
Integrated Pest Management Strategies to Control Varroa Mites and Their Effect on Viral Loads in Honey Bee Colonies

 , , , ,
, , , ,  ,
, 
Abstract
Simple Summary
Abstract
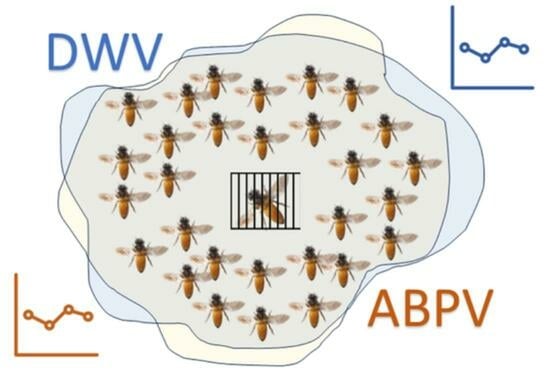
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
- Mengeško polje apiary (SI): located in the central Slovenian region (46.146453, 14.575566), characterized by a continental climate. Twenty-two colonies were included in the trials, housed on 10 frames (260 mm × 410 mm). Trials were conducted during the summer season. Day 0 of the protocol was on 12 July 2018, oxalic acid was administered on 6 August 2018, and follow-up treatment began on 21 August 2018.
- Ciampino apiary (IT): located in the Latium region in central Italy (41.808058, 12.613931), characterized by Mediterranean climate. Here, 26 colonies were housed on 10 Dadant Blatt frame (290 mm × 430 mm) hives. Trials were conducted during the late summer/early autumn seasons. Day 0 of the protocol was on 20 August 2018, oxalic acid was administered on 14 September 2018, and follow-up treatment began on 28 September 2018.
2.2. Experimental Setup and Treatments
- “Queen Caging” (QC) group: queens belonging to this group were caged in VAR-CONTROL® cages (Api-Mo.Bru, Campodoro, Padova, Italy) (Figure 1) from day 0 to day 24. In the SI experimental group, there were 6 colonies, and there were 8 colonies in the IT group.
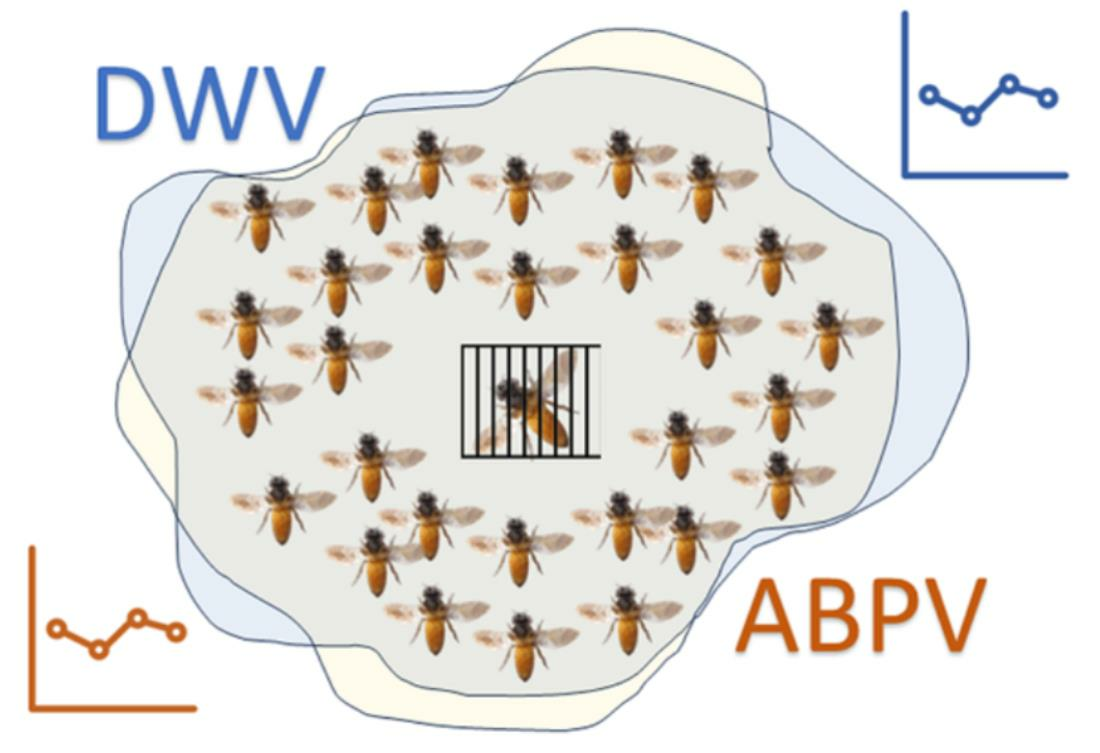
- “Trapping comb” (TC) group: queens in this group were caged on a trapping comb (Figure 2) on day 0. On day 20 of the protocol, the queens were transferred into a VAR-CONTROL® cage to evaluate the residual mite fall, and the trapping comb with the capped brood was removed. There were 6 colonies in the SI experimental group and 5 colonies in the IT group.
- The “Control 1” (CG1) group was established to evaluate the strength of the colonies (SI location: 5 colonies, IT location: 8 colonies).
- The “Control 2” (CG2) group was established to monitor the natural mite fall (SI location: 5 colonies, IT location: 5 colonies). Queens were caged on day 40 to evaluate the total number of mites killed by follow-up treatment.
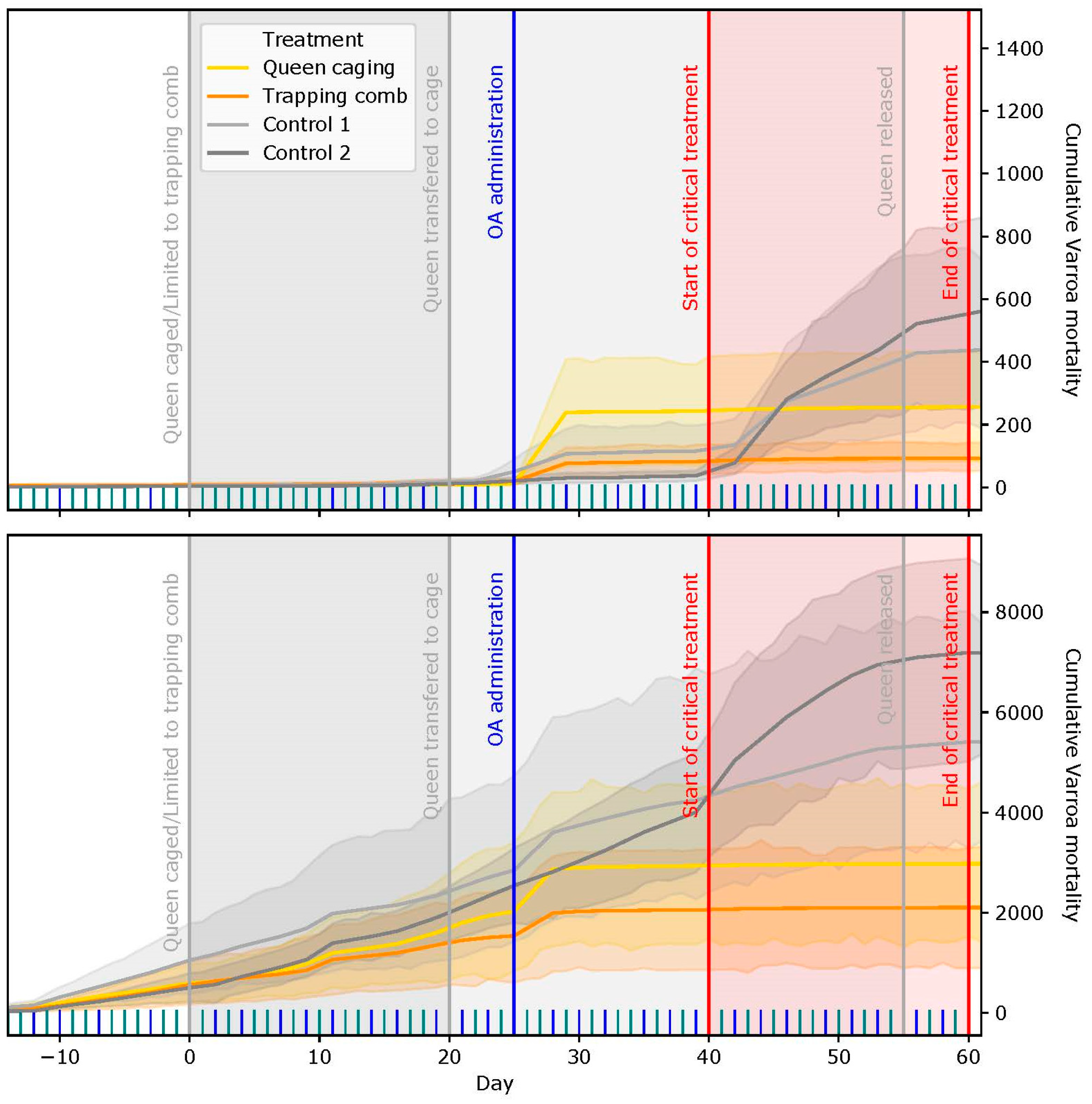
2.3. Mite Fall
2.4. Colony Strength
- -
- On day 7, to create homogenous groups;
- -
- On day 25, to evaluate the impact of the brood interruption techniques;
- -
- On day 40, to evaluate the impact of the oxalic acid treatment.
2.5. Viral Load
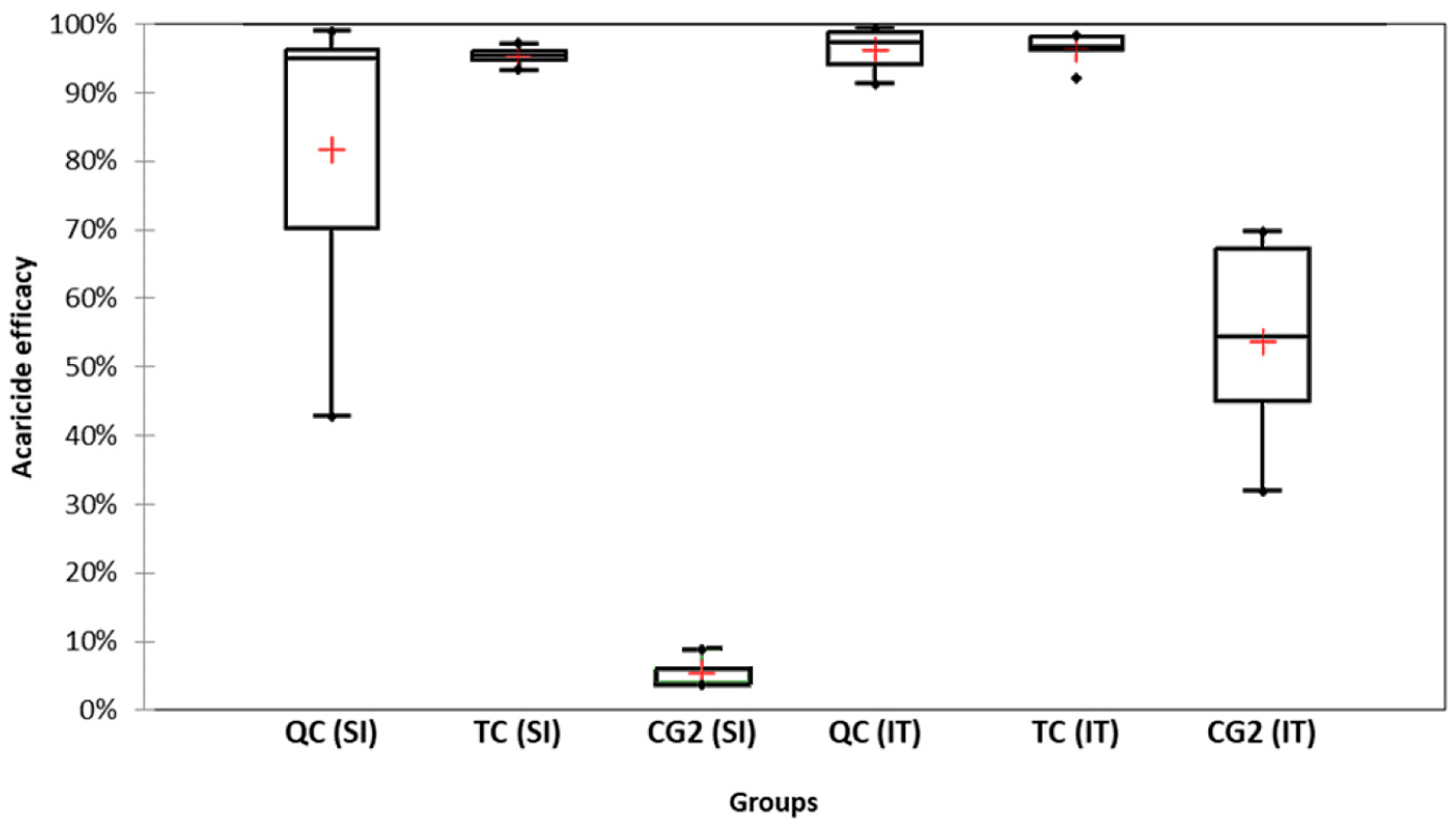
2.6. Acaricide Efficacy
2.7. Statistical Analyses
3. Results
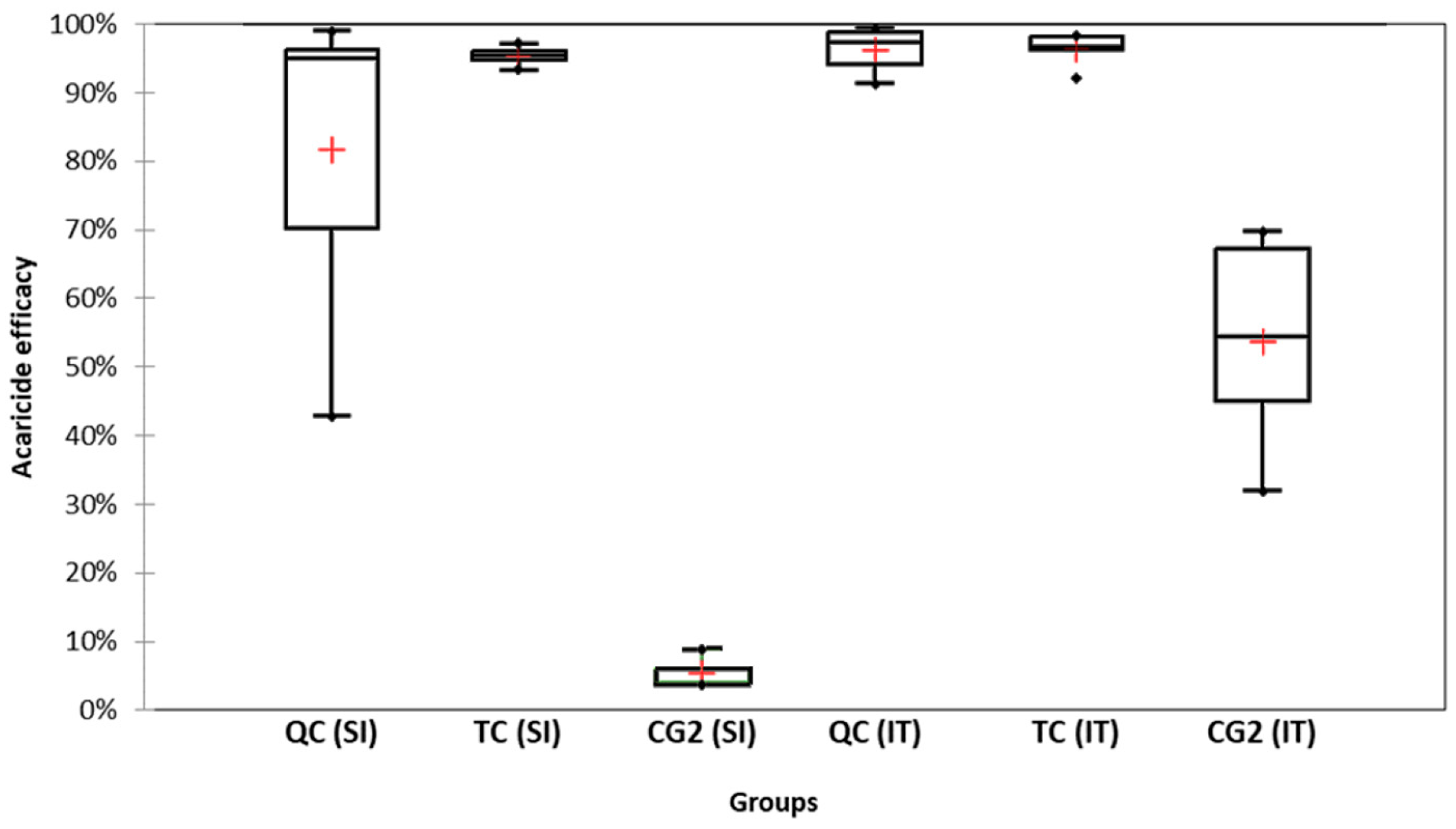
3.1. Acaricide Efficacy and Cumulative Mite Fall
3.1.1. Slovenian (SI) Apiary
3.1.2. Italian (IT) Apiary
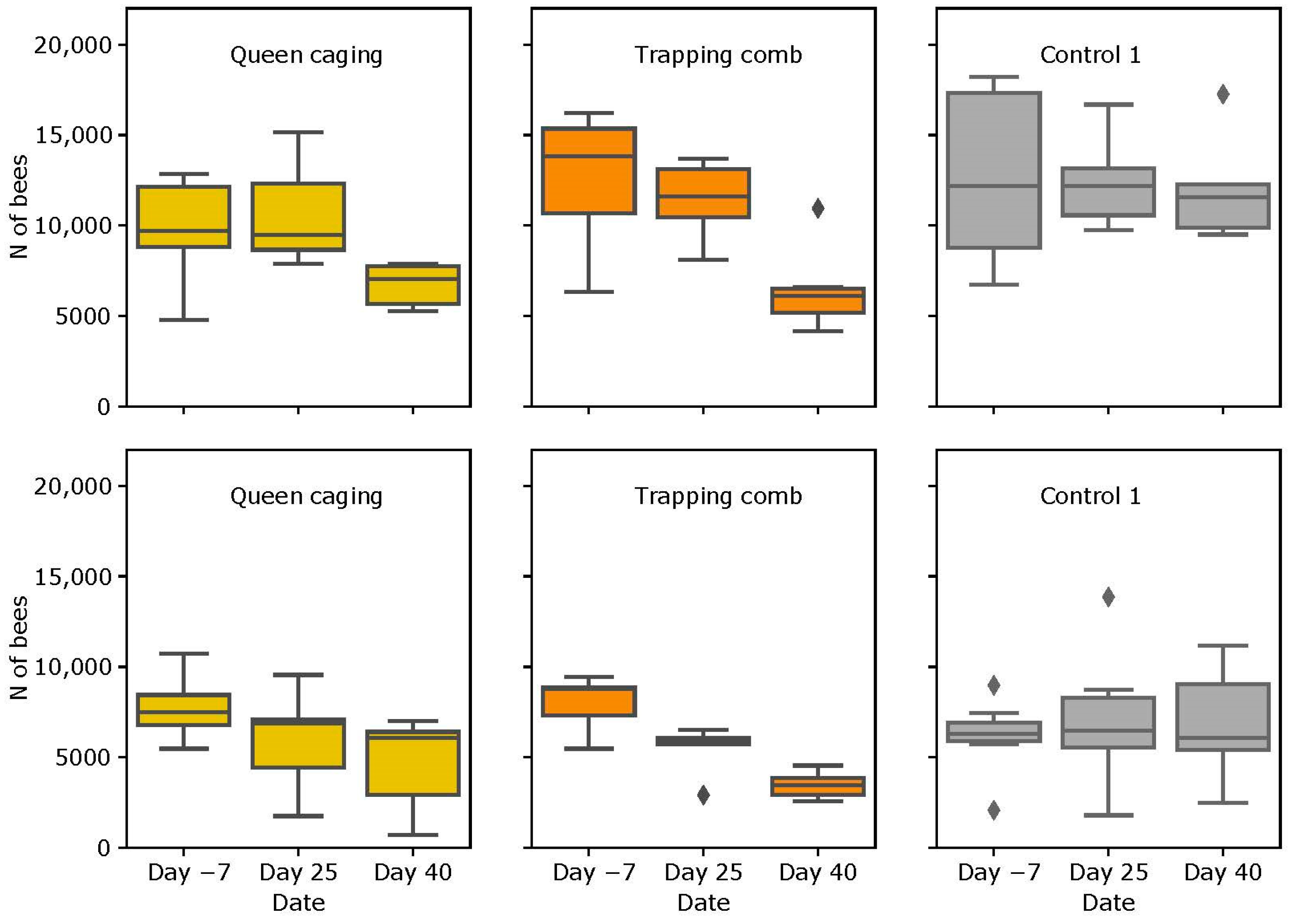
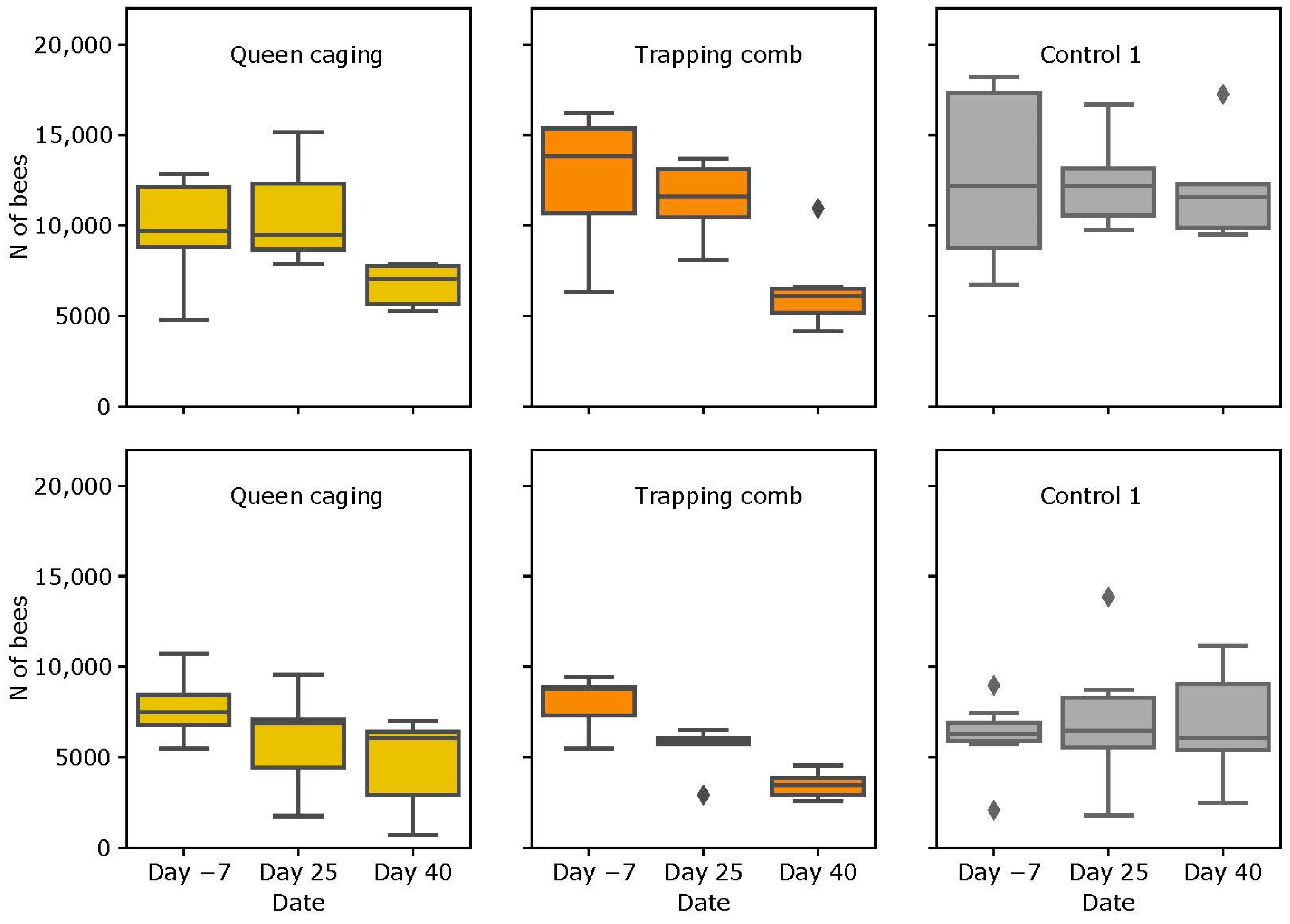
3.2. Colony Strength
3.2.1. Slovenian (SI) Apiary
3.2.2. Italian (IT) Apiary
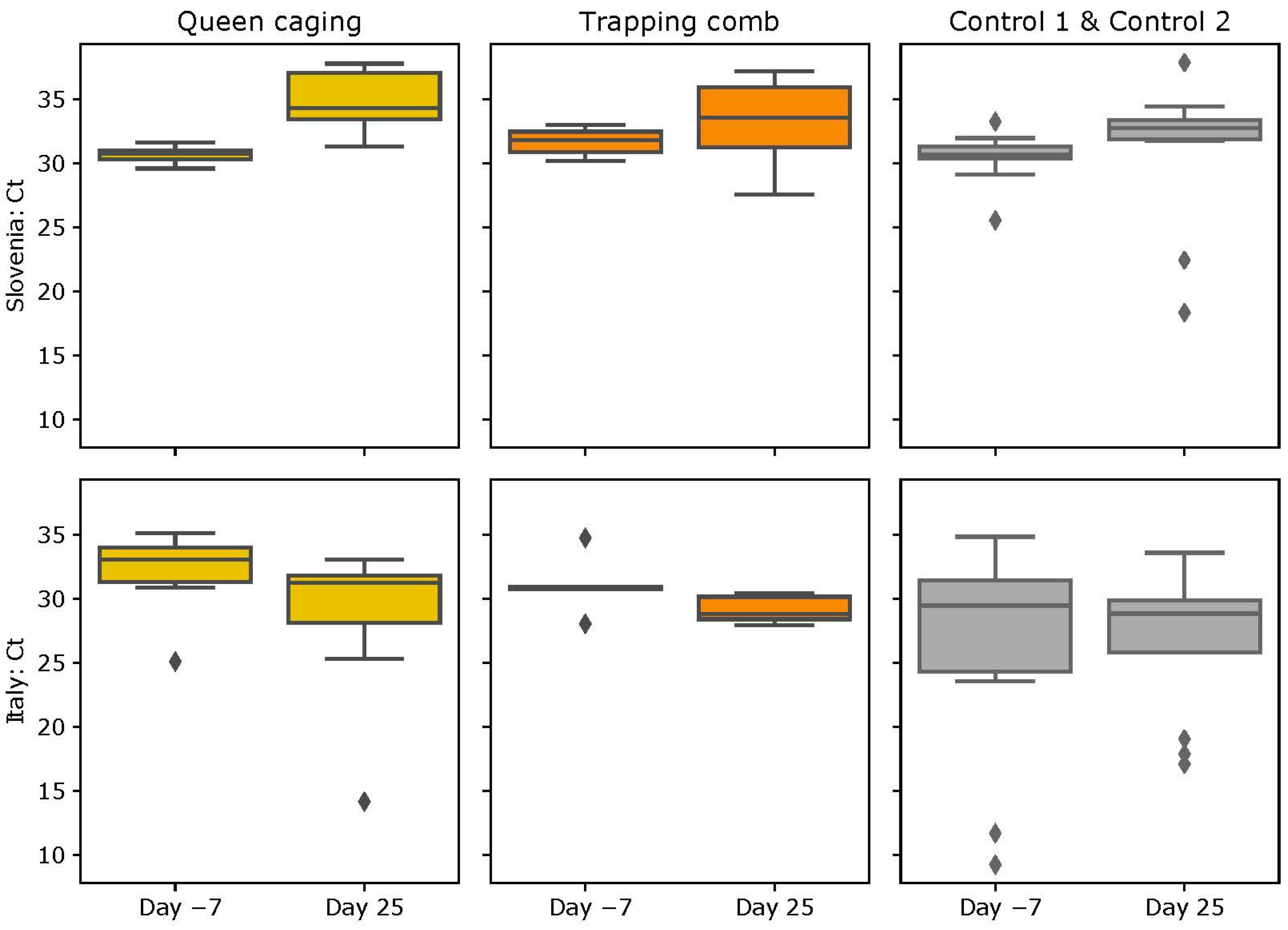
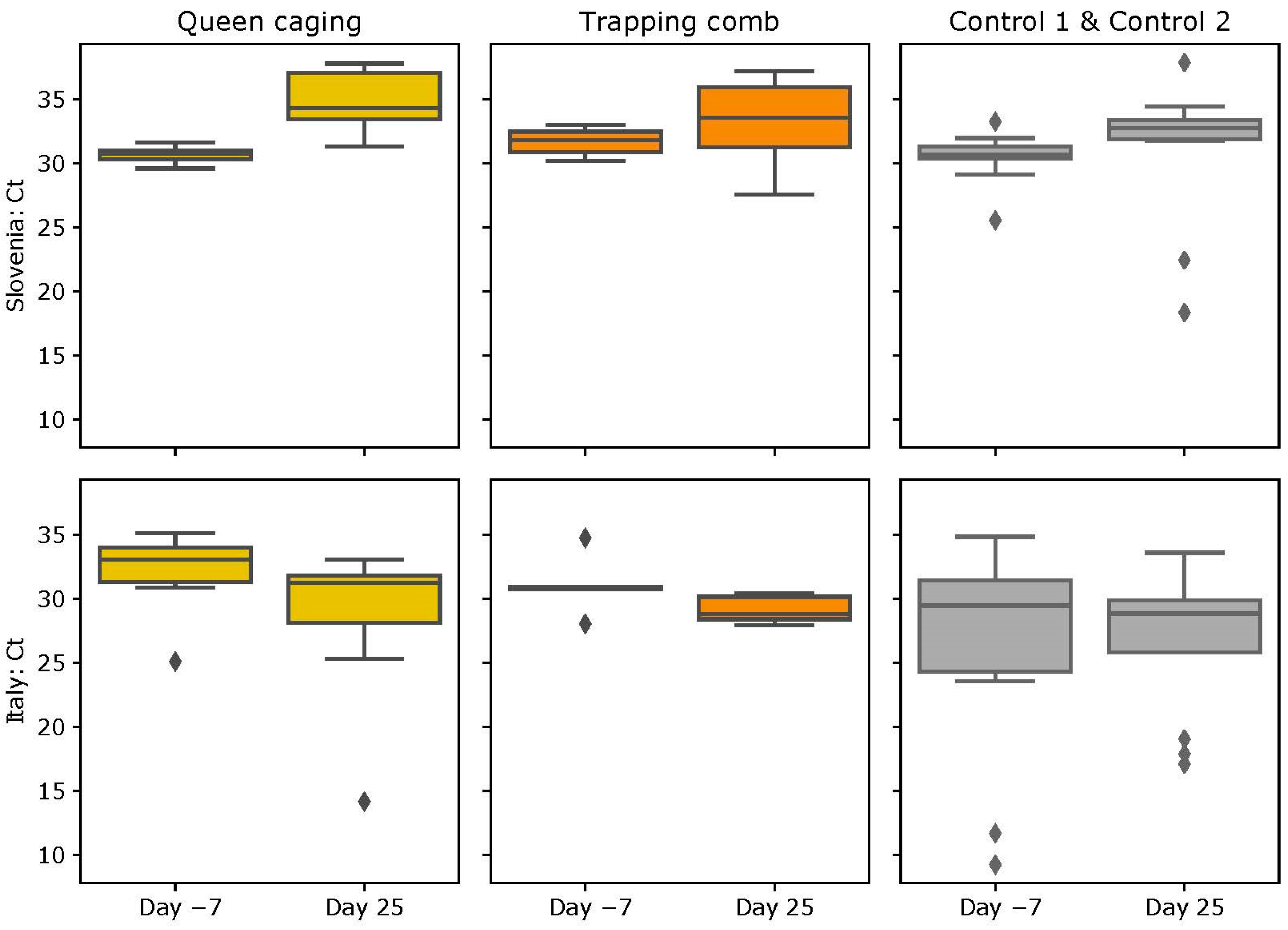
3.3. ABPV Loads
3.3.1. Slovenian (SI) Apiary
3.3.2. Italian (IT) Apiary
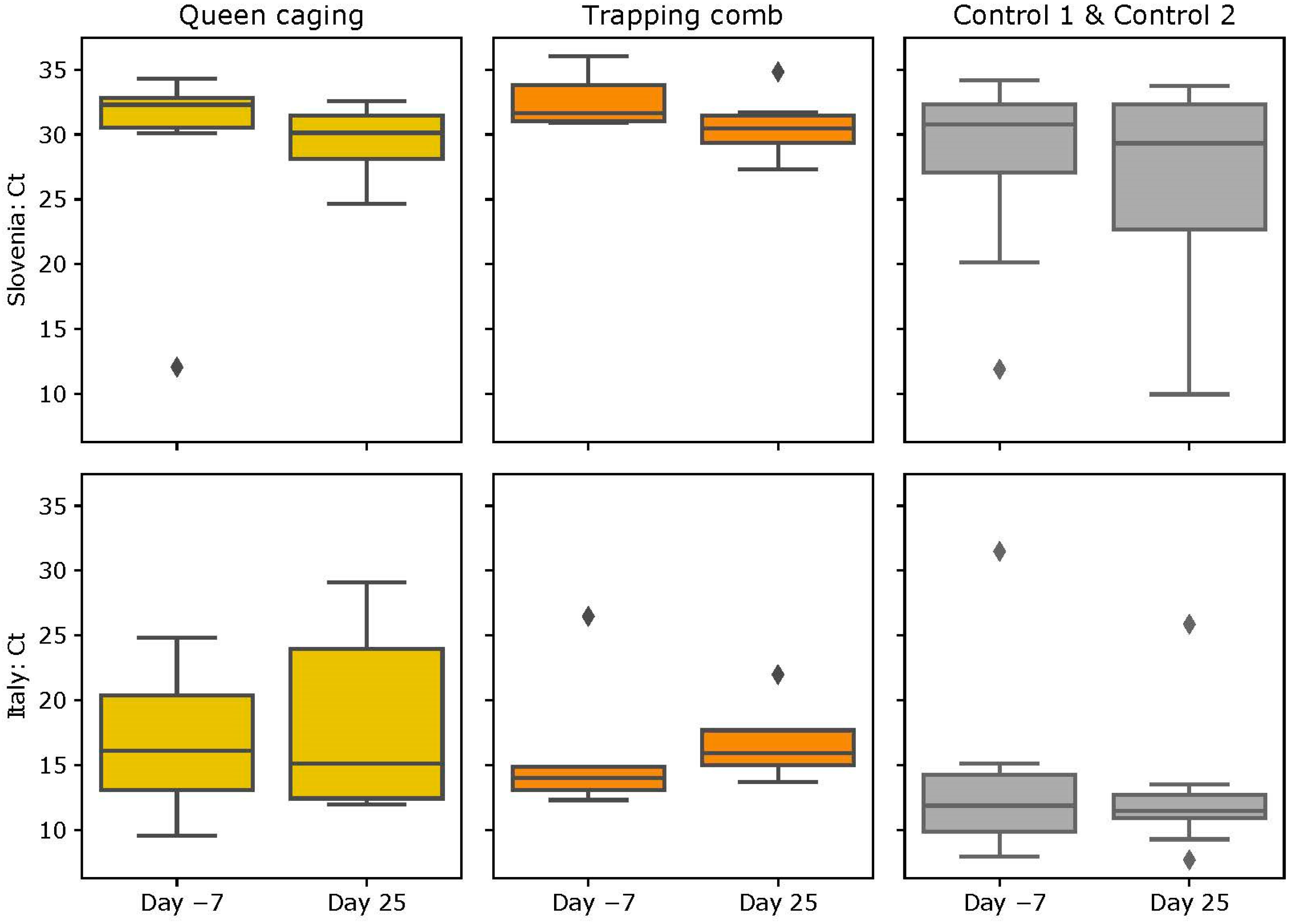
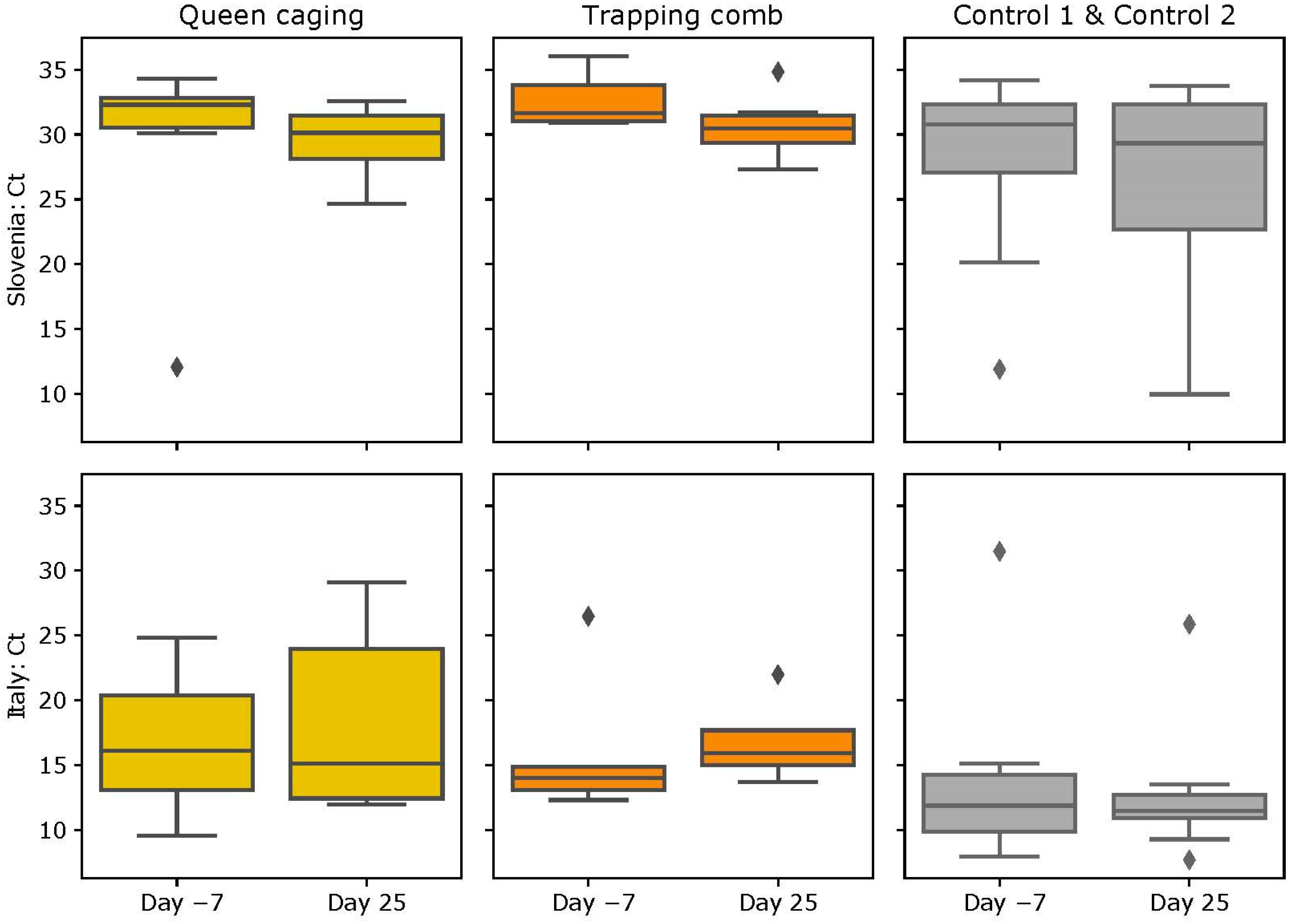
3.4. DWV Loads
3.4.1. Slovenian (SI) Apiary
3.4.2. Italian (IT) Apiary
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Bees | Brood Area (cm2) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Count | Mean | Std | Min | Max | Count | Mean | Std | Min | Max | ||
| Strength evaluation 1 | SI apiary | CG1 | 5 | 12,646 | 5074 | 6726 | 18,202 | 5 | 9384 | 3096 | 4845 | 13,430 |
| CG2 | 5 | 10,914 | 4254 | 4256 | 16,074 | 5 | 7650 | 6229 | 680 | 13,005 | ||
| QC | 6 | 9747 | 3006 | 4788 | 12,844 | 6 | 6134 | 4119 | 1360 | 11,985 | ||
| TC | 6 | 12,622 | 3820 | 6346 | 16,226 | 6 | 10,483 | 3853 | 7055 | 17,765 | ||
| IT apiary | CG1 | 8 | 6186 | 1965 | 2074 | 8971 | 8 | 15,677 | 7443 | 2074 | 26,581 | |
| CG2 | 5 | 7027 | 1925 | 5429 | 9553 | 5 | 11,973 | 2746 | 8660 | 16,190 | ||
| QC | 8 | 7739 | 1589 | 5476 | 10,718 | 8 | 15,060 | 3964 | 7907 | 19,202 | ||
| TC | 5 | 7969 | 1597 | 5476 | 9437 | 5 | 19,985 | 7743 | 9563 | 29,367 | ||
| Strength evaluation 2 | SI apiary | CG1 | 5 | 12,464 | 2712 | 9728 | 16,682 | 5 | 9758 | 2060 | 6715 | 12,325 |
| CG2 | 5 | 11,560 | 2614 | 9348 | 15,998 | 5 | 10,166 | 1627 | 8500 | 12,580 | ||
| QC | 6 | 10,602 | 2882 | 7866 | 15,162 | 6 | 0 | 0 | 0 | 0 | ||
| TC | 6 | 11,444 | 2120 | 8094 | 13,680 | 6 | 1530 | 3748 | 0 | 9180 | ||
| IT apiary | CG1 | 7 | 7103 | 3710 | 1794 | 13,864 | 7 | 19,739 | 9761 | 7756 | 39,608 | |
| CG2 | 5 | 4609 | 1185 | 3775 | 6408 | 5 | 21,912 | 2296 | 18,825 | 24,849 | ||
| QC | 7 | 5892 | 2849 | 1748 | 9553 | 0 | ||||||
| TC | 5 | 5429 | 1438 | 2913 | 6524 | 0 | ||||||
| Strength evaluation 3 | SI apiary | CG1 | 5 | 12,092 | 3105 | 9500 | 17,252 | 5 | 6052 | 2172 | 3655 | 8585 |
| CG2 | 0 | 0 | ||||||||||
| QC | 6 | 6739 | 1209 | 5282 | 7866 | 6 | 0 | 0 | 0 | 0 | ||
| TC | 6 | 6466 | 2376 | 4142 | 10,944 | 6 | 0 | 0 | 0 | 0 | ||
| IT apiary | CG1 | 7 | 6947 | 2985 | 2493 | 11,184 | 7 | 20,417 | 10,819 | 9864 | 41,415 | |
| CG2 | 0 | 0 | ||||||||||
| QC | 7 | 4630 | 2572 | 722 | 6990 | 0 | ||||||
| TC | 5 | 3467 | 782 | 2563 | 4544 | 0 | ||||||
| ABPV | DWV | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Count | Mean | Std | Min | Max | Count | Mean | Std | Min | Max | |||
| SI apiary | CG1 | before | 5 | 29.89 | 2.50 | 25.56 | 31.97 | 5 | 28.46 | 9.35 | 11.90 | 34.16 |
| after | 5 | 31.48 | 7.56 | 18.36 | 37.87 | 5 | 27.54 | 8.62 | 12.63 | 33.77 | ||
| CG2 | before | 5 | 31.02 | 1.51 | 29.15 | 33.26 | 5 | 27.77 | 4.80 | 20.16 | 32.42 | |
| after | 5 | 30.40 | 4.47 | 22.45 | 33.06 | 5 | 24.67 | 9.42 | 9.96 | 32.60 | ||
| QC | before | 6 | 30.66 | 0.70 | 29.60 | 31.61 | 6 | 28.98 | 8.40 | 12.06 | 34.28 | |
| after | 6 | 34.80 | 2.58 | 31.30 | 37.78 | 6 | 29.47 | 2.93 | 24.65 | 32.55 | ||
| TC | before | 6 | 31.68 | 1.10 | 30.18 | 32.99 | 6 | 32.59 | 2.15 | 30.89 | 36.03 | |
| after | 6 | 33.19 | 3.63 | 27.57 | 37.21 | 6 | 30.63 | 2.55 | 27.30 | 34.83 | ||
| IT apiary | CG3 | before | 7 | 24.21 | 10.03 | 9.25 | 34.84 | 8 | 13.93 | 7.47 | 7.96 | 31.49 |
| after | 7 | 26.77 | 6.56 | 17.11 | 33.59 | 7 | 13.99 | 5.32 | 10.99 | 25.87 | ||
| CG4 | before | 5 | 29.52 | 3.82 | 23.57 | 33.84 | 5 | 12.34 | 2.07 | 9.64 | 15.11 | |
| after | 5 | 26.88 | 4.39 | 19.06 | 29.30 | 5 | 10.39 | 1.97 | 7.69 | 12.72 | ||
| QC | before | 8 | 32.10 | 3.16 | 25.10 | 35.14 | 8 | 16.56 | 5.18 | 9.56 | 24.85 | |
| after | 8 | 28.50 | 6.28 | 14.17 | 33.06 | 8 | 18.19 | 7.01 | 11.95 | 29.09 | ||
| TC | before | 5 | 31.04 | 2.39 | 28.05 | 34.74 | 5 | 16.14 | 5.85 | 12.31 | 26.47 | |
| after | 5 | 29.14 | 1.09 | 27.95 | 30.43 | 5 | 16.85 | 3.22 | 13.67 | 21.98 | ||
References
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari: Varroidae) Is More than One Species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- La Rúa, P.D.; Jaffé, R.; Dall’Olio, R.; Muñoz, I.; Serrano, J. Biodiversity, Conservation and Current Threats to European Honeybees. Apidologie 2009, 40, 263–284. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor Feeds Primarily on Honey Bee Fat Body Tissue and Not Hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and Control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Nekoei, S.; Rezvan, M.; Khamesipour, F.; Mayack, C.; Molento, M.B.; Revainera, P.D. A Systematic Review of Honey Bee (Apis mellifera, Linnaeus, 1758) Infections and Available Treatment Options. Vet. Med. Sci. 2023, 9, 1848–1860. [Google Scholar] [CrossRef]
- Mondet, F.; de Miranda, J.R.; Kretzschmar, A.; Conte, Y.L.; Mercer, A.R. On the Front Line: Quantitative Virus Dynamics in Honeybee (Apis mellifera L.) Colonies along a New Expansion Front of the Parasite Varroa destructor. PLoS Pathog. 2014, 10, e1004323. [Google Scholar] [CrossRef]
- Noël, A.; Le Conte, Y.; Mondet, F. Varroa destructor: How Does It Harm Apis mellifera Honey Bees and What Can Be Done about It? Emerg. Top. Life Sci. 2020, 4, 45–57. [Google Scholar] [CrossRef]
- Boecking, O.; Genersch, E. Varroosis—The Ongoing Crisis in Bee Keeping. J. Für Verbraucherschutz Leb. 2008, 3, 221–228. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, B. Honey Bee Pathology; Acdemic Press: London, UK, 1991; Volume 1991. [Google Scholar]
- Ball, B.V. The Association of Varroa jacobsoni with Virus Diseases of Honey Bees. Wagenin-Gen 1983, 21–23. [Google Scholar] [CrossRef]
- Wiegers, F.P. Transmission of Honeybee Viruses by Varroa jacobsoni Oud. In European Research on Varroatosis Control; CRC Press: Boca Raton, FL, USA, 1988; ISBN 978-1-00-321146-4. [Google Scholar]
- Yue, C.; Genersch, E. RT-PCR Analysis of Deformed Wing Virus in Honeybees (Apis mellifera) and Mites (Varroa destructor). J. Gen. Virol. 2005, 86, 3419–3424. [Google Scholar] [CrossRef]
- de Miranda, J.R.; Cordoni, G.; Budge, G. The Acute Bee Paralysis Virus–Kashmir Bee Virus–Israeli Acute Paralysis Virus Complex. J. Invertebr. Pathol. 2010, 103, S30–S47. [Google Scholar] [CrossRef]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and Biological Characterization of Deformed Wing Virus of Honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef]
- Paxton, R.J.; Schäfer, M.O.; Nazzi, F.; Zanni, V.; Annoscia, D.; Marroni, F.; Bigot, D.; Laws-Quinn, E.R.; Panziera, D.; Jenkins, C.; et al. Epidemiology of a Major Honey Bee Pathogen, Deformed Wing Virus: Potential Worldwide Replacement of Genotype A by Genotype B. Int. J. Parasitol. Parasites Wildl. 2022, 18, 157–171. [Google Scholar] [CrossRef]
- Herrero, S.; Millán-Leiva, A.; Coll, S.; González-Martínez, R.M.; Parenti, S.; González-Cabrera, J. Identification of New Viral Variants Specific to the Honey Bee Mite Varroa destructor. Exp. Appl. Acarol. 2019, 79, 157–168. [Google Scholar] [CrossRef]
- Dainat, B.; Ken, T.; Berthoud, H.; Neumann, P. The Ectoparasitic Mite Tropilaelaps Mercedesae (Acari, Laelapidae) as a Vector of Honeybee Viruses. Insectes Sociaux 2009, 56, 40–43. [Google Scholar] [CrossRef]
- Eyer, M.; Chen, Y.P.; Schäfer, M.O.; Pettis, J.; Neumann, P. Small Hive Beetle, Aethina Tumida, as a Potential Biological Vector of Honeybee Viruses. Apidologie 2009, 40, 419–428. [Google Scholar] [CrossRef]
- Gisder, S.; Aumeier, P.; Genersch, E. Deformed Wing Virus: Replication and Viral Load in Mites (Varroa destructor). J. Gen. Virol. 2009, 90, 463–467. [Google Scholar] [CrossRef]
- Shen, M.; Yang, X.; Cox-Foster, D.; Cui, L. The Role of Varroa Mites in Infections of Kashmir Bee Virus (KBV) and Deformed Wing Virus (DWV) in Honey Bees. Virology 2005, 342, 141–149. [Google Scholar] [CrossRef]
- Bailey, L.; Gibbs, A.J.; Woods, R.D. Two Viruses from Adult Honey Bees (Apis mellifera Linnaeus). Virology 1963, 21, 390–395. [Google Scholar] [CrossRef]
- Genersch, E.; Aubert, M. Emerging and Re-Emerging Viruses of the Honey Bee (Apis mellifera L.). Vet. Res. 2010, 41, 54. [Google Scholar] [CrossRef]
- Bailey, L. Viruses Attacking the Honey Bee. In Advances in Virus Research; Lauffer, M.A., Bang, F.B., Maramorosch, K., Smith, K.M., Eds.; Academic Press: Cambridge, MA, USA, 1976; Volume 20, pp. 271–304. [Google Scholar]
- Yue, C.; Schröder, M.; Bienefeld, K.; Genersch, E. Detection of Viral Sequences in Semen of Honeybees (Apis mellifera): Evidence for Vertical Transmission of Viruses through Drones. J. Invertebr. Pathol. 2006, 92, 105–108. [Google Scholar] [CrossRef]
- Avalos, J.; Rosero, H.; Maldonado, G.; Reynaldi, F.J. Honey Bee Louse (Braula schmitzi) as a Honey Bee Virus Vector? J. Apic. Res. 2019, 58, 427–429. [Google Scholar] [CrossRef]
- Hunter, W.; Ellis, J.; vanEngelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-Scale Field Application of RNAi Technology Reducing Israeli Acute Paralysis Virus Disease in Honey Bees (Apis mellifera, Hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a Bee-Affecting Virus Associated with Colony Collapse Disorder Can Be Silenced by dsRNA Ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Yan, X.; Han, R. Prevention of Chinese Sacbrood Virus Infection in Apis cerana Using RNA Interference. Curr. Microbiol. 2010, 61, 422–428. [Google Scholar] [CrossRef]
- Locke, B.; Semberg, E.; Forsgren, E.; de Miranda, J.R. Persistence of Subclinical Deformed Wing Virus Infections in Honeybees Following Varroa Mite Removal and a Bee Population Turnover. PLoS ONE 2017, 12, e0180910. [Google Scholar] [CrossRef]
- Research and Extension Unit (AGDR). Thematic Catalogue for Smallholder Farmers to Promote Innovation—Main Bee Diseases: Good Beekeeping Practices; FAO: Rome, Italy, 2018; ISBN 978-92-5-130544-7. [Google Scholar]
- FAO NSP—Integrated Pest Management. Available online: https://www.fao.org/pest-and-pesticide-management/ipm/integrated-pest-management/en/ (accessed on 31 January 2024).
- Büchler, R.; Uzunov, A.; Kovačić, M.; Prešern, J.; Pietropaoli, M.; Hatjina, F.; Pavlov, B.; Charistos, L.; Formato, G.; Galarza, E.; et al. Summer Brood Interruption as Integrated Management Strategy for Effective Varroa Control in Europe. J. Apic. Res. 2020, 59, 764–773. [Google Scholar] [CrossRef]
- Giacomelli, A.; Pietropaoli, M.; Carvelli, A.; Iacoponi, F.; Formato, G. Combination of Thymol Treatment (Apiguard®) and Caging the Queen Technique to Fight Varroa destructor. Apidologie 2016, 47, 606–616. [Google Scholar] [CrossRef]
- Maul, V.; Klepsch, A.; Assmann-Werthmüller, U. Das Bannwabenverfahren Als Element Imkerlicher Betriebsweise Bei Starkem Befall Mit Varroa jacobsoni Oud. Apidologie 1988, 19, 139–154. [Google Scholar] [CrossRef]
- Calis, J.N.M.; Boot, W.J.; Beetsma, J.; van den Eijnde, J.H.P.M.; de Ruijter, A.; van der Steen, J.J.M. Effective Biotechnical Control of Varroa: Applying Knowledge on Brood Cell Invasion to Trap Honey Bee Parasites in Drone Brood. J. Apic. Res. 1999, 38, 49–61. [Google Scholar] [CrossRef]
- Gregorc, A.; Alburaki, M.; Werle, C.; Knight, P.R.; Adamczyk, J. Brood Removal or Queen Caging Combined with Oxalic Acid Treatment to Control Varroa Mites (Varroa destructor) in Honey Bee Colonies (Apis mellifera). Apidologie 2017, 48, 821–832. [Google Scholar] [CrossRef]
- Lodesani, M.; Franceschetti, S.; Dall’Ollio, R. Evaluation of Early Spring Bio-Technical Management Techniques to Control Varroosis in Apis mellifera. Apidologie 2019, 50, 131–140. [Google Scholar] [CrossRef]
- Branco, M.R.; Kidd, N.A.C.; Pickard, R.S. A Comparative Evaluation of Sampling Methods for Varroa destructor (Acari: Varroidae) Population Estimation. Apidologie 2006, 37, 452–461. [Google Scholar] [CrossRef]
- Delaplane, K.S.; van der Steen, J.; Guzman-Novoa, E. Standard Methods for Estimating Strength Parameters of Apis mellifera Colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard Methods for Varroa Research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef]
- Gabel, M.; Scheiner, R.; Büchler, R. Immediate and Long-Term Effects of Induced Brood Interruptions on the Reproductive Success of Varroa destructor. Apidologie 2023, 54, 20. [Google Scholar] [CrossRef]
- Liebig, G. Zur Behandlung von Bienenvölkern Mit Oxalsäure Und “Bienenwohl”. Dtsch. Bienen-J. 1999, 1999, 4–5. [Google Scholar]
- Formato, G.; Giacomelli, A.; Scaramozzino, P.; Comini, A.; Aquilini, E.; Ermenegildi, A.; Gallerini, D.; Muscolini, C.; Sirio, A.D.; Bragagnolo, A.; et al. Verifica dell’attività acaricida di acido ossalico sublimato in primavera con diverse condizioni di impiego. Lapis 2010, 7, 7–14. [Google Scholar]
- Gregorc, A.; Planinc, I. The Control of Varroa destructor Using Oxalic Acid. Vet. J. 2002, 163, 306–310. [Google Scholar] [CrossRef]
- Mutinelli, F.; Baggio, A.; Capolongo, F.; Piro, R.; Prandin, L.; Biasion, L. A Scientific Note on Oxalic Acid by Topical Application for the Control of Varroosis. Apidologie 1997, 28, 461–462. [Google Scholar] [CrossRef]
- Currie, R.W.; Gatien, P. Timing Acaricide Treatments to Prevent Varroa destructor (Acari: Varroidae) from Causing Economic Damage to Honey Bee Colonies. Can. Entomol. 2006, 138, 238–252. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Hood, W.M. Effects of Delayed Acaricide Treatment in Honey Bee Colonies Parasitized by Varroa jacobsoni and a Late-Season Treatment Threshold for the South-Eastern USA. J. Apic. Res. 1997, 36, 125–132. [Google Scholar] [CrossRef]
- Fluri, P.; Wille, H.; Gerig, L.; Lüscher, M. Juvenile Hormone, Vitellogenin and Haemocyte Composition in Winter Worker Honeybees (Apis mellifera). Experientia 1977, 33, 1240–1241. [Google Scholar] [CrossRef]
- Evans, K.C.; Underwood, R.M.; López-Uribe, M.M. Combined Effects of Oxalic Acid Sublimation and Brood Breaks on Varroa Mite (Varroa destructor) and Deformed Wing Virus Levels in Newly Established Honey Bee (Apis mellifera) Colonies. J. Apic. Res. 2022, 61, 197–205. [Google Scholar] [CrossRef]
- Anderson, K.E.; Copeland, D.C.; Erickson, R.J.; Floyd, A.S.; Maes, P.C.; Mott, B.M. A high-throughput sequencing survey characterizing European foulbrood disease and Varroosis in honey bees. Sci. Rep. 2023, 13, 1162. [Google Scholar] [CrossRef]
- Jack, C.J.; van Santen, E.; Ellis, J.D. Evaluating the Efficacy of Oxalic Acid Vaporization and Brood Interruption in Controlling the Honey Bee Pest Varroa destructor (Acari: Varroidae). J. Econ. Entomol. 2020, 113, 582–588. [Google Scholar] [CrossRef]



| −14 | −7 | 0 | 20 | 25 | 40 | 55 | 60 | 65 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| GROUPS | QC | Start | CS | C, SA | CS, OA, SA | CT | CE | CTE | End | |
| TC | Start | CS | C, SA | TCR | CS, OA, SA | CT | CE | CTE | End | |
| CG1 | Start | CS | SA | CS, OA, SA | End | |||||
| CG2 | Start | CS | SA | SA | CT, C | CE | CTE, End |
| Primer | Sequence |
|---|---|
| DWV (Forward) | 5′-ATGGGTTTGATTCRATATCTTGGAA-3′ |
| DWV (Reverse) | 5′-GATGTTCCRGGTGGCTTTAATGA-3′ |
| DWV Probe | 5′-FAM-ACTAGTGCTGGTTTTCCTTTGTC-NFQ-MGB. |
| ABPV (Forward) | 5′-GCCCAGACAAGCGCAGTACT-3′ |
| ABPV (Reverse) | 5′-AGCACGGAAAACGCGTCTT-3′ |
| ABPV Probe | 5′-FAM-TCCCCGATAGCRACCGA-MGBNFQ-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bubnič, J.; Prešern, J.; Pietropaoli, M.; Cersini, A.; Moškrič, A.; Formato, G.; Manara, V.; Smodiš Škerl, M.I. Integrated Pest Management Strategies to Control Varroa Mites and Their Effect on Viral Loads in Honey Bee Colonies. Insects 2024, 15, 115. https://doi.org/10.3390/insects15020115
Bubnič J, Prešern J, Pietropaoli M, Cersini A, Moškrič A, Formato G, Manara V, Smodiš Škerl MI. Integrated Pest Management Strategies to Control Varroa Mites and Their Effect on Viral Loads in Honey Bee Colonies. Insects. 2024; 15(2):115. https://doi.org/10.3390/insects15020115
Chicago/Turabian StyleBubnič, Jernej, Janez Prešern, Marco Pietropaoli, Antonella Cersini, Ajda Moškrič, Giovanni Formato, Veronica Manara, and Maja Ivana Smodiš Škerl. 2024. "Integrated Pest Management Strategies to Control Varroa Mites and Their Effect on Viral Loads in Honey Bee Colonies" Insects 15, no. 2: 115. https://doi.org/10.3390/insects15020115
APA StyleBubnič, J., Prešern, J., Pietropaoli, M., Cersini, A., Moškrič, A., Formato, G., Manara, V., & Smodiš Škerl, M. I. (2024). Integrated Pest Management Strategies to Control Varroa Mites and Their Effect on Viral Loads in Honey Bee Colonies. Insects, 15(2), 115. https://doi.org/10.3390/insects15020115