
RNAi-Mediated Knockdown of Acidic Ribosomal Stalk Protein P1 Arrests Egg Development in Adult Female Yellow Fever Mosquitoes, Aedes aegypti



Abstract
Simple Summary
Abstract
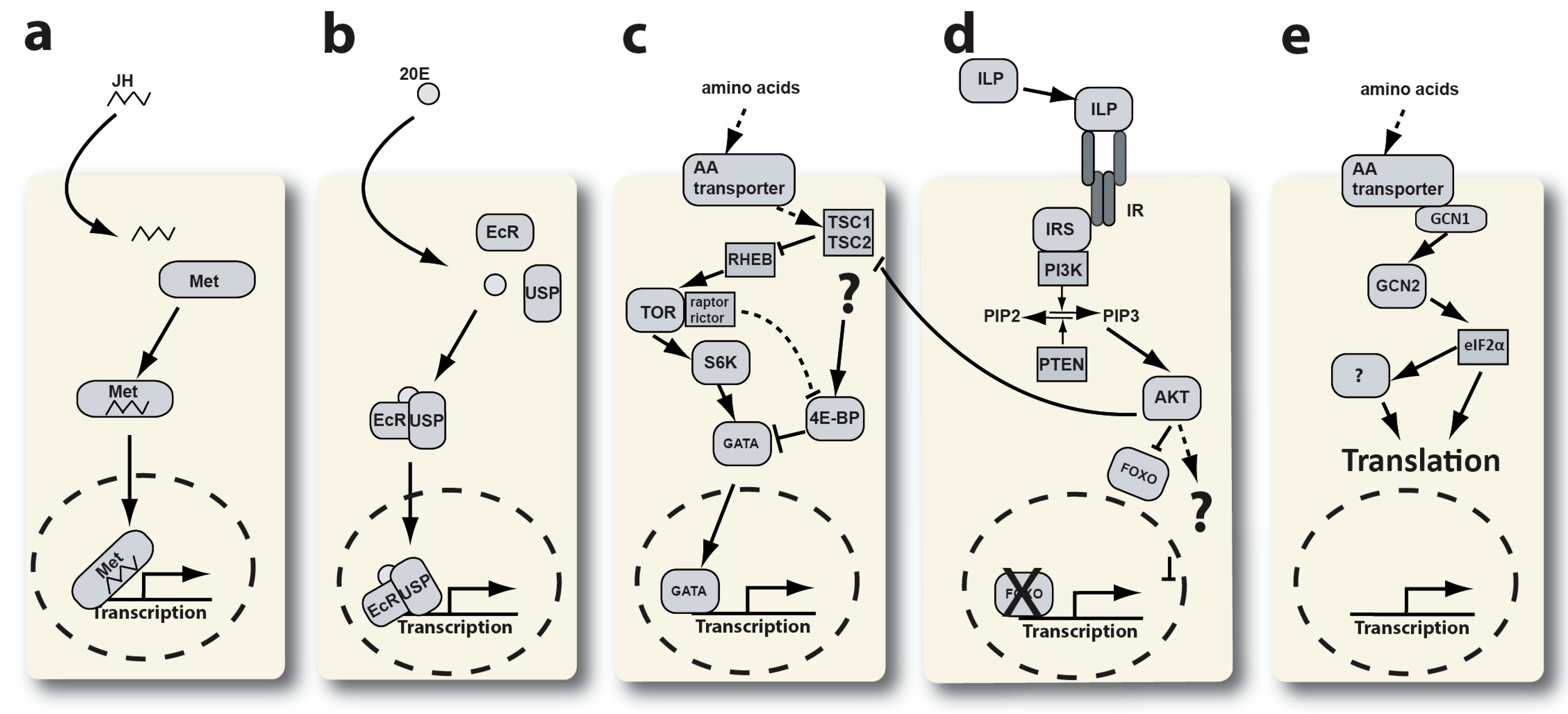
1. Introduction
2. Materials and Methods
3. Results
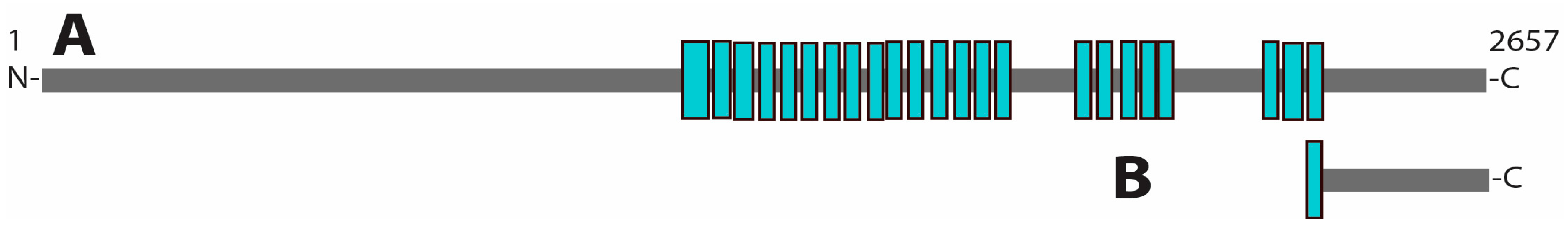
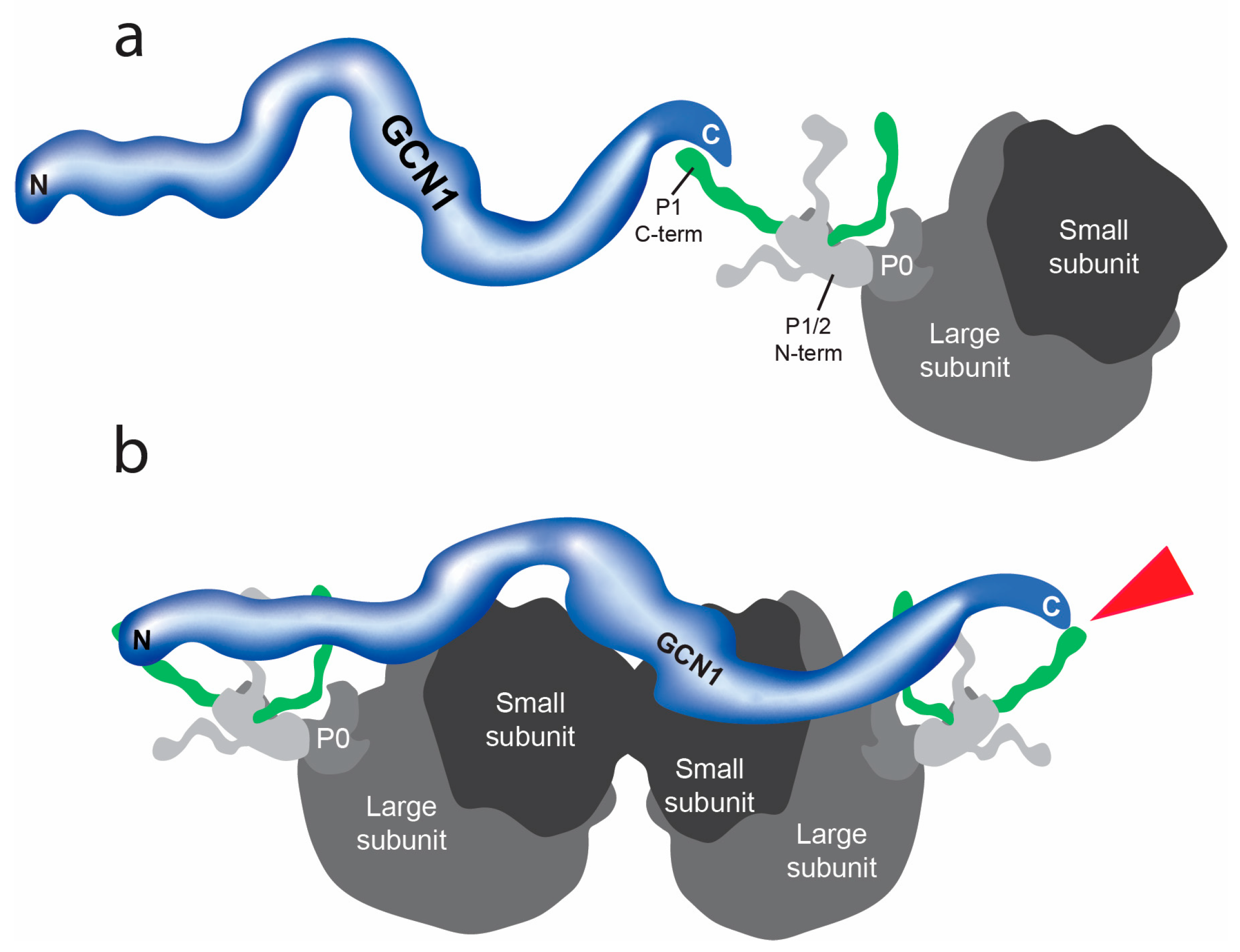
3.1. Protein–Protein Interactions of GCN1
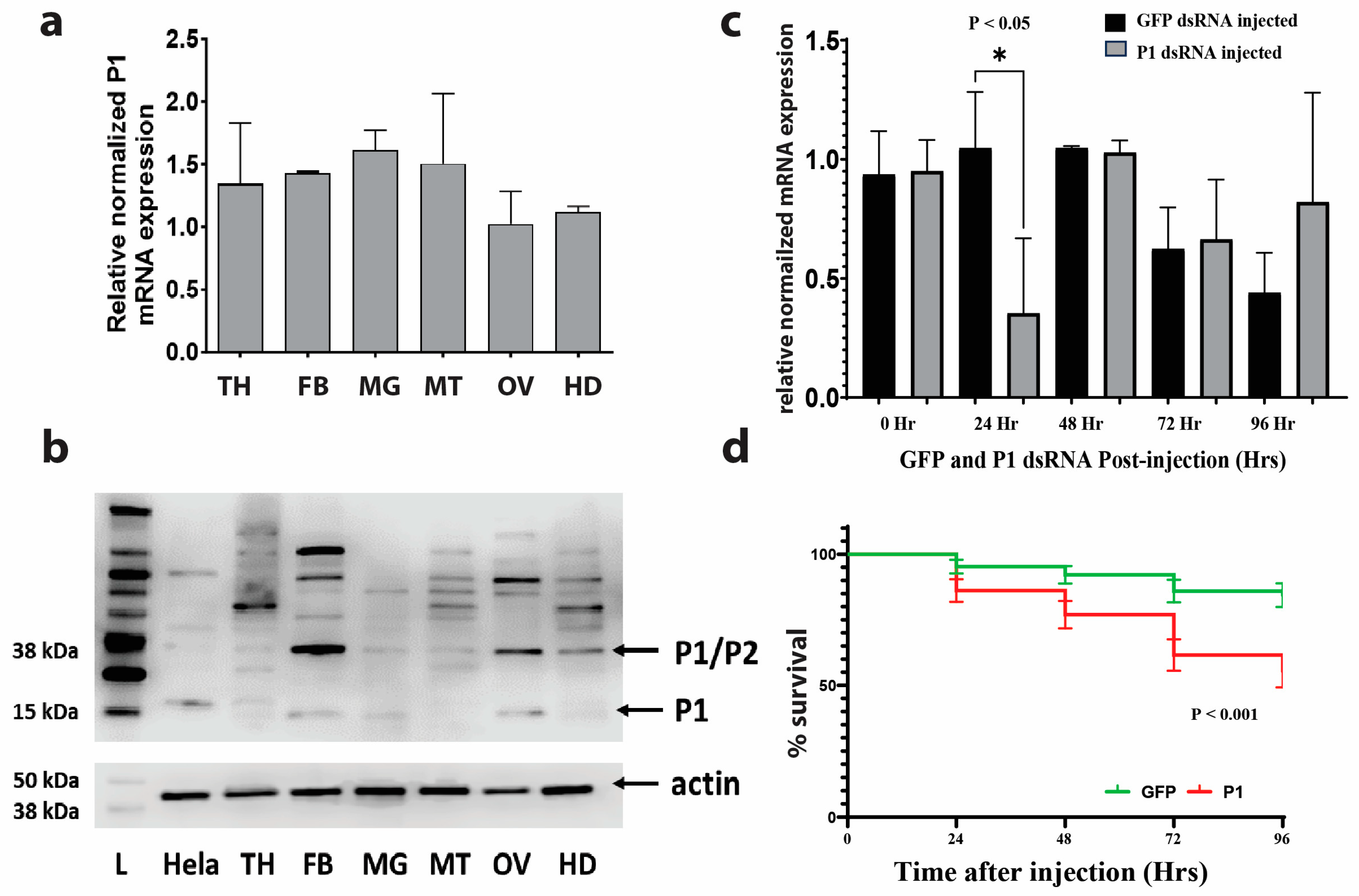
3.2. Gene Expression of P1 in Different Mosquito Organs and Structures
3.3. RNAi-Mediated Knockdown of P1 Is Transient
3.4. Mosquito Mortality after RNAi-Mediated P1 Knockdown
3.5. RNAi-Mediated P1 Knockdown Results in Ovary Phenotype in Mosquitoes
4. Discussion
5. Conclusions
6. Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, V.K.; Drake, L.L.; Aguirre, S.E.; Price, D.P.; Rodriguez, S.D.; Hansen, I.A. SLC7 amino acid transporters of the yellow fever mosquito Aedes aegypti and their role in fat body TOR signaling and reproduction. J. Insect Physiol. 2012, 58, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.J.; Vosshall, L.B. How to turn an organism into a model organism in 10 ‘easy’ steps. J. Exp. Biol. 2020, 223, jeb218198. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Chen, K.; Brown, M.R.; Strand, M.R. Multiple endocrine factors regulate nutrient mobilization and storage in Aedes aegypti during a gonadotrophic cycle. Insect Sci. 2023, 30, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Pinch, M.; Mitra, S.; Rodriguez, S.D.; Li, Y.; Kandel, Y.; Dungan, B.; Holguin, F.O.; Attardo, G.M.; Hansen, I.A. Fat and happy: Profiling mosquito fat body lipid storage and composition post-blood meal. Front. Insect Sci. 2021, 1, 693168. [Google Scholar] [CrossRef]
- Clemons, A.; Haugen, M.; Flannery, E.; Tomchaney, M.; Kast, K.; Jacowski, C.; Le, C.; Mori, A.; Holland, W.S.; Sarro, J. Aedes aegypti: An emerging model for vector mosquito development. Cold Spring Harb. Protoc. 2010, 2010, pdb.emo141. [Google Scholar] [CrossRef] [PubMed]
- Gulia-Nuss, M.; Elliot, A.; Brown, M.R.; Strand, M.R. Multiple factors contribute to anautogenous reproduction by the mosquito Aedes aegypti. J. Insect Physiol. 2015, 82, 8–16. [Google Scholar] [CrossRef]
- Attardo, G.M.; Higgs, S.; Klingler, K.A.; Vanlandingham, D.L.; Raikhel, A.S. RNA interference-mediated knockdown of a GATA factor reveals a link to anautogeny in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2003, 100, 13374–13379. [Google Scholar] [CrossRef]
- Ariani, C.V.; Smith, S.C.; Osei-Poku, J.; Short, K.; Juneja, P.; Jiggins, F.M. Environmental and genetic factors determine whether the mosquito Aedes aegypti lays eggs without a blood meal. Am. J. Trop. Med. Hyg. 2015, 92, 715. [Google Scholar] [CrossRef]
- Valle, D. Vitellogenesis in insects and other groups: A review. Mem. Inst. Oswaldo Cruz 1993, 88, 1–26. [Google Scholar] [CrossRef]
- Roth, T.F.; Porter, K.R. Yolk protein uptake in the oocyte of the mosquito Aedes aegypti. L. J. Cell Biol. 1964, 20, 313–332. [Google Scholar] [CrossRef]
- Snigirevskaya, E.S.; Hays, A.R.; Raikhel, A.S. Secretory and internalization pathways of mosquito yolk protein precursors. Cell Tissue Res. 1997, 290, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Sappington, T.W.; Kokoza, V.A.; Cho, W.-L.; Raikhel, A.S. Molecular characterization of the mosquito vitellogenin receptor reveals unexpected high homology to the Drosophila yolk protein receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 8934–8939. [Google Scholar] [CrossRef] [PubMed]
- Bownes, M.; Shirras, A.; Blair, M.; Collins, J.; Coulson, A. Evidence that insect embryogenesis is regulated by ecdysteroids released from yolk proteins. Proc. Natl. Acad. Sci. USA 1988, 85, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Townson, H. The biology of mosquitoes. Volume 1. Development, nutrition and reproduction. By A.N. Clements. (London: Chapman & Hall, 1992). viii + 509 pp. Hard cover £50. ISBN 0-412-40180-0. Bull. Entomol. Res. 1993, 83, 307–308. [Google Scholar] [CrossRef]
- Roy, S.G.; Raikhel, A.S. The small GTPase Rheb is a key component linking amino acid signaling and TOR in the nutritional pathway that controls mosquito egg development. Insect Biochem. Mol. Biol. 2011, 41, 62–69. [Google Scholar] [CrossRef]
- Avruch, J.; Long, X.; Ortiz-Vega, S.; Rapley, J.; Papageorgiou, A.; Dai, N. Amino acid regulation of TOR complex 1. Am. J. Physiol.-Endocrinol. Metab. 2009, 296, E592–E602. [Google Scholar] [CrossRef]
- Smykal, V.; Raikhel, A.S. Nutritional control of insect reproduction. Curr. Opin. Insect Sci. 2015, 11, 31–38. [Google Scholar] [CrossRef]
- Ling, L.; Raikhel, A.S. Cross-talk of insulin-like peptides, juvenile hormone, and 20-hydroxyecdysone in regulation of metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2021, 118, e2023470118. [Google Scholar] [CrossRef]
- Hansen, I.A.; Attardo, G.M.; Rodriguez, S.D.; Drake, L.L. Four-way regulation of mosquito yolk protein precursor genes by juvenile hormone-, ecdysone-, nutrient-, and insulin-like peptide signaling pathways. Front. Physiol. 2014, 5, 103. [Google Scholar] [CrossRef]
- Hundal, H.S.; Taylor, P.M. Amino acid transceptors: Gate keepers of nutrient exchange and regulators of nutrient signaling. Am. J. Physiol.-Endocrinol. Metab. 2009, 296, E603–E613. [Google Scholar] [CrossRef]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Fotiadis, D.; Kanai, Y.; Palacin, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Asp. Med. 2013, 34, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Verrey, F.; Closs, E.I.; Wagner, C.A.; Palacin, M.; Endou, H.; Kanai, Y. CATs and HATs: The SLC7 family of amino acid transporters. Pflug. Arch. 2004, 447, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Hansen, I.A.; Shiao, S.-H.; Raikhel, A.S. Identification of two cationic amino acid transporters required for nutritional signaling during mosquito reproduction. J. Exp. Biol. 2006, 209, 3071–3078. [Google Scholar] [CrossRef]
- Boudko, D.Y.; Tsujimoto, H.; Rodriguez, S.D.; Meleshkevitch, E.A.; Price, D.P.; Drake, L.L.; Hansen, I.A. Substrate specificity and transport mechanism of amino-acid transceptor Slimfast from Aedes aegypti. Nat. Commun. 2015, 6, 8546. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.A.; Boudko, D.Y.; Shiao, S.-H.; Voronov, D.A.; Meleshkevitch, E.A.; Drake, L.L.; Aguirre, S.E.; Fox, J.M.; Attardo, G.M.; Raikhel, A.S. AaCAT1 of the yellow fever mosquito, Aedes aegypti: A novel histidine-specific amino acid transporter from the SLC7 family. J. Biol. Chem. 2011, 286, 10803–10813. [Google Scholar] [CrossRef] [PubMed]
- Staschke, K.A.; Dey, S.; Zaborske, J.M.; Palam, L.R.; McClintick, J.N.; Pan, T.; Edenberg, H.J.; Wek, R.C. Integration of general amino acid control and target of rapamycin (TOR) regulatory pathways in nitrogen assimilation in yeast. J. Biol. Chem. 2010, 285, 16893–16911. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, P.T.; Hay, N. The two TORCs and AKT. Dev. Cell 2007, 12, 487–502. [Google Scholar] [CrossRef]
- Battu, S.; Minhas, G.; Mishra, A.; Khan, N. Amino acid sensing via general control nonderepressible-2 kinase and immunological programming. Front. Immunol. 2017, 8, 1719. [Google Scholar] [CrossRef]
- Kubota, H.; Ota, K.; Sakaki, Y.; Ito, T. Budding yeast GCN1 binds the GI domain to activate the eIF2α kinase GCN2. J. Biol. Chem. 2001, 276, 17591–17596. [Google Scholar] [CrossRef]
- Gottfried, S.; Koloamatangi, S.M.; Daube, C.; Schiemann, A.H.; Sattlegger, E. A genetic approach to identify amino acids in Gcn1 required for Gcn2 activation. PLoS ONE 2022, 17, e0277648. [Google Scholar] [CrossRef] [PubMed]
- Sattlegger, E.; Hinnebusch, A.G. Polyribosome binding by GCN1 is required for full activation of eukaryotic translation initiation factor 2α kinase GCN2 during amino acid starvation. J. Biol. Chem. 2005, 280, 16514–16521. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zou, Y.; Mao, D.; Sun, D.; Gao, G.; Shi, J.; Liu, X.; Zhu, C.; Yang, M.; Ye, W. The general amino acid control pathway regulates mTOR and autophagy during serum/glutamine starvation. J. Cell Biol. 2014, 206, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Raina, M.; Ibba, M. tRNAs as regulators of biological processes. Front. Genet. 2014, 5, 171. [Google Scholar] [CrossRef] [PubMed]
- Pinch, M.; Muka, T.; Kandel, Y.; Lamsal, M.; Martinez, N.; Teixeira, M.; Boudko, D.Y.; Hansen, I.A. General control nonderepressible 1 interacts with cationic amino acid transporter 1 and affects Aedes aegypti fecundity. Parasites Vectors 2022, 15, 383. [Google Scholar] [CrossRef] [PubMed]
- Castilho, B.A.; Shanmugam, R.; Silva, R.C.; Ramesh, R.; Himme, B.M.; Sattlegger, E. Keeping the eIF2 alpha kinase Gcn2 in check. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 1948–1968. [Google Scholar] [CrossRef]
- Marton, M.; Crouch, D.; Hinnebusch, A. GCN1, a translational activator of GCN4 in Saccharomyces cerevisiae, is required for phosphorylation of eukaryotic translation initiation factor 2 by protein kinase GCN2. Mol. Cell. Biol. 1993, 13, 3541–3556. [Google Scholar]
- Marton, M.J.; Vazquez de Aldana, C.R.; Qiu, H.; Chakraburtty, K.; Hinnebusch, A.G. Evidence that GCN1 and GCN20, translational regulators of GCN4, function on elongating ribosomes in activation of eIF2alpha kinase GCN2. Mol. Cell. Biol. 1997, 17, 4474–4489. [Google Scholar] [CrossRef]
- Inglis, A.J.; Masson, G.R.; Shao, S.; Perisic, O.; McLaughlin, S.H.; Hegde, R.S.; Williams, R.L. Activation of GCN2 by the ribosomal P-stalk. Proc. Natl. Acad. Sci. USA 2019, 116, 4946–4954. [Google Scholar] [CrossRef]
- Lee, S.J.; Swanson, M.J.; Sattlegger, E. Gcn1 contacts the small ribosomal protein Rps10, which is required for full activation of the protein kinase Gcn2. Biochem. J. 2015, 466, 547–559. [Google Scholar] [CrossRef]
- Sattlegger, E.; Hinnebusch, A.G. Separate domains in GCN1 for binding protein kinase GCN2 and ribosomes are required for GCN2 activation in amino acid-starved cells. EMBO J. 2000, 19, 6622–6633. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.H.; Hirano, T. HEAT repeats–versatile arrays of amphiphilic helices working in crowded environments? J. Cell Sci. 2016, 129, 3963–3970. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, D.; Marintchev, A.; Arthanari, H. The metaphorical swiss army knife: The multitude and diverse roles of HEAT domains in eukaryotic translation initiation. Nucleic Acids Res. 2022, 50, 5424–5442. [Google Scholar] [CrossRef] [PubMed]
- Rakesh, R.; Krishnan, R.; Sattlegger, E.; Srinivasan, N. Recognition of a structural domain (RWDBD) in Gcn1 proteins that interacts with the RWD domain containing proteins. Biol. Direct 2017, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E. Determination of a physiological saline solution for Aedes aegypti (L.). J. Econ. Entomol. 1953, 46, 624–627. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Drake, L.L.; Price, D.P.; Aguirre, S.E.; Hansen, I.A. RNAi-mediated gene knockdown and in vivo diuresis assay in adult female Aedes aegypti mosquitoes. J. Vis Exp. 2012, e3479. [Google Scholar] [CrossRef]
- Dzaki, N.; Ramli, K.N.; Azlan, A.; Ishak, I.H.; Azzam, G. Evaluation of reference genes at different developmental stages for quantitative real-time PCR in Aedes aegypti. Sci. Rep. 2017, 7, 43618. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Calzolari, M. Mosquito-borne diseases in Europe: An emerging public health threat. Rep. Parasitol. 2016, 5, 1–12. [Google Scholar] [CrossRef]
- Patterson, J.; Sammon, M.; Garg, M. Dengue, Zika and chikungunya: Emerging arboviruses in the New World. West. J. Emerg. Med. 2016, 17, 671. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, D. Outbreak of locally acquired mosquito-transmitted (autochthonous) malaria—Florida and Texas, May–July 2023. MMWR. Morb. Mortal. Wkly. Rep. 2023, 72, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Anoopkumar, A.; Aneesh, E.M. A critical assessment of mosquito control and the influence of climate change on mosquito-borne disease epidemics. Environ. Dev. Sustain. 2022, 24, 8900–8929. [Google Scholar] [CrossRef]
- Jones, R.T.; Ant, T.H.; Cameron, M.M.; Logan, J.G. Novel control strategies for mosquito-borne diseases. Philos. Trans. R. Soc. B 2021, 376, 20190802. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological control of mosquito vectors: Past, present, and future. Insects 2016, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.; Cardoso, J.C.; Felix, R.C.; Santana, R.A.; Guerra, M.d.G.B.; Power, D.; Silveira, H. Fresh-blood-free diet for rearing malaria mosquito vectors. Sci. Rep. 2018, 8, 17807. [Google Scholar] [CrossRef]
- Clements, A.N. The Biology of Mosquitoes; Chapman & Hall: London, UK, 1992; Volume 1. [Google Scholar]
- Perdomo, H.D.; Hussain, M.; Parry, R.; Etebari, K.; Hedges, L.M.; Zhang, G.; Schulz, B.L.; Asgari, S. Human blood microRNA hsa-miR-21-5p induces vitellogenin in the mosquito Aedes aegypti. Commun. Biol. 2021, 4, 856. [Google Scholar] [CrossRef]
- Hansen, I.A.; Attardo, G.M.; Park, J.H.; Peng, Q.; Raikhel, A.S. Target of rapamycin-mediated amino acid signaling in mosquito anautogeny. Proc. Natl. Acad. Sci. USA 2004, 101, 10626–10631. [Google Scholar] [CrossRef]
- Hansen, I.A.; Attardo, G.M.; Roy, S.G.; Raikhel, A.S. Target of rapamycin-dependent activation of S6 kinase is a central step in the transduction of nutritional signals during egg development in a mosquito. J. Biol. Chem. 2005, 280, 20565–20572. [Google Scholar] [CrossRef]
- Remacha, M.; Jimenez-Diaz, A.; Santos, C.; Briones, E.; Zambrano, R.; Gabriel, M.R.; Guarinos, E.; Ballesta, J.P. Proteins P1, P2, and P0, components of the eukaryotic ribosome stalk. New structural and functional aspects. Biochem. Cell Biol. 1995, 73, 959–968. [Google Scholar] [CrossRef]
- Tchórzewski, M. The acidic ribosomal P proteins. Int. J. Biochem. Cell Biol. 2002, 34, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Hinnebusch, A.G. Differential requirements for P stalk components in activating yeast protein kinase Gcn2 by stalled ribosomes during stress. Proc. Natl. Acad. Sci. USA 2023, 120, e2300521120. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-M.; Yu, C.W.-H.; Chan, D.S.-B.; Chiu, T.Y.-H.; Zhu, G.; Sze, K.-H.; Shaw, P.-C.; Wong, K.-B. Solution structure of the dimerization domain of ribosomal protein P2 provides insights for the structural organization of eukaryotic stalk. Nucleic Acids Res. 2010, 38, 5206–5216. [Google Scholar] [CrossRef]
- Kulczyk, A.W.; Sorzano, C.O.S.; Grela, P.; Tchórzewski, M.; Tumer, N.E.; Li, X.-P. Cryo-EM structure of Shiga toxin 2 in complex with the native ribosomal P-stalk reveals residues involved in the binding interaction. J. Biol. Chem. 2023, 299, 102795. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.K.; Wong, B.; Xie, X.; Lu, Y.-F.; Shi, P.-Y.; Pompon, J.; Garcia-Blanco, M.A.; Bradrick, S.S. RPLP1 and RPLP2 are essential flavivirus host factors that promote early viral protein accumulation. J. Virol. 2017, 91, e01706-16. [Google Scholar] [CrossRef]
- Tchórzewski, M.; Boldyreff, B.; Issinger, O.-G.; Grankowski, N. Analysis of the protein–protein interactions between the human acidic ribosomal P-proteins: Evaluation by the two hybrid system. Int. J. Biochem. Cell Biol. 2000, 32, 737–746. [Google Scholar] [CrossRef]
- Wahl, M.C.; Moller, W. Structure and function of the acidic ribosomal stalk proteins. Curr. Protein Pept. Sci. 2002, 3, 93–106. [Google Scholar] [CrossRef]
- Ballesta, J.P.; Remacha, M. The large ribosomal subunit stalk as a regulatory element of the eukaryotic translational machinery. Prog. Nucleic Acid Res. Mol. Biol. 1996, 55, 157–193. [Google Scholar]
- Wu, C.C.-C.; Peterson, A.; Zinshteyn, B.; Regot, S.; Green, R. Ribosome collisions trigger general stress responses to regulate cell fate. Cell 2020, 182, 404–416.e414. [Google Scholar] [CrossRef]
- Ryder, L.; Arendrup, F.S.; Martínez, J.F.; Snieckute, G.; Pecorari, C.; Shah, R.A.; Lund, A.H.; Blasius, M.; Bekker-Jensen, S. Nitric oxide-induced ribosome collision activates ribosomal surveillance mechanisms. Cell Death Dis. 2023, 14, 467. [Google Scholar] [CrossRef]
- Vind, A.C.; Snieckute, G.; Bekker-Jensen, S.; Blasius, M. Run, Ribosome, Run: From compromised translation to human health. Antioxidants Redox Signal. 2023, 39, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Pochopien, A.A.; Beckert, B.; Kasvandik, S.; Berninghausen, O.; Beckmann, R.; Tenson, T.; Wilson, D.N. Structure of Gcn1 bound to stalled and colliding 80S ribosomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2022756118. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.K.; Wijeratne, H.S.; Shah, P.; Garcia-Blanco, M.A.; Bradrick, S.S. Ribosomal stalk proteins RPLP1 and RPLP2 promote biogenesis of flaviviral and cellular multi-pass transmembrane proteins. Nucleic Acids Res. 2020, 48, 9872–9885. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, E.; Levanon, E.Y. Human housekeeping genes, revisited. Trends Genet. 2013, 29, 569–574. [Google Scholar] [CrossRef]
- Martinez, N.P.; Pinch, M.; Kandel, Y.; Hansen, I.A. Knockdown of the Sodium/Potassium ATPase Subunit Beta 2 Reduces Egg Production in the Dengue Vector. Aedes aegypti. Insects 2023, 14, 50. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Product Size (bp) |
|---|---|---|
| P1 forward | TAATACGACTCACTATAGGGAG ATGTTTTTCGTCATTGG | 363 |
| P1 reverse | TAATACGACTCACTATAGGGAG TCAACCTGCGATTCC | |
| GFP forward | TAATACGACTCACTATAGGG CGATGCCACCT | 518 |
| GFP reverse | TAATACGACTCACTATAGGG CGGACTGGGTG |
| Primer | Sequence | Product Size (bp) |
|---|---|---|
| P1 forward | CGACGATGTCGCTGTGACCG | 95 |
| P1 reverse | CCTTGACGAACAGAGCGGGC | |
| β-actin * forward | GACTACCTGATGAAGATCCTGAC | 93 |
| β-actin * reverse | GCACAGCTTCTCCTTAATGTCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamsal, M.; Luker, H.A.; Pinch, M.; Hansen, I.A. RNAi-Mediated Knockdown of Acidic Ribosomal Stalk Protein P1 Arrests Egg Development in Adult Female Yellow Fever Mosquitoes, Aedes aegypti. Insects 2024, 15, 84. https://doi.org/10.3390/insects15020084
Lamsal M, Luker HA, Pinch M, Hansen IA. RNAi-Mediated Knockdown of Acidic Ribosomal Stalk Protein P1 Arrests Egg Development in Adult Female Yellow Fever Mosquitoes, Aedes aegypti. Insects. 2024; 15(2):84. https://doi.org/10.3390/insects15020084
Chicago/Turabian StyleLamsal, Mahesh, Hailey A. Luker, Matthew Pinch, and Immo A. Hansen. 2024. "RNAi-Mediated Knockdown of Acidic Ribosomal Stalk Protein P1 Arrests Egg Development in Adult Female Yellow Fever Mosquitoes, Aedes aegypti" Insects 15, no. 2: 84. https://doi.org/10.3390/insects15020084
APA StyleLamsal, M., Luker, H. A., Pinch, M., & Hansen, I. A. (2024). RNAi-Mediated Knockdown of Acidic Ribosomal Stalk Protein P1 Arrests Egg Development in Adult Female Yellow Fever Mosquitoes, Aedes aegypti. Insects, 15(2), 84. https://doi.org/10.3390/insects15020084