

A New Genus of Praeaulacidae (Hymenoptera: Evanioidea) from Mid-Cretaceous Kachin Amber: Insights into a Putative New Praeaulacinae Subclade

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
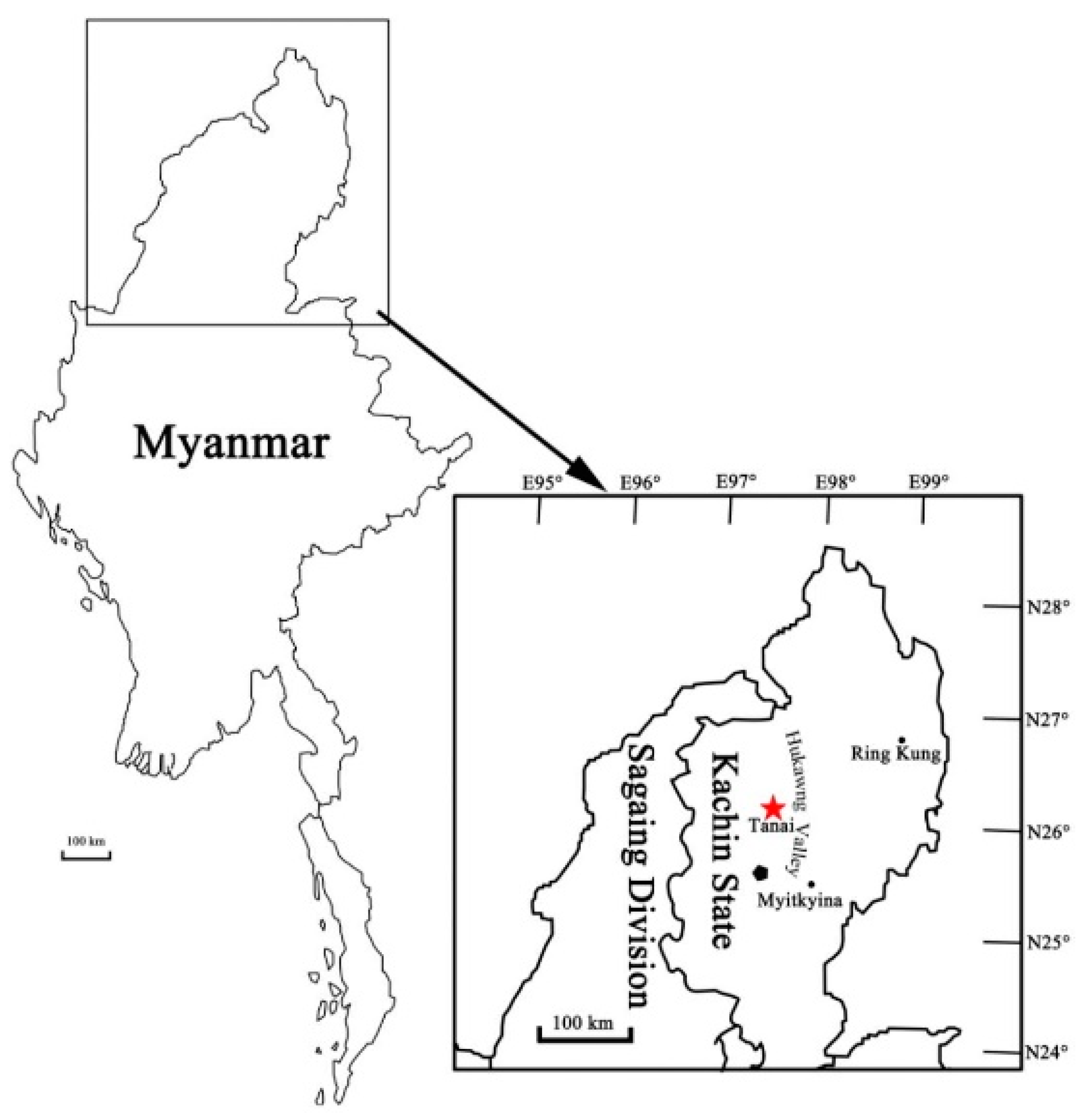
2. Materials and Methods
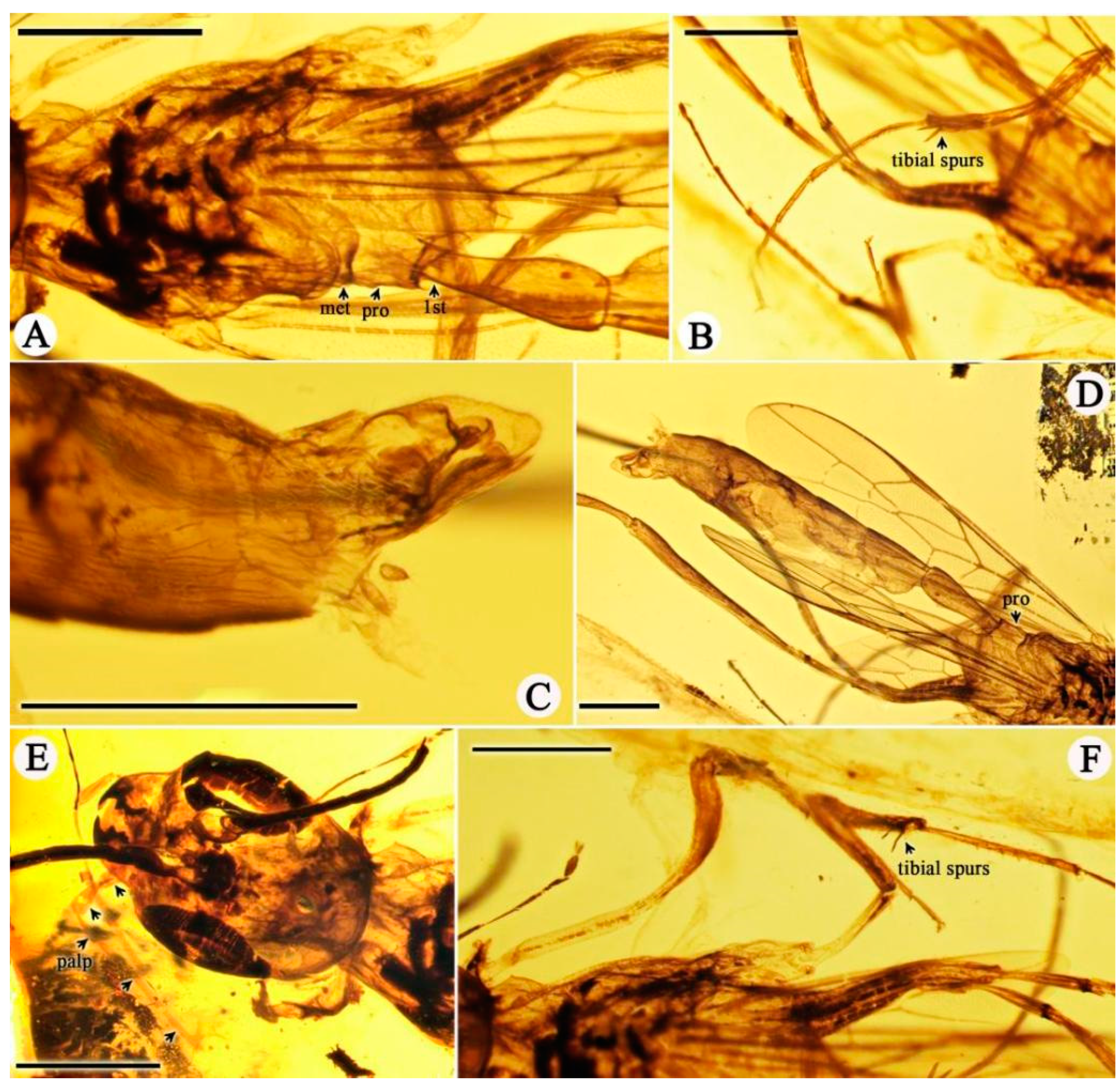
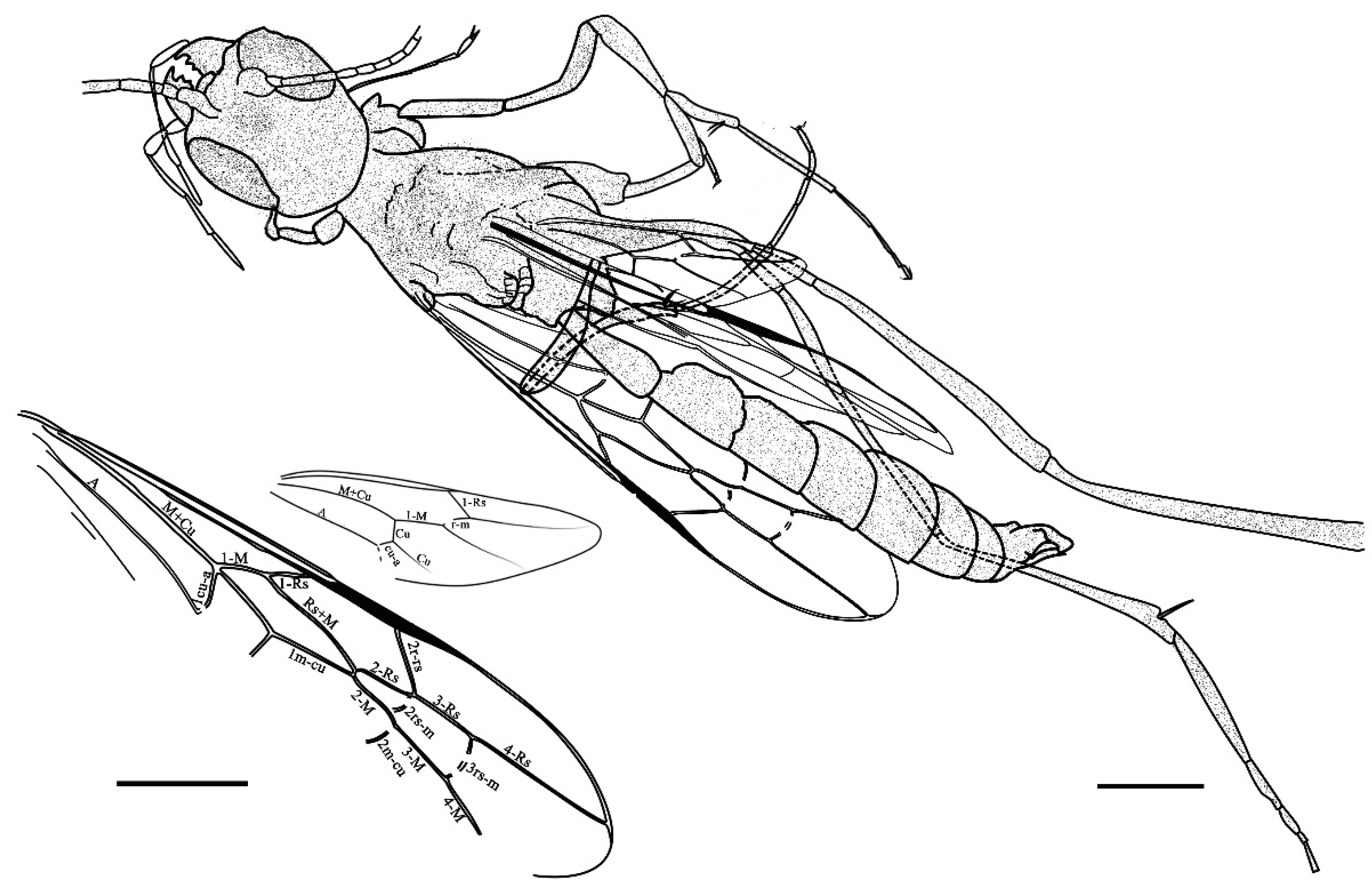
3. Results
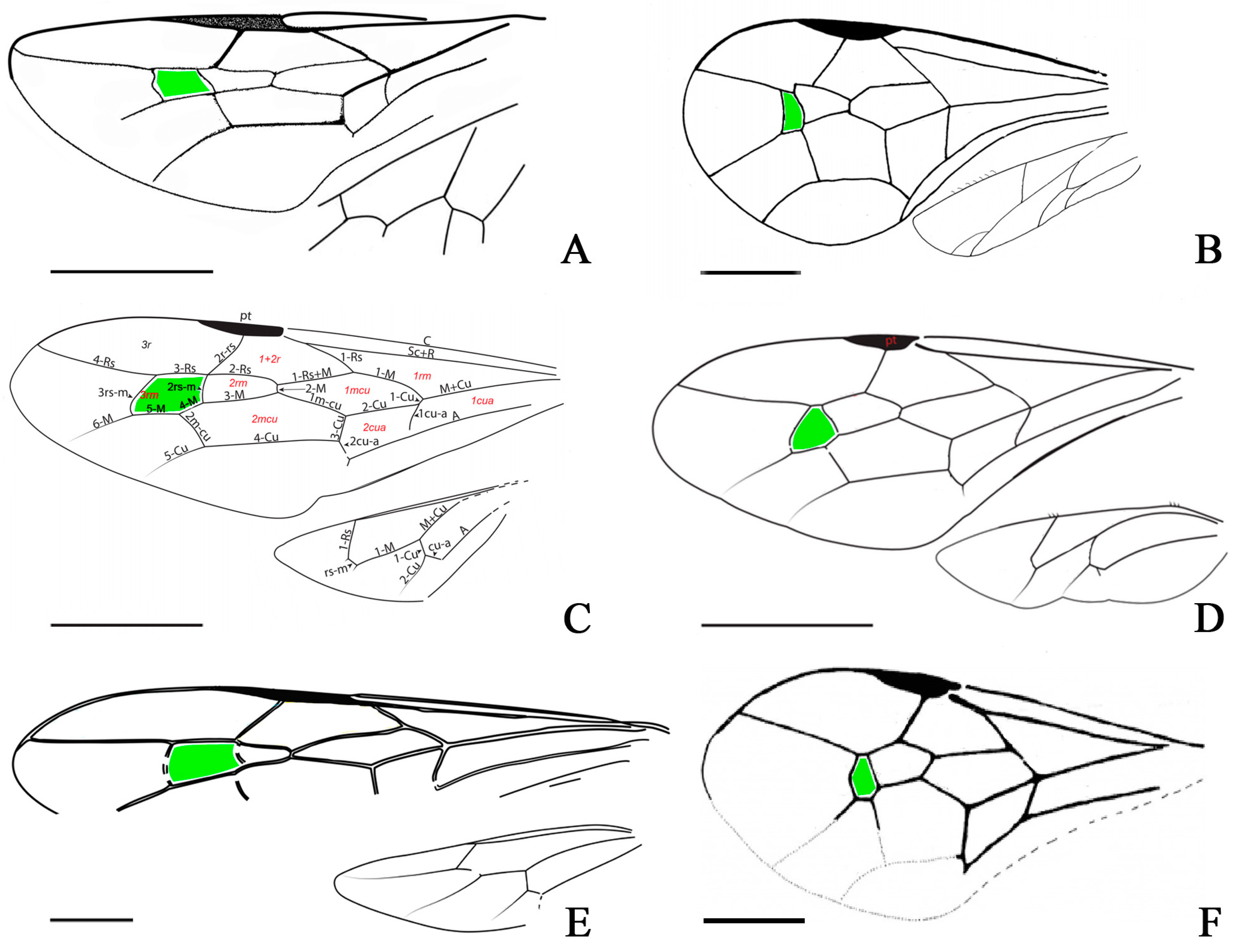
4. Discussion
- Comparisons with other Praeaulacidae from the Kachin amber biota
- A hidden phylogenetic signal in the fore wing venation of Praeaulacidae?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.C.; Rasnitsyn, A.P. Nevaniinae subfam. n., a new fossil taxon (Insecta: Hymenoptera: Evanioidea: Praeaulacidae) from the Middle Jurassic of Daohugou in Inner Mongolia, China. Insect Syst. Evol. 2007, 38, 149–166. [Google Scholar] [CrossRef]
- Li, L.; Rasnitsyn, A.P.; Shih, C.; Labandeira, C.C.; Buffington, M.; Li, D.; Ren, D. Phylogeny of Evanioidea (Hymenoptera, Apocrita), with descriptions of new Mesozoic species from China and Myanmar. Syst. Entomol. 2018, 43, 810–842. [Google Scholar] [CrossRef]
- Jouault, C.; Marechal, A.; Condamine, F.L.; Wang, B.; Nel, A.; Legendre, F.; Perrichot, V. Including fossils in phylogeny: A glimpse into the evolution of the superfamily Evanioidea (Hymenoptera: Apocrita) under tip-dating and the fossilized birth-death process. Zool. J. Linn. Soc. 2022, 194, 1396–1423. [Google Scholar] [CrossRef]
- Jouault, C.; Nel, A. New evanioid wasps (Hymenoptera: Evaniidae; Praeaulacidae) from mid-Cretaceous Kachin amber. Ann. Paléontol. 2024, 110, 102662. [Google Scholar]
- Li, L.; Rasnitsyn, A.P.; Shih, C.; Ren, D. A new genus and species of Praeaulacidae (Hymenoptera: Evanioidea) from Upper Cretaceous Myanmar amber. Cretac. Res. 2015, 55, 19–24. [Google Scholar] [CrossRef]
- Jouault, C.; Nel, A.; Perrichot, V. New evanioid wasps (Hymenoptera: Praeaulacidae, Aulacidae) from Cenomanian Burmese amber. Cretac. Res. 2020, 110, 104407. [Google Scholar] [CrossRef]
- Li, L.; Shih, C.; Yang, J.; Ren, D. A new praeaulacid genus with a new species in Praeaulacidae (Hymenoptera: Evanioidea) from mid-Cretaceous Myanmar amber. Cretac. Res. 2023, 146, 105483. [Google Scholar] [CrossRef]
- Jouault, C.; Rosse-Guillevic, S. A new genus of praeaulacid wasp (Hymenoptera: Evanioidea) from the mid-Cretaceous Kachin amber (Myanmar). Ann. Paléontol. 2023, 109, 102599. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P. Hymenopterans, Vespida, in Pozdne-Mezozoyskie Nasekomye Vostochnogo Zabaykal’ya. Tr. Paleontol. Instituta 1990, 239, 177–205. [Google Scholar]
- Rasnitsyn, A.P. Praeaulacidae (Hymenoptera) from the Upper Jurassic of Karatau. Paleontol. Zh. 1972, 1, 72–87. (In Russian) [Google Scholar]
- Rasnitsyn, A.P. Hymenoptera Apocrita of Mesozoic. Tr. Paleontol. Instituta 1975, 147, 1–134. (In Russian) [Google Scholar]
- Zhang, H.C.; Rasnitsyn, A.P. Middle Jurassic Praeaulacidae (Insecta: Hymenoptera: Evanioidea) of Inner Mongolia and Kazakhstan. J. Syst. Palaeontol. 2008, 6, 463–487. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P.; Zhang, H.C. Early evolution of Apocrita (Insect, Hymenoptera) as indicated by new findings in the Middle Jurassic of Daohugou, Northeast China. Acta Geol. Sin. 2010, 84, 834–873. [Google Scholar] [CrossRef]
- Shi, G.H.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.C.; Lei, W.Y.; Li, Q.L.; Li, X.H. Age constraint on Kachin amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Yu, T.T.; Thomson, U.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.Y.; Zhang, H.C.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, R.; Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Engel, M.S.; Sheng, X.; Shih, C.; Ren, D. Early evolution of wing scales prior to the rise of moths and butterflies. Curr. Biol. 2022, 32, 3808–3814.e2. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Bardin, J.; Wipfler, B.; Demers-Potvin, A.; Bai, M.; Tong, Y.; Chen, G.N.; Chen, H.; Zhao, Z.; Ren, D.; et al. A winged relative of ice-crawlers in amber bridges the cryptic extant Xenonomia and a rich fossil record. Insect Sci. 2024. [CrossRef]
- Lai, D.; Chen, P.; Li, S.; Xiang, X.; Ou, H.; Kang, N.; Yang, J.; Pang, H.; Shih, C.; Labandeira, C.C.; et al. The associated evolution of raptorial foreleg and mantispid diversification during 200 million years. Natl. Sci. Rev. 2023, 10, nwad278. [Google Scholar] [CrossRef]
- Zhang, Y.; Rasnitsyn, A.P.; Zhang, W.; Song, F.; Shih, C.; Ren, D.; Wang, Y.; Li, H.; Gao, T. Stem chewing lice on Cretaceous feathers preserved in amber. Curr. Biol. 2024, 34, 916–922.e1. [Google Scholar] [CrossRef]
- Wang, Z.; Vilhelmsen, L.; Rasnitsyn, A.P.; Viertler, A.; Shih, C.K.; Wen, S.S.; Yang, H.R.; Wu, Q.; Zhang, Y.J.; Ren, D.; et al. Specialized ovipositor sensilla of Cretaceous wasp (Insecta: Hymenoptera) possibly reveal a unique way of host detection. Cladistics 2024. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Zhang, W.; Fang, H.; Wang, J.; Shih, C.; Ren, D. Geometric morphometric analyses of Micropterigidae lineages (Lepidoptera)with two new species from mid-Cretaceous amber of northern Myanmar. Cretac. Res. 2024, 160, 105897. [Google Scholar] [CrossRef]
- Ma, Y.-N.; DU, S.-L.; Ren, D.; Yao, Y.-Z. A new genus and species of Nabinae (Heteroptera: Cimicomorpha: Nabidae) from mid-Cretaceous Burmese amber. Palaeoentomology 2024, 7, 306–313. [Google Scholar] [CrossRef]
- Engel, M.S. Two ensign wasps in Cretaceous amber from New Jersey and Myanmar (Hymenoptera: Evaniidae). Pol. J. Entomol. 2006, 75, 443–454. [Google Scholar]
- Engel, M.S. New evanioid wasps from the Cenomanian of Myanmar (Hymenoptera: Othbiodellithidae, Aulacidae), with s summary of family-group names among Evanioidea. Am. Mus. Novit. 2017, 3871, 1–28. [Google Scholar] [CrossRef]
- Engel, M.S.; Huang, D.; Alqarni, A.S.; Cai, C. A remarkable evanioid wasp in mid-Cretaceous amber from northern Myanmar (Hymenoptera: Evanioidea). Cretac. Res. 2016, 60, 121–127. [Google Scholar] [CrossRef]
- Poinar, G., Jr. A new genus and species of wasp (Hymenoptera: Evanioidea: Praeaulacidae) associated with an angiosperm flower in Burmese amber. Palaeoentomology 2019, 2, 474–481. [Google Scholar] [CrossRef]
- Shih, P.J.; Li, L.; Li, D.; Ren, D. Application of geometric morphometric analyses to confirm three new wasps of Evaniidae (Hymenoptera: Evanioidea) from mid-Cretaceous Myanmar amber. Cretac. Res. 2019, 109, 104249. [Google Scholar] [CrossRef]
- Goulet, H.; Huber, J.T. Hymenoptera of the World: An Identification Guide to Families; Agriculture Canada Publication: Ottawa, ON, Canada, 1993; p. 680. [Google Scholar]
- Li, L.; Rasnitsyn, A.P.; Shih, C.; Ren, D. Anomopterellidae Restored, with Two New Genera and Its Phylogeny in Evanioidea (Hymenoptera). PLoS ONE 2013, 8, e82587. [Google Scholar] [CrossRef]
- Rasnitsyn, A.P.; Martínez-Delclòs, X. Wasps (Insecta: Vespida = Hymenoptera) from the Early Cretaceous of Spain. Acta Geol. Hisp. 2000, 35, 65–95. [Google Scholar]
- Li, L.-F.; Shih, C.-K.; Ren, D. Two new species of Nevania (Hymenoptera: Evanioidea: Praeaulacidae: Nevaniinae) from the Middle Jurassic of China. Alcheringa Australas. J. Palaeontol. 2014, 39, 140–147. [Google Scholar] [CrossRef]
- Zhang, H.C.; Zhang, J.F. Discovery of Praeaulacidae (Insecta Hymenoptera) in China and its significance. Acta Micro-Palaeontol. Sin. 2000, 17, 416–421. [Google Scholar]
- Basibuyuk, H.H.; Rasnitsyn, A.P.; Fitton, M.G.; Quicke, D.L.J. An archaic new genus of Evaniidae (Insecta: Hymenoptera) and implications for the biology of ancestral evanioids. Bull. Nat. Hist. Mus. Lond. (Geol.) 2000, 56, 54–56. [Google Scholar]
- Rasnitsyn, A.P. The origin and evolution of Lower Hymenoptera. Trans. Paleontol. Inst. 1969, 123, 1–196. (In Russian) [Google Scholar]
- Rasnitsyn, A.P. Origin and evolution of Hymenoptera. Trans. Paleontol. Inst. 1980, 174, 1–192. (In Russian) [Google Scholar]
- Jouault, C.; Ngô-Muller, V.; Pouillon, J.-M.; Nel, A. New Burmese amber fossils clarify the evolution of bethylid wasps (Hymenoptera: Chrysidoidea). Zool. J. Linn. Soc. 2021, 191, 1044–1058. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Jouault, C.; Li, L.; Shih, C.; Ren, D. A New Genus of Praeaulacidae (Hymenoptera: Evanioidea) from Mid-Cretaceous Kachin Amber: Insights into a Putative New Praeaulacinae Subclade. Insects 2024, 15, 351. https://doi.org/10.3390/insects15050351
Yang J, Jouault C, Li L, Shih C, Ren D. A New Genus of Praeaulacidae (Hymenoptera: Evanioidea) from Mid-Cretaceous Kachin Amber: Insights into a Putative New Praeaulacinae Subclade. Insects. 2024; 15(5):351. https://doi.org/10.3390/insects15050351
Chicago/Turabian StyleYang, Jingtao, Corentin Jouault, Longfeng Li, Chungkun Shih, and Dong Ren. 2024. "A New Genus of Praeaulacidae (Hymenoptera: Evanioidea) from Mid-Cretaceous Kachin Amber: Insights into a Putative New Praeaulacinae Subclade" Insects 15, no. 5: 351. https://doi.org/10.3390/insects15050351
APA StyleYang, J., Jouault, C., Li, L., Shih, C., & Ren, D. (2024). A New Genus of Praeaulacidae (Hymenoptera: Evanioidea) from Mid-Cretaceous Kachin Amber: Insights into a Putative New Praeaulacinae Subclade. Insects, 15(5), 351. https://doi.org/10.3390/insects15050351