
Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae)

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Sample Collection under Various Biotic Factors
2.3. Sample Collection under Various Abiotic Conditions
2.4. RNA Extraction and cDNA Synthesis
2.5. Candidate Reference Gene Selection and Primer Design
2.6. qRT-PCR Analysis
2.7. Statistical Analysis
2.8. Validation of Reference Genes
3. Results
3.1. Primer Specificity and Efficiency
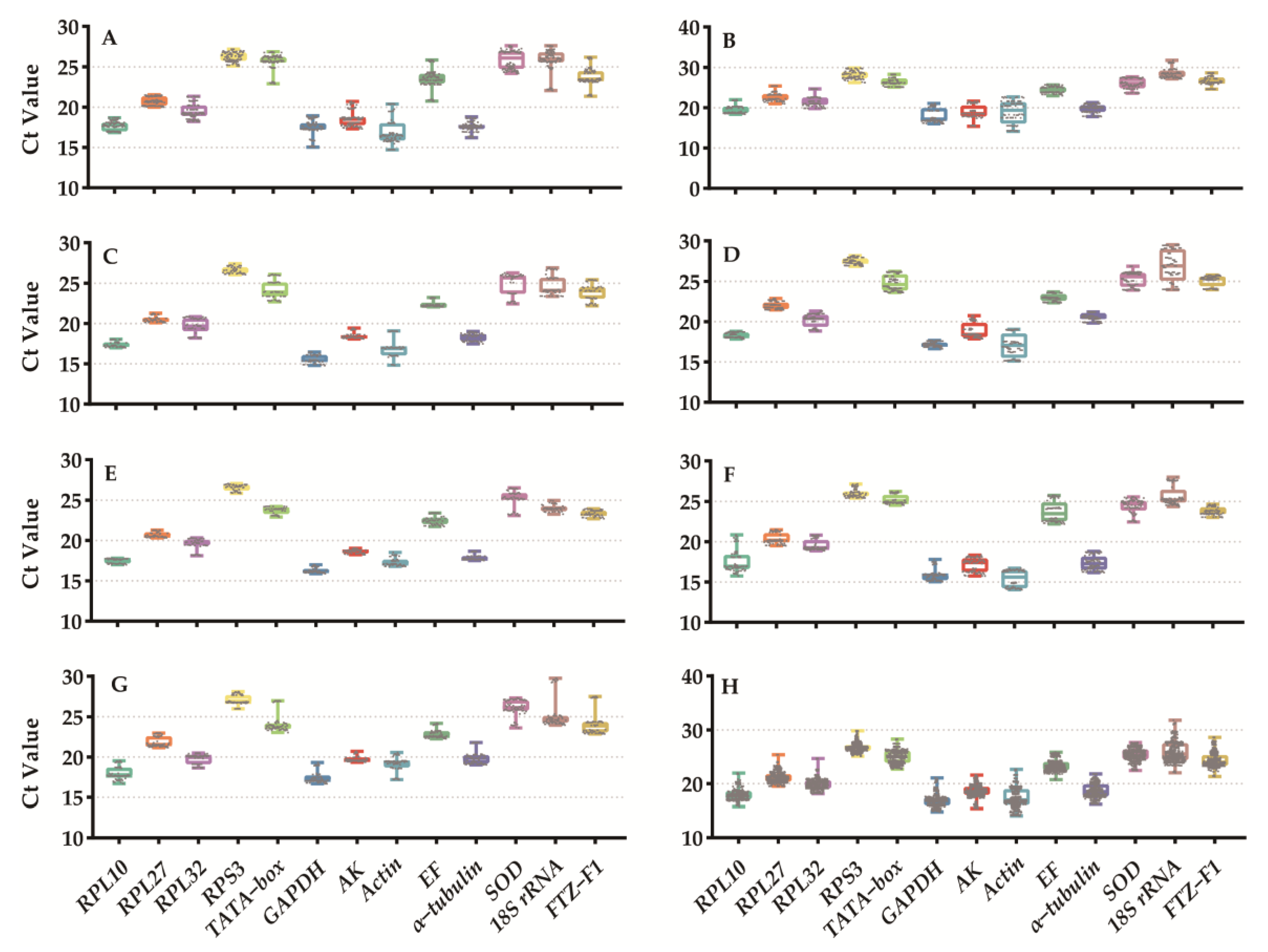
3.2. Expression Profiles of Candidate Reference Genes
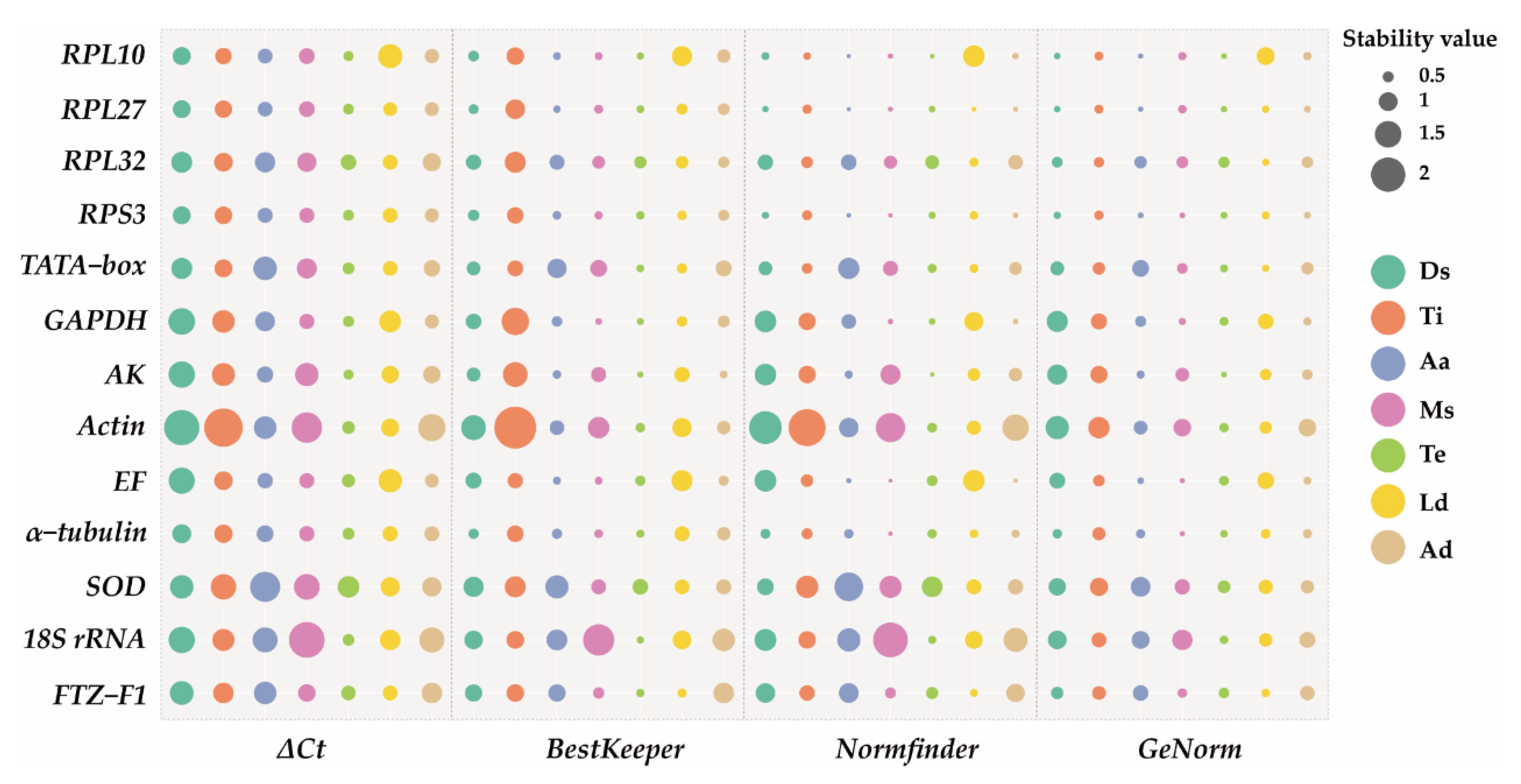
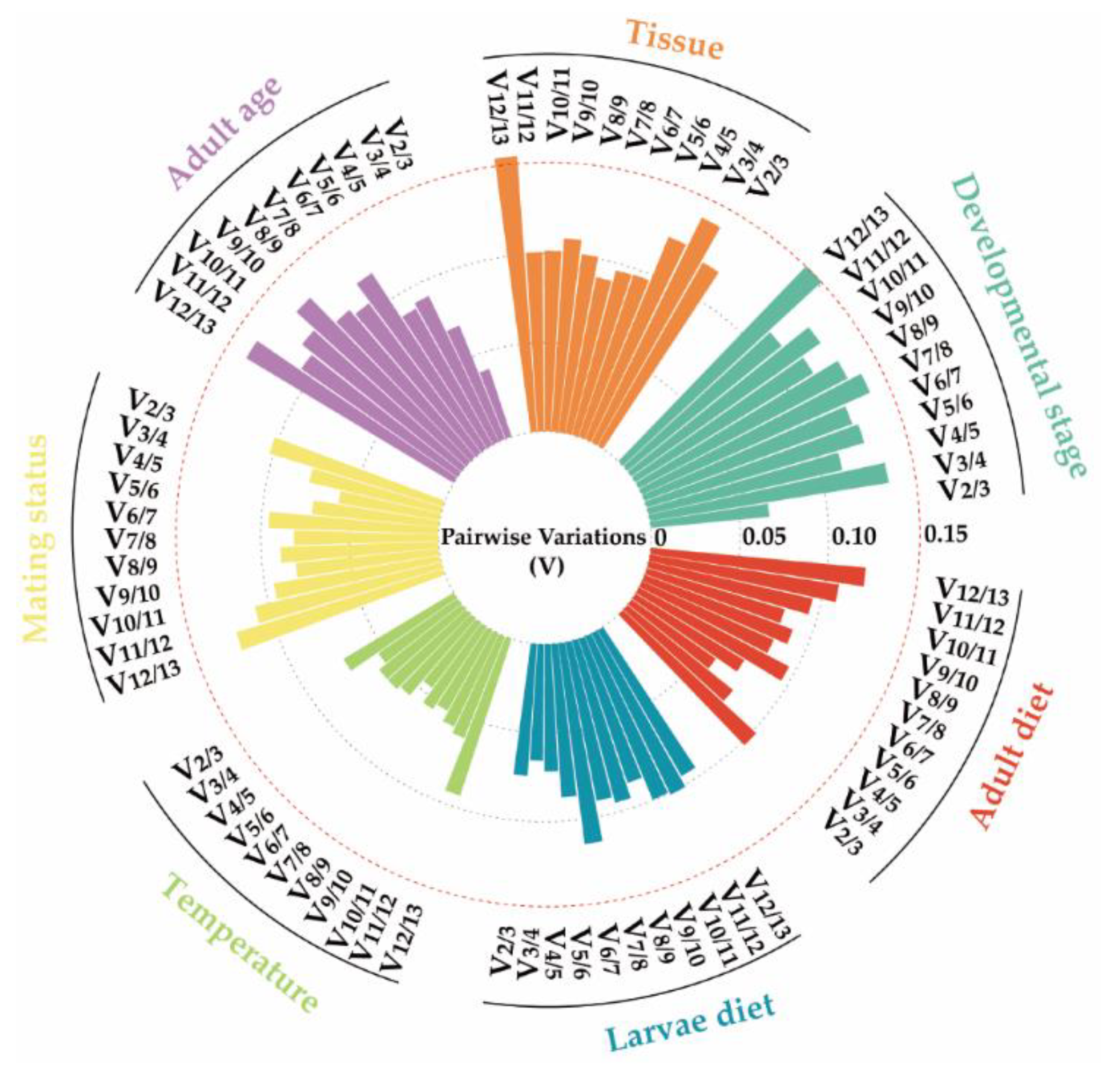
3.3. Stability of Reference Genes across Biotic Factors
3.4. Stability of Reference Genes under Abiotic Stresses
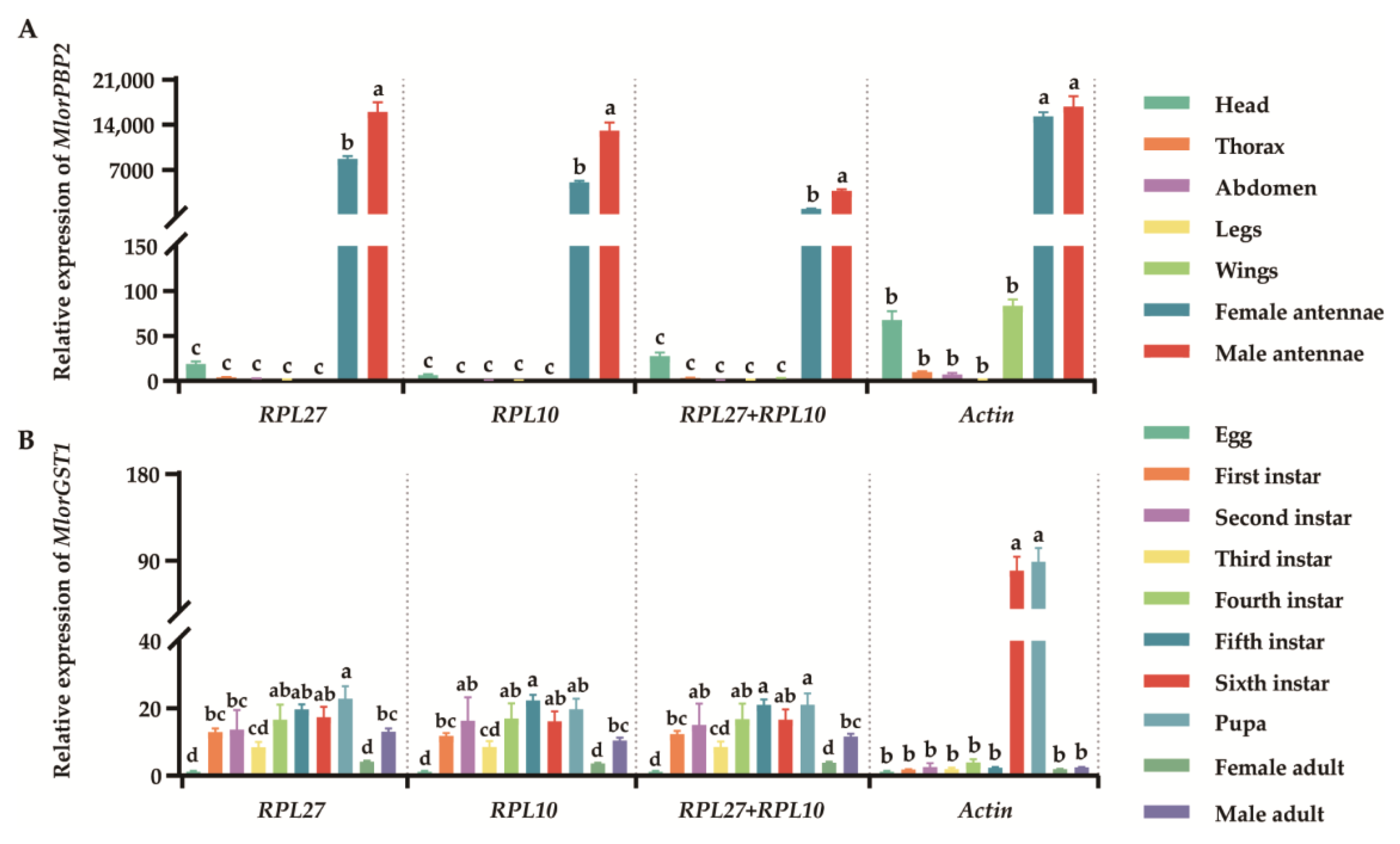
3.5. Validation of Candidate Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75–85. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Lü, J.; Yang, C.; Zhang, Y.; Pan, H. Selection of Reference Genes for the Normalization of RT-qPCR Data in Gene Expression Studies in Insects: A Systematic Review. Front. Physiol. 2018, 9, 1560. [Google Scholar] [CrossRef]
- Han, S.; Qin, Q.; Wang, D.; Zhou, Y.; He, Y. Selection and Evaluation of Reference Genes for qRT-PCR in Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2021, 12, 902. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Shi, D.; Wang, C.; Zhai, R.; Lyu, L.; He, Y.; Wang, D. Identification and Validation of Reference Genes for Expression Analysis Using qRT-PCR in Cimex hemipterus (Hemiptera: Cimicidae). Insects 2022, 13, 784. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, J.; Qiu, Z.; Hu, P.; Chen, X.; Yang, Z. Identification and Validation of Reference Genes for Expression Analysis Using RT-qPCR in Leptocybe invasa Fisher and La Salle (Hymenoptera: Eulophidae). Insects 2023, 14, 456. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Q.; Guo, P.; Gao, Z.; Chen, D.; Zhang, T.; Ning, J. Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in the Bean Bug, Riptortus pedestris (Hemiptera: Alydidae). Insects 2023, 14, 960. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, A.; Verma, S.; Muthamilarasan, M.; Rajam, M.V. Identification of suitable reference genes for expression profiling studies using qRT-PCR in an important insect pest, Maruca vitrata. Mol. Biol. Rep. 2021, 48, 7477–7485. [Google Scholar] [CrossRef]
- Fu, X.; Meyer-Rochow, V.B. Selection and Validation of Suitable Reference Genes for RT-qPCR Analysis in the Rare Aquatic Firefly Aquatica leii (Coleoptera: Lampyridae). Insects 2021, 12, 359. [Google Scholar] [CrossRef]
- Bansal, R.; Haviland, D.R.; Hunter, W.B. Selection and validation of reference genes for quantifying gene expression in the Gill’s mealybug. J. Econ. Entomol. 2023, 116, 2166–2172. [Google Scholar] [CrossRef]
- Nam, H.Y.; Kwon, M.; Kim, H.J.; Kim, J. Development of a Species Diagnostic Molecular Tool for an Invasive Pest, Mythimna loreyi, Using LAMP. Insects 2020, 11, 817. [Google Scholar] [CrossRef]
- Sertkaya, E.; Bayram, A. Parasitoid community of the loreyi leaf worm Mythimna (Acantholeucania) loreyi: Novel host-parasitoid associations and their efficiency in the eastern mediterranean region of Turkey. Phytoparasitica 2005, 33, 441–449. [Google Scholar] [CrossRef]
- Mediouni Ben Jemâa, J.; Soltani, A.; Djebbi, T.; Mejri, I.; Kanyesigye, D.; Otim, M.H. The Maize Caterpillar Mythimna (=Leucania) loreyi (Duponchel, 1827) (Lepidoptera: Noctuidae): Identification, Distribution, Population Density and Damage in Tunisia. Insects 2023, 14, 786. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Zhang, L.; Liu, Y.; Sappington, T.W.; Cheng, Y.; Luo, L.; Jiang, X. Population Projection and Development of the Mythimna loreyi (Lepidoptera: Noctuidae) as Affected by Temperature: Application of an Age-Stage, Two-Sex Life Table. J. Econ. Entomol. 2017, 110, 1583–1591. [Google Scholar] [CrossRef]
- Jiang, X.; Luo, L.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of Migration in Mythimna separata (Walker) in China: A Review Integrating Environmental, Physiological, Hormonal, Genetic, and Molecular Factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef]
- Feng, H.-Q.; Zhao, X.-C.; Wu, X.-F.; Wu, B.; Wu, K.-M.; Cheng, D.-F.; Guo, Y.-Y. Autumn Migration of Mythimna separata (Lepidoptera: Noctuidae) over the Bohai Sea in Northern China. Environ. Entomol. 2014, 37, 774–781. [Google Scholar] [CrossRef]
- Qin, J.; Liu, Y.; Zhang, L.; Cheng, Y.; Sappington, T.W.; Jiang, X. Effects of Moth Age and Rearing Temperature on the Flight Performance of the Loreyi Leafworm, Mythimna loreyi (Lepidoptera: Noctuidae), in Tethered and Free Flight. J. Econ. Entomol. 2018, 111, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Y.; Guo, J.-M.; Wei, Z.-Q.; Zhang, X.-T.; Liu, S.-R.; Guo, H.-F.; Dong, S.-L. Identification and sex expression profiles of olfactory-related genes in Mythimna loreyi based on antennal transcriptome analysis. J. Asia-Pac. Entomol. 2022, 25, 101934. [Google Scholar] [CrossRef]
- Wang, C.; Liu, L.; Huang, T.-Y.; Zhang, Y.; Liu, Y.; Wang, G.-R. Characterization of the pheromone receptors in Mythimna loreyi reveals the differentiation of sex pheromone recognition in Mythimna species. Insect Sci. 2023, 31, 173–185. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Bai, T.-F.; Guo, J.-M.; Wei, Z.-Q.; Liu, S.-R.; He, Y.; Ye, J.-J.; Yan, Q.; Zhang, J.; Dong, S.-L. Molecular mechanism of sex pheromone perception in male Mythimna loreyi revealed by in vitro system. Pest. Manag. Sci. 2024, 80, 744–755. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, H.; Liu, Y.; Wang, C.; Ma, B.; Zhang, M.; Zhang, Y.; Liu, Y.; Yang, B.; Wang, S.; et al. Chromosome-level genomes of two armyworms, Mythimna separata and Mythimna loreyi, provide insights into the biosynthesis and reception of sex pheromones. Mol. Ecol. Resour. 2023, 23, 1423–1441. [Google Scholar] [CrossRef]
- Shakeel, M.; Zhu, X.; Kang, T.; Wan, H.; Li, J. Selection and evaluation of reference genes for quantitative gene expression studies in cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2015, 18, 123–130. [Google Scholar] [CrossRef]
- Li, H.-B.; Dai, C.-G.; Zhang, C.-R.; He, Y.-F.; Ran, H.-Y.; Chen, S.-H. Screening potential reference genes for quantitative real-time PCR analysis in the oriental armyworm, Mythimna separata. PLoS ONE 2018, 13, e0195096. [Google Scholar] [CrossRef]
- Shen, C.-H.; Peng, L.-J.; Zhang, Y.-X.; Zeng, H.-R.; Yu, H.-F.; Jin, L.; Li, G.-Q. Reference Genes for Expression Analyses by qRT-PCR in Phthorimaea operculella (Lepidoptera: Gelechiidae). Insects 2022, 13, 140. [Google Scholar] [CrossRef]
- Pavlidi, N.; Vontas, J.; Van Leeuwen, T. The role of glutathione S-transferases (GSTs) in insecticide resistance in crop pests and disease vectors. Curr. Opin. Insect Sci. 2018, 27, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, R.; Zhou, T.; Cheng, S.; Li, C.; Ye, X.; Li, Y.; Tian, Z. Structural evidence for pheromone discrimination by the pheromone binding protein 3 from Plutella xylostella. Int. J. Biol. Macromol. 2021, 169, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Ran, H.; Liu, W.; Qu, Z.; Li, J. Mass rearing methods of Mythimna separata (Walker) and its parasitoid, Microplitis tuberculifer (Wesmael). J. Environ. Entomol. 2013, 35, 683–687. [Google Scholar]
- Zhang, Y.; Zhang, Y.; Yao, S.; Wang, G.; Wei, J.; Du, M.; An, S.; Yin, X. Supplemental Sugar Is Required for Sex Pheromone Biosynthesis in Mythimna separata. Front. Physiol. 2020, 11, 605145. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Wang, J.; Zhang, B. RefFinder: A web-based tool for comprehensively analyzing and identifying reference genes. Funct. Integr. Genom. 2023, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yuan, M.; Gao, X.; Kang, T.; Zhan, S.; Wan, H.; Li, J. Identification and Validation of Reference Genes for Gene Expression Analysis Using Quantitative PCR in Spodoptera litura (Lepidoptera: Noctuidae). PLoS ONE 2013, 8, e68059. [Google Scholar] [CrossRef]
- Zhang, S.; An, S.; Li, Z.; Wu, F.; Yang, Q.; Liu, Y.; Cao, J.; Zhang, H.; Zhang, Q.; Liu, X. Identification and validation of reference genes for normalization of gene expression analysis using qRT-PCR in Helicoverpa armigera (Lepidoptera: Noctuidae). Gene 2015, 555, 393–402. [Google Scholar] [CrossRef]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and Evaluation of Reference Genes for Expression Analysis Using qRT-PCR in the Beet Armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). PLoS ONE 2014, 9, e84730. [Google Scholar] [CrossRef]
- Xu, J.; Lu, M.-X.; Cui, Y.-D.; Du, Y.-Z. Selection and Evaluation of Reference Genes for Expression Analysis Using qRT-PCR in Chilo suppressalis (Lepidoptera: Pyralidae). J. Econ. Entomol. 2017, 110, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Rodriguez, A.; Tahir, U.B.; Jin, F. Gene expression studies of reference genes for quantitative real-time PCR: An overview in insects. Biotechnol. Lett. 2018, 40, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, Y.; Xu, K.; Wang, Y.; Yang, W. Selection and Validation of Reference Genes for Gene Expression Analysis in Tuta absoluta Meyrick (Lepidoptera: Gelechiidae). Insects 2021, 12, 589. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, J.; Lu, Y.; Sun, T.; Tian, J.; Huang, J.; Xu, H.; Wang, Z.; Lu, Z. Reference Genes for Expression Analysis Using RT-qPCR in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Insects 2022, 13, 1046. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Lu, M.-X.; Tang, X.-T.; Du, Y.-Z. Exploring Valid Reference Genes for Quantitative Real-Time PCR Analysis in Sesamia inferens (Lepidoptera: Noctuidae). PLoS ONE 2015, 10, e0115979. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-Y.; Li, Z.-L.; Lu, M.-X.; Cao, S.-S.; Du, Y.-Z. Selection of valid reference genes for quantitative real-time PCR in Cotesia chilonis (Hymenoptera: Braconidae) exposed to different temperatures. PLoS ONE 2019, 14, e0226139. [Google Scholar] [CrossRef]
- Tang, J.; Liang, G.; Dong, S.; Shan, S.; Zhao, M.; Guo, X. Selection and Validation of Reference Genes for Quantitative Real-Time PCR Normalization in Athetis dissimilis (Lepidoptera: Noctuidae) Under Different Conditions. Front. Physiol. 2022, 13, 842195. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.K.; Jain, G.; Upadhyay, S.K.; Sarita; Singh, H.; Dixit, S.; Verma, P.C. Reference genes validation in Phenacoccus solenopsis under various biotic and abiotic stress conditions. Sci. Rep. 2017, 7, 13520. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Zhang, J.; Cui, G.; Sun, R.; Sethuraman, V.; Yi, X.; Zhong, G. Evaluation of Reference Genes for Real-Time Quantitative PCR Analysis in Larvae of Spodoptera litura Exposed to Azadirachtin Stress Conditions. Front. Physiol. 2018, 9, 372. [Google Scholar] [CrossRef]
- Chapuis, M.-P.; Tohidi-Esfahani, D.; Dodgson, T.; Blondin, L.; Ponton, F.; Cullen, D.; Simpson, S.J.; Sword, G.A. Assessment and validation of a suite of reverse transcription-quantitative PCR reference genes for analyses of density-dependent behavioural plasticity in the Australian plague locust. BMC Mol. Biol. 2011, 12, 7. [Google Scholar] [CrossRef]
- Scharlaken, B.; de Graaf, D.C.; Goossens, K.; Brunain, M.; Peelman, L.J.; Jacobs, F.J. Reference gene selection for insect expression studies using quantitative real-time PCR: The head of the honeybee, Apis mellifera, after a bacterial challenge. J. Insect Sci. 2008, 8, 33. [Google Scholar] [CrossRef]
- Yang, A.-P.; Wang, Y.-S.; Huang, C.; Lv, Z.-C.; Liu, W.-X.; Bi, S.-Y.; Wan, F.-H.; Wu, Q.; Zhang, G.-F. Screening Potential Reference Genes in Tuta absoluta with Real-Time Quantitative PCR Analysis under Different Experimental Conditions. Genes 2021, 12, 1253. [Google Scholar] [CrossRef]
- De Jonge, H.J.M.; Fehrmann, R.S.N.; de Bont, E.S.J.M.; Hofstra, R.M.W.; Gerbens, F.; Kamps, W.A.; de Vries, E.G.E.; van der Zee, A.G.J.; te Meerman, G.J.; ter Elst, A. Evidence Based Selection of Housekeeping Genes. PLoS ONE 2007, 2, e898. [Google Scholar] [CrossRef] [PubMed]
- Omondi, B.A.; Latorre-Estivalis, J.M.; Rocha Oliveira, I.H.; Ignell, R.; Lorenzo, M.G. Evaluation of reference genes for insect olfaction studies. Parasites Vectors 2015, 8, 243. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequences (5′-3′, F/R) | Amplicon Length (bp) | Tm (°C) | Efficiency (%) | R2 |
|---|---|---|---|---|---|---|
| RPL10 | KAJ8725396.1 | ACCTGGTGTCTGATGAGTA | 295 | 55 | 97.2 | 0.994 |
| ATGACCTGAGCCTTCCAA | ||||||
| RPL27 | KAJ8722287.1 | TGAAGAACTACGACGAAGG | 187 | 55 | 97.7 | 0.990 |
| TCAACTGAGTAGCGAGTG | ||||||
| RPL32 | KAJ8705474.1 | CAATCGGATCGCTATGACA | 339 | 55 | 108.6 | 0.995 |
| TTATTCGTTCTCCTGGCTAC | ||||||
| RPS3 | KAJ8720489.1 | GTCCGCAAGAGGTTCAAT | 339 | 55 | 102.3 | 0.997 |
| CTTAATTCCGAGCACTCCT | ||||||
| TATA−box | KAJ8730825.1 | TCATACTCCTGTCCACTGT | 180 | 55 | 107.0 | 0.992 |
| TCTGTTCCACTCACCATTG | ||||||
| GAPDH | KAJ8714943.1 | GGCACAGTCGGTATAGAAG | 283 | 55 | 97.9 | 0.997 |
| AGGAAGCGTTGGAGATGA | ||||||
| AK | KAJ8736045.1 | CTGGTGTCGGAATCTACG | 108 | 55 | 102.3 | 0.995 |
| GCTTGTCGGTCTTCTTGA | ||||||
| Actin | KAJ8708612.1 | AATCGTGCGTGACATCAA | 473 | 55 | 95.3 | 0.999 |
| ACTCGTCGTATTCCTCCTT | ||||||
| EF | KAJ8731840.1 | ACACAGCTCGGATACAGT | 100 | 55 | 106.8 | 0.992 |
| GCATCAACCCAGTCGTTA | ||||||
| α−tubulin | KAJ8707351.1 | CGCTACCATCAAGACCAA | 257 | 55 | 107.7 | 0.996 |
| ACTCTCCTTCCTCCATACC | ||||||
| SOD | KAJ8723090.1 | CGAGTAATTGCGGTGTCA | 146 | 55 | 100.2 | 0.998 |
| CGTAGTCTTGCTCAGGTC | ||||||
| 18S rRNA | KAJ8712979.1 | ATCCAAGCACAGATGACAG | 464 | 55 | 103.5 | 0.998 |
| ACAACTACACCTCCATAGAAG | ||||||
| FTZ−F1 | KAJ8734564.1 | TAACAGACGGCACATCAC | 220 | 55 | 100.1 | 0.999 |
| TGTAAGGCACCAATGAGTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Yang, C.; Liu, Q.; Zhang, X.; Mei, X.; Zhang, T.; Ning, J. Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae). Insects 2024, 15, 185. https://doi.org/10.3390/insects15030185
Wang L, Yang C, Liu Q, Zhang X, Mei X, Zhang T, Ning J. Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae). Insects. 2024; 15(3):185. https://doi.org/10.3390/insects15030185
Chicago/Turabian StyleWang, Liuyang, Chaoxia Yang, Qingyu Liu, Xiaofang Zhang, Xiangdong Mei, Tao Zhang, and Jun Ning. 2024. "Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae)" Insects 15, no. 3: 185. https://doi.org/10.3390/insects15030185
APA StyleWang, L., Yang, C., Liu, Q., Zhang, X., Mei, X., Zhang, T., & Ning, J. (2024). Validation and Evaluation of Reference Genes for Quantitative Real-Time PCR Analysis in Mythimna loreyi (Lepidoptera: Noctuidae). Insects, 15(3), 185. https://doi.org/10.3390/insects15030185