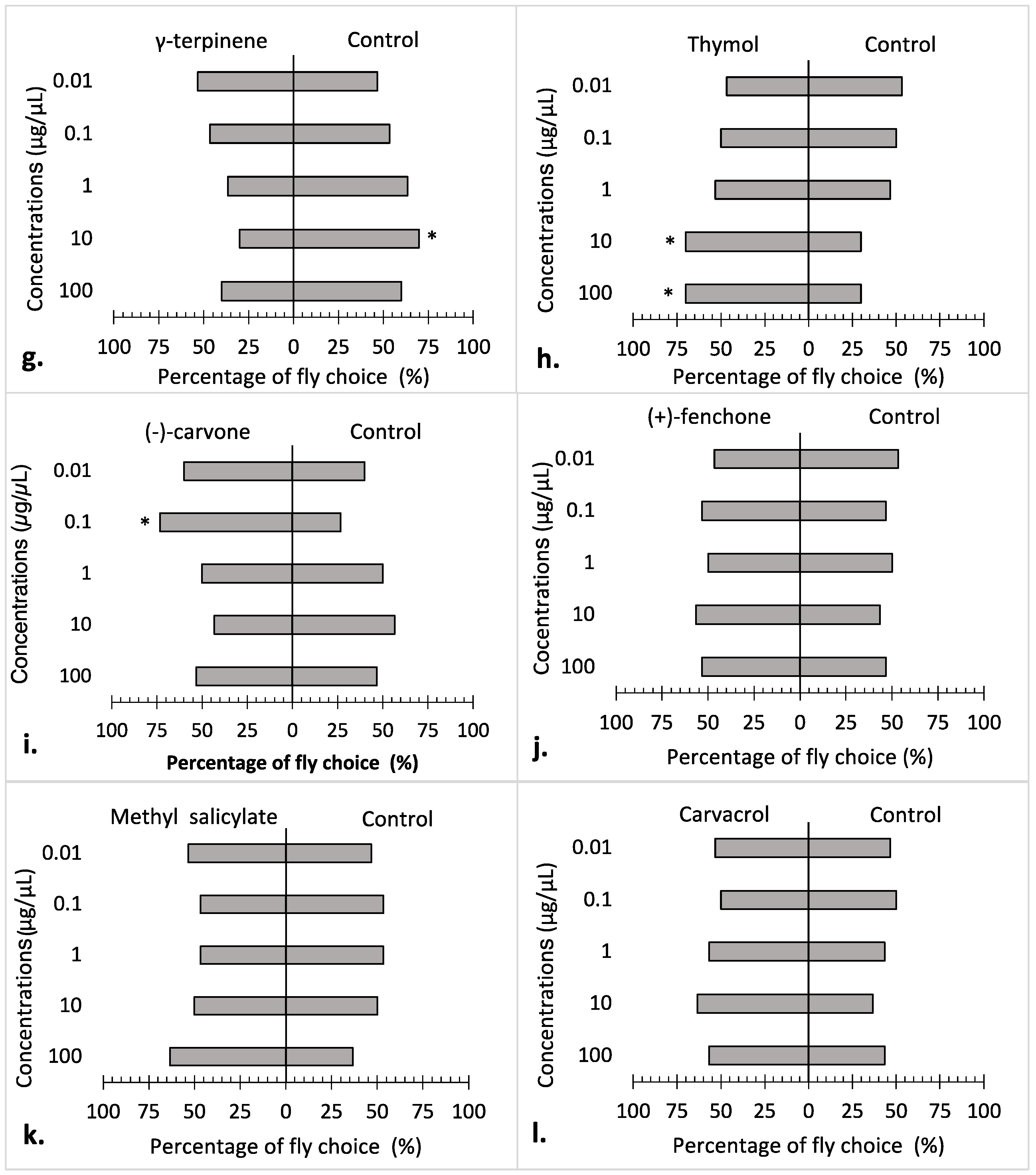
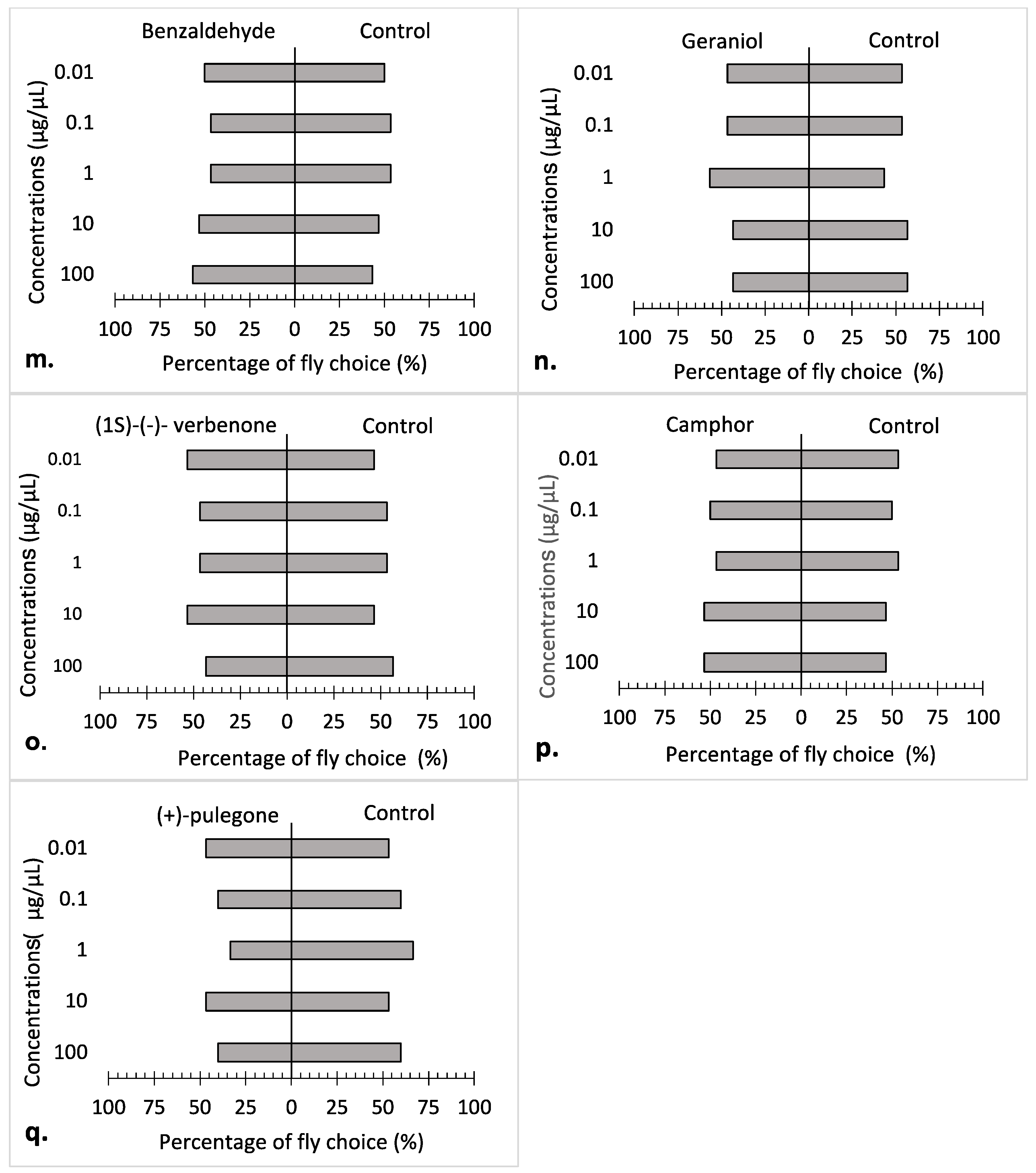
3.2. Repellency Bioassay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hogsette, J.; Farkas, R. Secretophagous and hematophagous higher Diptera. Contrib. A Man. Palearct. Diptera 2000, 1, 769–792. [Google Scholar]
- Crespo, D.C.; Lecuona, R.E.; Hogsette, J.A. Biological control: An important component in integrated management of Musca domestica (Diptera: Muscidae) in caged-layer poultry houses in Buenos Aires, Argentina. Biol. Control 1998, 13, 16–24. [Google Scholar] [CrossRef]
- Fotedar, R. Vector potential of houseflies (Musca domestica) in the transmission of Vibrio cholerae in India. Acta Trop. 2001, 78, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Efficacy of Mentha× piperita and Mentha citrata essential oils against housefly, Musca domestica L. Ind. Crops Prod. 2012, 39, 106–112. [Google Scholar] [CrossRef]
- Barin, A.; Arabkhazaeli, F.; Rahbari, S.; Madani, S.A. The housefly, Musca domestica, as a possible mechanical vector of Newcastle disease virus in the laboratory and field. Med. Vet. Entomol. 2010, 24, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Khamesipour, F.; Lankarani, K.B.; Honarvar, B.; Kwenti, T.E. A systematic review of human pathogens carried by the housefly (Musca domestica L.). BMC Public Health 2018, 18, 1049. [Google Scholar] [CrossRef] [PubMed]
- Merchant, M.E.; Flanders, R.V.; Williams, R.E. Seasonal abundance and parasitism of house fly (Diptera: Muscidae) pupae in enclosed, shallow-pit poultry houses in Indiana. Environ. Entomol. 1987, 16, 716–721. [Google Scholar] [CrossRef]
- Pugh, D.G.; Hu, X.P.; Blagburn, B. Habronemiasis: Biology, signs, and diagnosis, and treatment and prevention of the nematodes and vector flies. J. Equine Vet. Sci. 2014, 34, 241–248. [Google Scholar] [CrossRef]
- Premaalatha, B.; Chandrawathani, P.; Jamnah, O.; Erwanas, A.; Lily Rozita, M.; Ramlan, M. Intestinal cestode Choanotaenia infundibulum in chicken. Malays. J. Vet. Res. 2014, 5, 73–75. [Google Scholar]
- Oaya, C.; Malgwi, A.; Degri, M.; Samaila, A. Impact of synthetic pesticides utilization on humans and the environment: An overview. Agric. Sci. Technol. 2019, 4, 279–286. [Google Scholar] [CrossRef]
- Ali, S.; Ullah, M.I.; Sajjad, A.; Shakeel, Q.; Hussain, A. Environmental and health effects of pesticide residues. In Sustainable Agriculture Reviews 48: Pesticide Occurrence, Analysis and Remediation Vol. 2 Analysis; Springer: Berlin/Heidelberg, Germany, 2021; pp. 311–336. [Google Scholar]
- Kaufman, P.E.; Nunez, S.C.; Geden, C.J.; Scharf, M.E. Selection for resistance to imidacloprid in the house fly (Diptera: Muscidae). J. Econ. Entomol. 2010, 103, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 741. [Google Scholar] [CrossRef]
- Dos Santos, V.F.; Abeijon, L.M.; da Cruz Araújo, S.H.; Garcia, F.R.M.; de Oliveira, E.E. The Potential of plant-based biorational products for the Drosophila suzukii control: Current status, opportunities, and limitations. Neotrop. Entomol. 2023, 53, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Regnault-Roger, C. Recherche de nouveaux biopesticides d’origine végétale à caractère insecticide: Démarche méthodologique et application aux plantes aromatiques méditerranéennes. Biopestic. D’origine Végétale 2008, 25–49. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Singh, K.D.; Mobolade, A.J.; Bharali, R.; Sahoo, D.; Rajashekar, Y. Main plant volatiles as stored grain pest management approach: A review. J. Agric. Food Res. 2021, 4, 100127. [Google Scholar] [CrossRef]
- Suthisut, D.; Fields, P.G.; Chandrapatya, A. Contact toxicity, feeding reduction, and repellency of essential oils from three plants from the ginger family (Zingiberaceae) and their major components against Sitophilus zeamais and Tribolium castaneum. J. Econ. Entomol. 2011, 104, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Nerio, L.S.; Olivero-Verbel, J.; Stashenko, E. Repellent activity of essential oils: A review. Bioresour. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef]
- Isman, M.B. A renaissance for botanical insecticides? Pest Manag. Sci. 2015, 71, 1587–1590. [Google Scholar] [CrossRef]
- Xie, Y.; Jin, H.; Yang, X.; Gu, Q.; Zhang, D. Toxicity of the essential oil from Thymus serpyllum and thymol to larvae and pupae of the housefly Musca domestica L. (Diptera: Muscidae). Environ. Sci. Pollut. Res. 2020, 27, 35330–35340. [Google Scholar] [CrossRef]
- Yoon, J.; Tak, J.-H. Cuticular property affects the insecticidal synergy of major constituents in thyme oil against houseflies, Musca domestica. Sci. Rep. 2023, 13, 12654. [Google Scholar] [CrossRef] [PubMed]
- Soonwera, M.; Sinthusiri, J.; Passara, H.; Moungthipmalai, T.; Puwanard, C.; Sittichok, S.; Murata, K. Combinations of lemongrass and star anise essential oils and their main constituent: Synergistic housefly repellency and safety against non-target organisms. Insects 2024, 15, 210. [Google Scholar] [CrossRef] [PubMed]
- Soonwera, M.; Moungthipmalai, T.; Puwanard, C.; Sittichok, S.; Sinthusiri, J.; Passara, H. Adulticidal synergy of two plant essential oils and their major constituents against the housefly Musca domestica and bioassay on non-target species. Heliyon 2024, 10, e26910. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.R.M.; Ovruski, S.M.; Suárez, L.; Cancino, J.; Liburd, O.E. Biological control of tephritid fruit flies in the Americas and Hawaii: A review of the use of parasitoids and predators. Insects 2020, 11, 662. [Google Scholar] [CrossRef]
- Kaufman, P.E.; Nunez, S.C.; Mann, R.S.; Geden, C.J.; Scharf, M.E. Nicotinoid and pyrethroid insecticide resistance in houseflies (Diptera: Muscidae) collected from Florida dairies. Pest Manag. Sci. Former. Pestic. Sci. 2010, 66, 290–294. [Google Scholar] [CrossRef]
- Hogsette, J.A. New diets for production of house flies and stable flies (Diptera: Muscidae) in the laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef]
- Pavela, R. Insecticidal properties of several essential oils on the house fly (Musca domestica L.). Phytother. Res. 2008, 22, 274–278. [Google Scholar] [CrossRef]
- West, L.S. The Housefly: Its Natural History, Medical Importance, and Control; CABI: Wallingford, UK, 1951. [Google Scholar]
- Haselton, A.T.; Acevedo, A.; Kuruvilla, J.; Werner, E.; Kiernan, J.; Dhar, P. Repellency of α-pinene against the house fly, Musca domestica. Phytochemistry 2015, 117, 469–475. [Google Scholar] [CrossRef]
- Yang, L.; Hu, X.P.; Allan, S.A.; Alborn, H.T.; Bernier, U.R. Electrophysiological and behavioral responses of the kudzu bug, Megacopta cribraria (Hemiptera: Plataspidae), to volatile compounds from kudzu and soybean plants. J. Agric. Food Chem. 2019, 67, 4177–4183. [Google Scholar] [CrossRef]
- Norris, E.J.; Kline, J.; Bloomquist, J.R. Repellency and toxicity of vapor-active benzaldehydes against Aedes aegypti. ACS Infect. Dis. 2023, 10, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Rice, P.J.; Coats, J.R. Insecticidal properties of several monoterpenoids to the house fly (Diptera: Muscidae), red flour beetle (Coleoptera: Tenebrionidae), and southern corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1994, 87, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Tsao, R.; Peterson, C.; Coats, J.R. Insecticidal activity of monoterpenoids to western corn rootworm (Coleoptera: Chrysomelidae), twospotted spider mite (Acari: Tetranychidae), and house fly (Diptera: Muscidae). J. Econ. Entomol. 1997, 90, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.L.; Boonchu, N.; Sukontason, K.; Choochote, W. Effects of eucalyptol on house fly (Diptera: Muscidae) and blow fly (Diptera: Calliphoridae). Rev. Inst. Med. Trop. São Paulo 2004, 46, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Mee, K.C.; Sulaiman, S.; Othman, H. Efficacy of Piper aduncum extract against the adult housefly (Musca domestica). J. Trop. Med. Parasitol. 2009, 32, 52–57. [Google Scholar]
- Carrière, Y. Haplodiploidy, sex, and the evolution of pesticide resistance. J. Econ. Entomol. 2003, 96, 1626–1640. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Sivakumar, R.; Rajeswary, M.; Veerakumar, K. Mosquito larvicidal activity of thymol from essential oil of Coleus aromaticus Benth. against Culex tritaeniorhynchus, Aedes albopictus, and Anopheles subpictus (Diptera: Culicidae). Parasitol. Res. 2013, 112, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Geden, C.; Nayduch, D.; Scott, J.; Burgess, I.V.E.; Gerry, A.; Kaufman, P.; Thomson, J.; Pickens, V.; Machtinger, E. House fly (Diptera: Muscidae): Biology, pest status, current management prospects, and research needs. J. Integr. Pest. Manag. 2021, 12, 39. [Google Scholar] [CrossRef]
- Ahmadi, E.; Khajehali, J.; Jonckheere, W.; Van Leeuwen, T. Biochemical and insecticidal effects of plant essential oils on insecticide resistant and susceptible populations of Musca domestica L. point to a potential cross-resistance risk. Pestic. Biochem. Physiol. 2022, 184, 105115. [Google Scholar] [CrossRef]
- Gross, A.D.; Temeyer, K.B.; Day, T.A.; de León, A.A.P.; Kimber, M.J.; Coats, J.R. Interaction of plant essential oil terpenoids with the southern cattle tick tyramine receptor: A potential biopesticide target. Chem. Biol. Interact. 2017, 263, 1–6. [Google Scholar] [CrossRef]
- Norris, E.J.; Gross, A.D.; Kimber, M.J.; Bartholomay, L.; Coats, J. Plant terpenoids modulate α-adrenergic type 1 octopamine receptor (PaOA1) isolated from the American cockroach (Periplaneta americana). In Advances in the Biorational Control of Medical and Veterinary Pests; ACS Publications: Washington, DC, USA, 2018; pp. 219–235. [Google Scholar]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Quantitative structure–activity relationships of monoterpenoid binding activities to the housefly GABA receptor. Pest Manag. Sci. 2012, 68, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.-S.; Park, B.-S.; Ku, S.-K.; Lee, S.-E. Repellent activities of essential oils and monoterpenes against Culex pipiens pallens. J. Am. Mosq. Control. Assoc. 2002, 18, 348–351. [Google Scholar] [PubMed]
- Yoon, C.; Kang, S.-H.; Jang, S.-A.; Kim, Y.-J.; Kim, G.-H. Repellent efficacy of caraway and grapefruit oils for Sitophilus oryzae (Coleoptera: Curculionidae). J. Asia-Pac. Entomol. 2007, 10, 263–267. [Google Scholar] [CrossRef]
- Yoon, C.; Kang, S.-H.; Yang, J.-O.; Noh, D.-J.; Indiragandhi, P.; Kim, G.-H. Repellent activity of citrus oils against the cockroaches Blattella germanica, Periplaneta americana and P. fuliginosa. J. Pestic. Sci. 2009, 34, 77–88. [Google Scholar] [CrossRef]
- Gillij, Y.G.; Gleiser, R.M.; Zygadlo, J.A. Mosquito repellent activity of essential oils of aromatic plants growing in Argentina. Bioresour. Technol. 2008, 99, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.C.; Junnila, A.; Kravchenko, V.D.; Revay, E.E.; Butler, J.; Schlein, Y. Indoor protection against mosquito and sand fly bites: A comparison between citronella, linalool, and geraniol candles. J. Am. Mosq. Control. Assoc. 2008, 24, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Obeng-Ofori, D.; Reichmuth, C. Bioactivity of eugenol, a major component of essential oil of Ocimum suave (Wild.) against four species of stored-product Coleoptera. Int. J. Pest Manag. 1997, 43, 89–94. [Google Scholar] [CrossRef]
- Pandey, S.K.; Upadhyay, S.; Tripathi, A.K. Insecticidal and repellent activities of thymol from the essential oil of Trachyspermum ammi (Linn) Sprague seeds against Anopheles stephensi. Parasitol. Res. 2009, 105, 507–512. [Google Scholar] [CrossRef]
- Toloza, A.C.; Zygadlo, J.; Cueto, G.M.; Biurrun, F.; Zerba, E.; Picollo, M.I. Fumigant and repellent properties of essential oils and component compounds against permethrin-resistant Pediculus humanus capitis (Anoplura: Pediculidae) from Argentina. J. Med. Entomol. 2006, 43, 889–895. [Google Scholar] [CrossRef]
- Kim, S.-I.; Yoon, J.-S.; Jung, J.W.; Hong, K.-B.; Ahn, Y.-J.; Kwon, H.W. Toxicity and repellency of origanum essential oil and its components against Tribolium castaneum (Coleoptera: Tenebrionidae) adults. J. Asia-Pac. Entomol. 2010, 13, 369–373. [Google Scholar] [CrossRef]
- Vartak, P.H.; Sharma, R.N. Vapour toxicity & repellence of some essential oils & terpenoids to adults of Aedes aegypti (L) (Diptera: Culicidae). Indian J. Med. Res. 1993, 97, 122–127. [Google Scholar] [PubMed]
- Ibrahim, M.A.; Kainulainen, P.; Aflatuni, A.; Tiilikkala, K.; Holopainen, J.K. Insecticidal, repellent, antimicrobial activity and phytotoxicity of essential oils: With special reference to limonene and its suitability for control of insect pests. Agric. Food Sci. 2001, 10, 243–259. [Google Scholar] [CrossRef]
- Hao, H.; Sun, J.; Dai, J. Dose-dependent behavioral response of the mosquito Aedes albopictus to floral odorous compounds. J. Insect Sci. 2013, 13, 127. [Google Scholar] [CrossRef] [PubMed]
- Naik, D.G.; Vaidya-Kannur, H.; Deshpande, P.V.; Dandge, C.N.; Reddy, G.V.P. Potential use of an essential oil from the flower of Swertia densifolia as a repellent for Apis florea (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 2015, 108, 18–25. [Google Scholar] [CrossRef]
- Fang, R.; Jiang, C.H.; Wang, X.Y.; Zhang, H.M.; Liu, Z.L.; Zhou, L.; Du, S.S.; Deng, Z.W. Insecticidal activity of essential oil of Carum carvi fruits from China and its main components against two grain storage insects. Molecules 2010, 15, 9391–9402. [Google Scholar] [CrossRef] [PubMed]
- Abdelgaleil, S.A.; Mohamed, M.I.; Badawy, M.E.; El-arami, S.A. Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (Herbst) and their inhibitory effects on acetylcholinesterase activity. J. Chem. Ecol. 2009, 35, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Hummelbrunner, L.A.; Isman, M.B. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.K.; Appel, A.G.; Sims, S.R. Topical toxicity of essential oils to the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 2010, 103, 448–459. [Google Scholar] [CrossRef]
- Gallardo, A.; Picollo, M.I.; Mougabure-Cueto, G. Lethal activity of individual and mixed monoterpenoids of geranium essential oil on Musca domestica. Parasitol. Res. 2015, 114, 1229–1232. [Google Scholar] [CrossRef]
- Wang, C.F.; Yang, K.; Zhang, H.M.; Cao, J.; Fang, R.; Liu, Z.L.; Du, S.S.; Wang, Y.Y.; Deng, Z.W.; Zhou, L. Components and insecticidal activity against the maize weevils of Zanthoxylum schinifolium fruits and leaves. Molecules 2011, 16, 3077–3088. [Google Scholar] [CrossRef]
- Kim, S.-I.; Lee, D.-W. Toxicity of basil and orange essential oils and their components against two coleopteran stored products insect pests. J. Asia-Pac. Entomol. 2014, 17, 13–17. [Google Scholar] [CrossRef]
- Waliwitiya, R.; Isman, M.B.; Vernon, R.S.; Riseman, A. Insecticidal activity of selected monoterpenoids and rosemary oil to Agriotes obscurus (Coleoptera: Elateridae). J. Ecol. Entomol. 2005, 98, 1560–1565. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Yang, K.; You, C.-X.; Wang, Y.; Wang, C.-F.; Wu, Y.; Geng, Z.-F.; Su, Y.; Du, S.-S.; Deng, Z.-W. Bioactivity of essential oil from Artemisia stolonifera (Maxim.) Komar. and its main compounds against two stored-product insects. J. Oleo Sci. 2015, 64, 299–307. [Google Scholar] [CrossRef]
- Norris, E.J.; Gross, A.D.; Dunphy, B.M.; Bessette, S.; Bartholomay, L.; Coats, J.R. Comparison of the insecticidal characteristics of commercially available plant essential oils against Aedes aegypti and Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 993–1002. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Liu, J.-Y.; Huang, C.-G.; Hsui, Y.-R.; Chen, W.-J.; Chang, S.-T. Insecticidal activities of leaf essential oils from Cinnamomum osmophloeum against three mosquito species. Bioresour. Technol. 2009, 100, 457–464. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | n | Slope ± SE | LD50 (95% CI) (µg/fly) | LD95 (95% CI) (µg/fly) | χ2 |
|---|---|---|---|---|---|
| Thymol | 500 | 1.8 ± 0.2 | 43.8 (34.1–55.6) | 360.4 (221.0–823.7) | 36.2 |
| (+)-Pulegone | 500 | 5.0 ± 0.6 | 73.0 (62.1–82.7) | 155.6 (125.9–241.8) | 46.7 |
| Eugenol | 500 | 5.3 ± 0.4 | 89.5 (71.5–108.1) | 182.9 (144.7–285.5) | 110.6 |
| Carvacrol | 500 | 3.4 ± 0.2 | 90.8 (77.2–106.5) | 275.7 (216.6–390.8) | 40.0 |
| Citronellic acid | 500 | 2.5 ± 0.2 | 93.4 (75.5–115.7) | 430.1 (300.3–754.9) | 44.6 |
| Benzaldehyde | 500 | 2.6 ± 0.2 | 94.7 (78.7–112.5) | 392.4 (294.2–602.4) | 29.4 |
| Geraniol | 500 | 2.7 ± 0.2 | 99.7 (85.7–116.4) | 404.7 (311.0–582.9) | 26.6 |
| p-Cymene | 600 | 3.1 ± 0.3 | 119.8 (106.1–133.4) | 404.5 (342.7–502.7) | 21.7 |
| Estragole | 500 | 3.3 ± 0.3 | 189.5 (161.4–222.5) | 603.3 (460.4–922.1) | 43.8 |
| (-)-Carvone | 500 | 4.8 ± 0.4 | 213.7 (184.6–242.0) | 469.8 (396.8–605.8) | 45.0 |
| (R)-(+)-Limonene | 500 | 2.1 ± 0.2 | 226.6 (185.1–286.6) | 1208.5 (772.2–2751.4) | 31.4 |
| γ-Terpinene | 500 | 4.4 ± 0.3 | 236.5 (216.2–256.8) | 556.9 (486.9–667.8) | 21.8 |
| Linalool | 600 | 2.2 ± 0.2 | 238.1 (201.7–282.2) | 1322.1 (943.4–2169.8) | 37.6 |
| Methyl salicylate | 500 | 5.0 ± 0.4 | 260.7 (237.4–285.5) | 557.9 (479.6–694.1) | 27.9 |
| (+)-Fenchone | 700 | 3.8 ± 0.3 | 405.1 (359.3–457.7) | 1094.9 (869.0–1590.4) | 75.5 |
| (1S)-(-)-Verbenone | 600 | 5.0 ± 0.4 | 426.7 (399.9–454.8) | 909.3 (802.0–1081.1) | 28.1 |
| Camphor | 500 | 9.2 ± 0. 7 | 512.1 (486.6–539.3) | 774.9 (709.4–883.0) | 33.6 |
| Chemical | n | Slope ± SE | LD50 (95% CI) (µg/fly) | LD95 (95% CI) (µg/fly) | χ2 |
|---|---|---|---|---|---|
| Thymol | 500 | 1.9 ± 0.2 | 41.1 (32.1–51.8) | 317.2 (199.8–686.4) | 36.5 |
| (+)-Pulegone | 500 | 8.8 ± 0.9 | 68.2 (63.0–73.0) | 104.8 (94.7–123.4) | 69.0 |
| Eugenol | 500 | 5.7 ± 0.4 | 78.5 (64.3–93.5) | 153.1 (123.6–222.6) | 93.7 |
| Carvacrol | 500 | 3.5 ± 0.2 | 80.6 (68.3–95. 0) | 237.7 (186.1–340.2) | 42.7 |
| Geraniol | 500 | 2.6 ± 0.2 | 85.6 (71.2–102.7) | 373.0 (274.2–586.6) | 34.6 |
| Citronellic acid | 500 | 2.4 ± 0.2 | 85.8 (69.9–105.0) | 430.3 (302.6–736.6) | 37.9 |
| Benzaldehyde | 500 | 2.6 ± 0.2 | 86.8 (71.8–103.7) | 381.2 (283.1–594.2) | 29.9 |
| p-Cymene | 600 | 3.1 ± 0.3 | 111.5 (98.5–124.4) | 375.9 (319.0–465.9) | 21.5 |
| Estragole | 500 | 2.9 ± 0.2 | 164.8 (135.6–199.1) | 614.8 (445.6–1047.3) | 51.1 |
| (-)-Carvone | 500 | 4.8 ± 0.4 | 190.4 (166.4–213.9) | 422.0 (360.9–527.6) | 35.2 |
| Linalool | 600 | 2.4 ± 0.2 | 209.7 (179.7–244.5) | 1042.4 (778.1–1580.2) | 35.3 |
| (R)-(+)-Limonene | 500 | 1.9 ± 0.2 | 213.4 (162.0–275.2) | 1356.9 (819.0–3558.8) | 32.4 |
| γ-Terpinene | 500 | 4.4 ± 0.3 | 221.6 (200.7–242.) | 526.5 (457.1–628.6) | 24.8 |
| Methyl salicylate | 500 | 4.5 ± 0.4 | 238.7 (221.4–256.5) | 549.5 (485.4–646.0) | 17.0 |
| (+)-Fenchone | 700 | 3.5 ± 0.3 | 385.3 (341.3–434.9) | 1138.6 (898.4–1653.3) | 66.6 |
| (1S)-(-)-Verbenone | 600 | 5.2 ± 0.4 | 410.0 (383.3–437.3) | 853.9 (756.2–1009.7) | 30.5 |
| Camphor | 500 | 8.8 ± 0.7 | 478.0 (454.3–502.3) | 734.7 (675.8–828.0) | 30.7 |
| Chemical | n | Slope ± SE | LD50 (95% CI) (µg/fly) | LD95 (95% CI) (µg/fly) | χ2 |
|---|---|---|---|---|---|
| Thymol | 150 | 2.0 ± 0.3 | 35.2 (19.4–59.7) | 227.5 (110.6–1659.9) | 5.7 |
| (+)-Pulegone | 125 | 2.5 ± 0.5 | 56.5 (43.3–75.3) | 262.9 (156.0–912.9) | 2.7 |
| Eugenol | 125 | 2.5 ± 0.5 | 60.9 (45.7–79.9) | 276.2 (174.9–703.2) | 2.5 |
| Carvacrol | 150 | 2.1 ± 0.4 | 77.4 (46.8–122.4) | 463.0 (232.4–3641.1) | 4.5 |
| Citronellic acid | 150 | 2.2 ± 0.4 | 92.0 (45.8–165.4) | 530.1 (249.8–7648.1) | 6.3 |
| Geraniol | 150 | 2.4 ± 0.5 | 93.1 (70.5–121.9) | 463.8 (284.2–1312.2) | 2.9 |
| Chemical | n | Slope ± SE | LD50 (95% CI) (µg/fly) | LD95 (95% CI) (µg/fly) | χ2 |
|---|---|---|---|---|---|
| Thymol | 150 | 1.8 ± 0.3 | 29.2 (13.8–53.2) | 244.1 (106.0–3264.8) | 6.2 |
| (+)-Pulegone | 125 | 2.3 ± 0.4 | 38.8 (28.4–50.5) | 207.2 (127.9–572.3) | 1.6 |
| Eugenol | 125 | 2.3 ± 0.4 | 45.1 (30.5–63.3) | 232.4 (120.4–1922.3) | 3.1 |
| Carvacrol | 150 | 2.0 ± 0.4 | 65.7 (48.1–87.2) | 430.0 (258.5–1132.0) | 2.7 |
| Citronellic acid | 150 | 2.4 ± 0.3 | 75.5 (55.3–100.7) | 327.1 (194.1–4273.7) | 3.7 |
| Geraniol | 150 | 2.6 ± 0.5 | 75.7 (48.5–107.6) | 479.4 (292.7–1175.5) | 6.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Hogsette, J.A.; Norris, E.J.; Hu, X.P. Topical Toxicity and Repellency Profiles of 17 Essential Oil Components against Insecticide-Resistant and Susceptible Strains of Adult Musca domestica (Diptera: Muscidae). Insects 2024, 15, 384. https://doi.org/10.3390/insects15060384
Tian Y, Hogsette JA, Norris EJ, Hu XP. Topical Toxicity and Repellency Profiles of 17 Essential Oil Components against Insecticide-Resistant and Susceptible Strains of Adult Musca domestica (Diptera: Muscidae). Insects. 2024; 15(6):384. https://doi.org/10.3390/insects15060384
Chicago/Turabian StyleTian, Yuexun, Jerome A. Hogsette, Edmund J. Norris, and Xing Ping Hu. 2024. "Topical Toxicity and Repellency Profiles of 17 Essential Oil Components against Insecticide-Resistant and Susceptible Strains of Adult Musca domestica (Diptera: Muscidae)" Insects 15, no. 6: 384. https://doi.org/10.3390/insects15060384
APA StyleTian, Y., Hogsette, J. A., Norris, E. J., & Hu, X. P. (2024). Topical Toxicity and Repellency Profiles of 17 Essential Oil Components against Insecticide-Resistant and Susceptible Strains of Adult Musca domestica (Diptera: Muscidae). Insects, 15(6), 384. https://doi.org/10.3390/insects15060384