
Identification on Key Volatiles Contributed to Oviposition Preference of Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae) from High and Normal Oleic Varieties of Peanut

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Peanuts
2.2. Insect Rearing
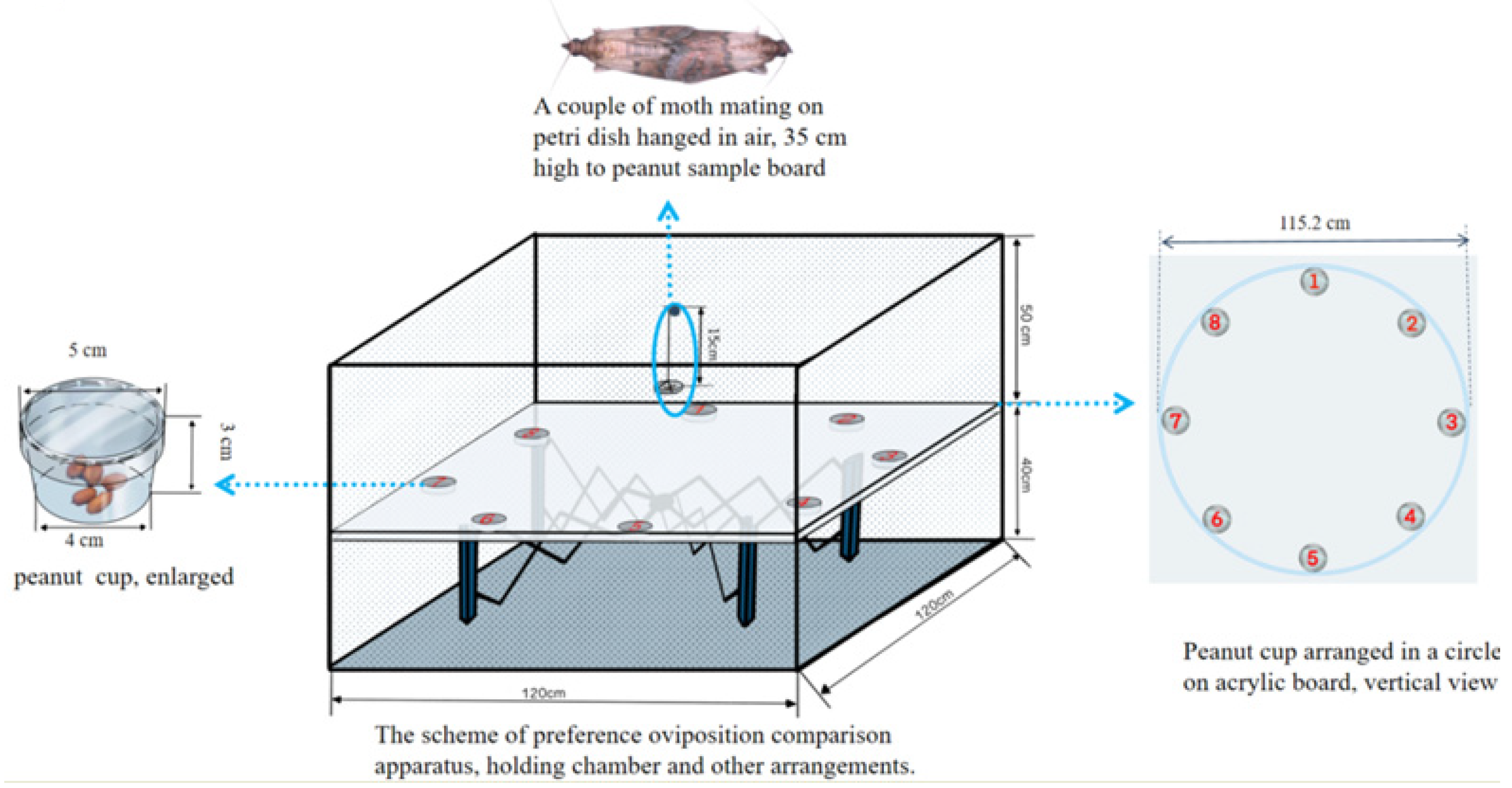
2.3. Oviposition Preference Measurement
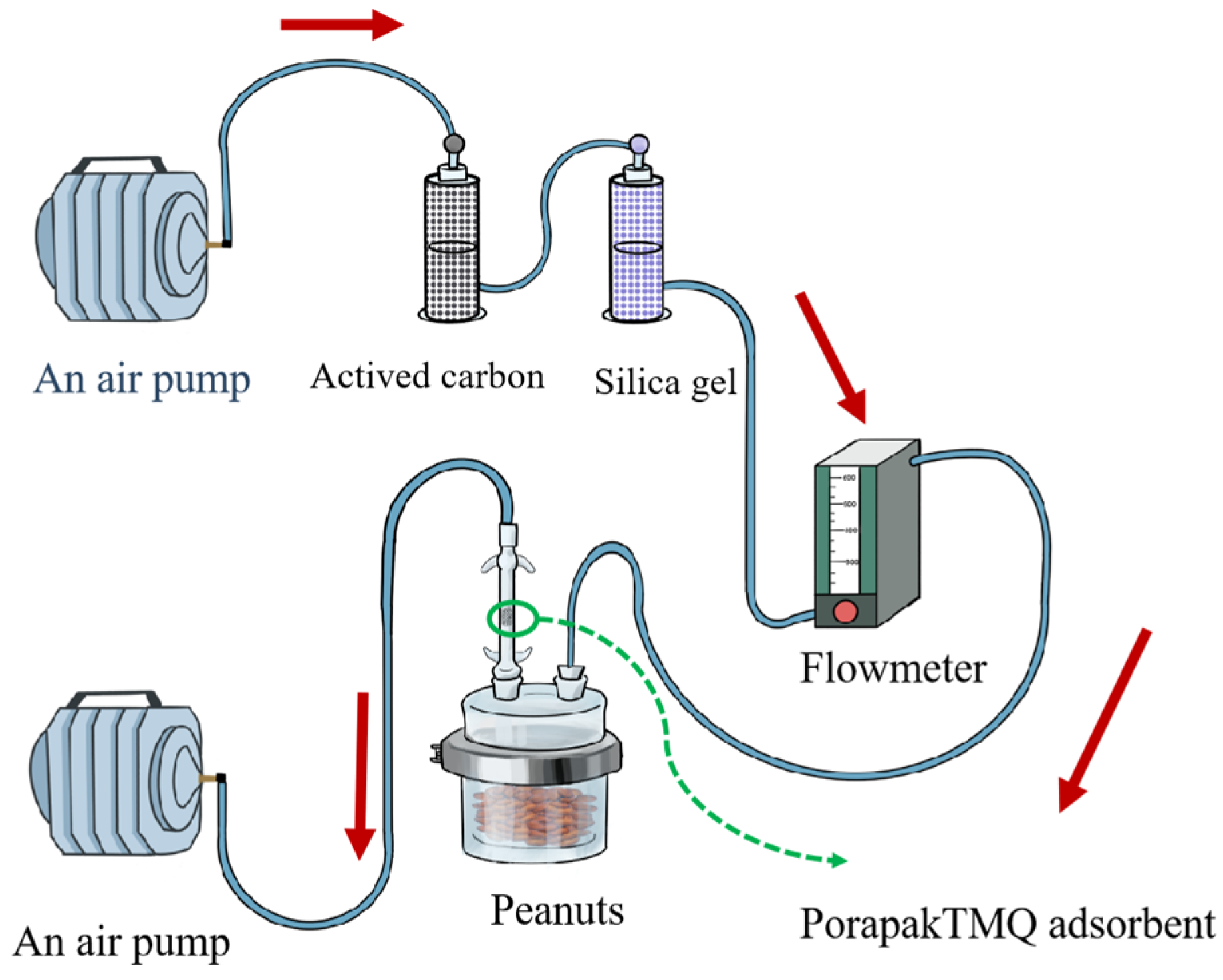
2.4. Volatile Organic Compounds Collection and Chemical Analysis
2.5. Chemicals
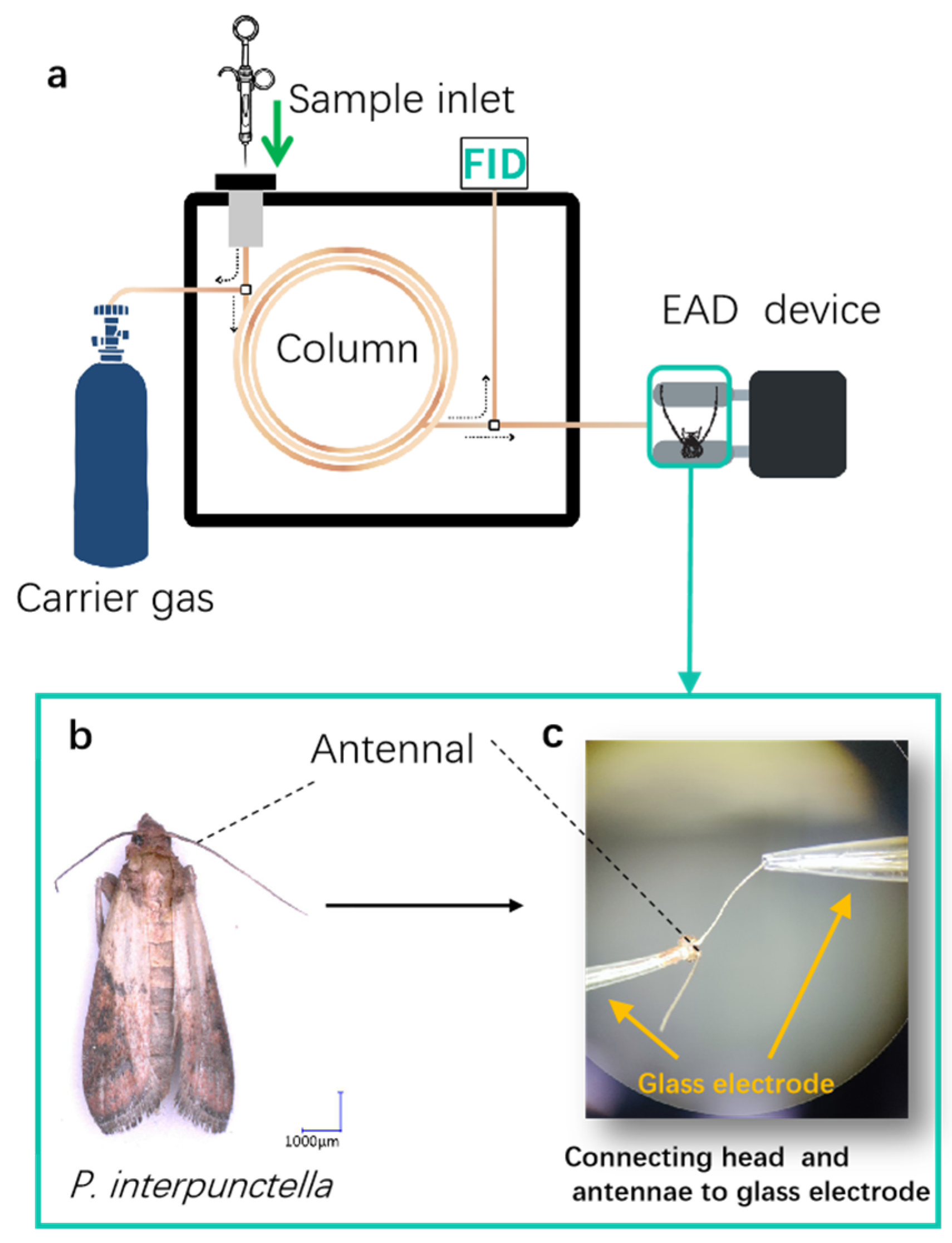
2.6. Electrophysiological Assays
2.7. Y-Tube Olfactometer Bioassay
2.8. Wind Tunnel Bioassay
2.9. Data Analysis
3. Results
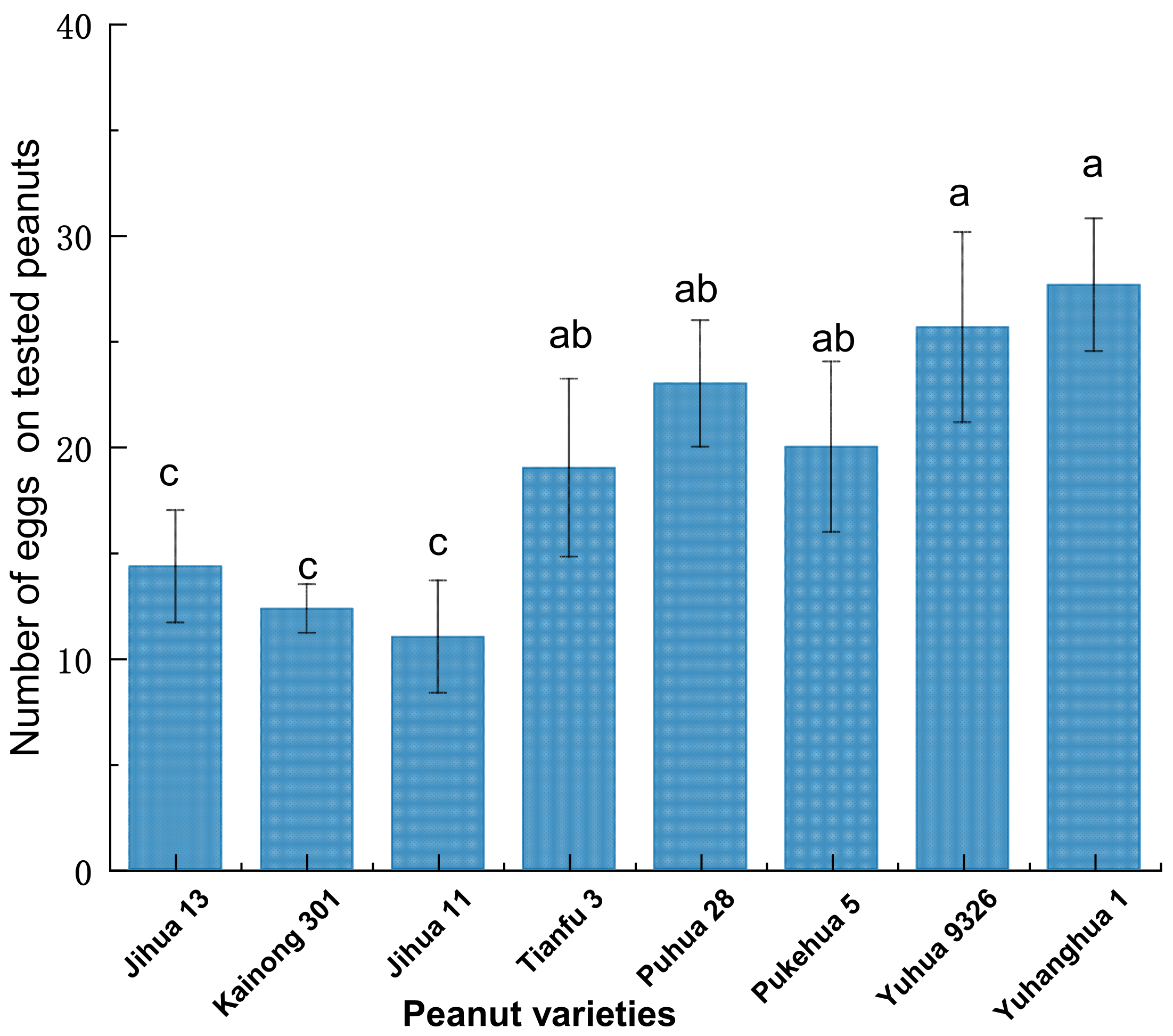
3.1. The Oviposition Preference of Plodia interpunctella Females
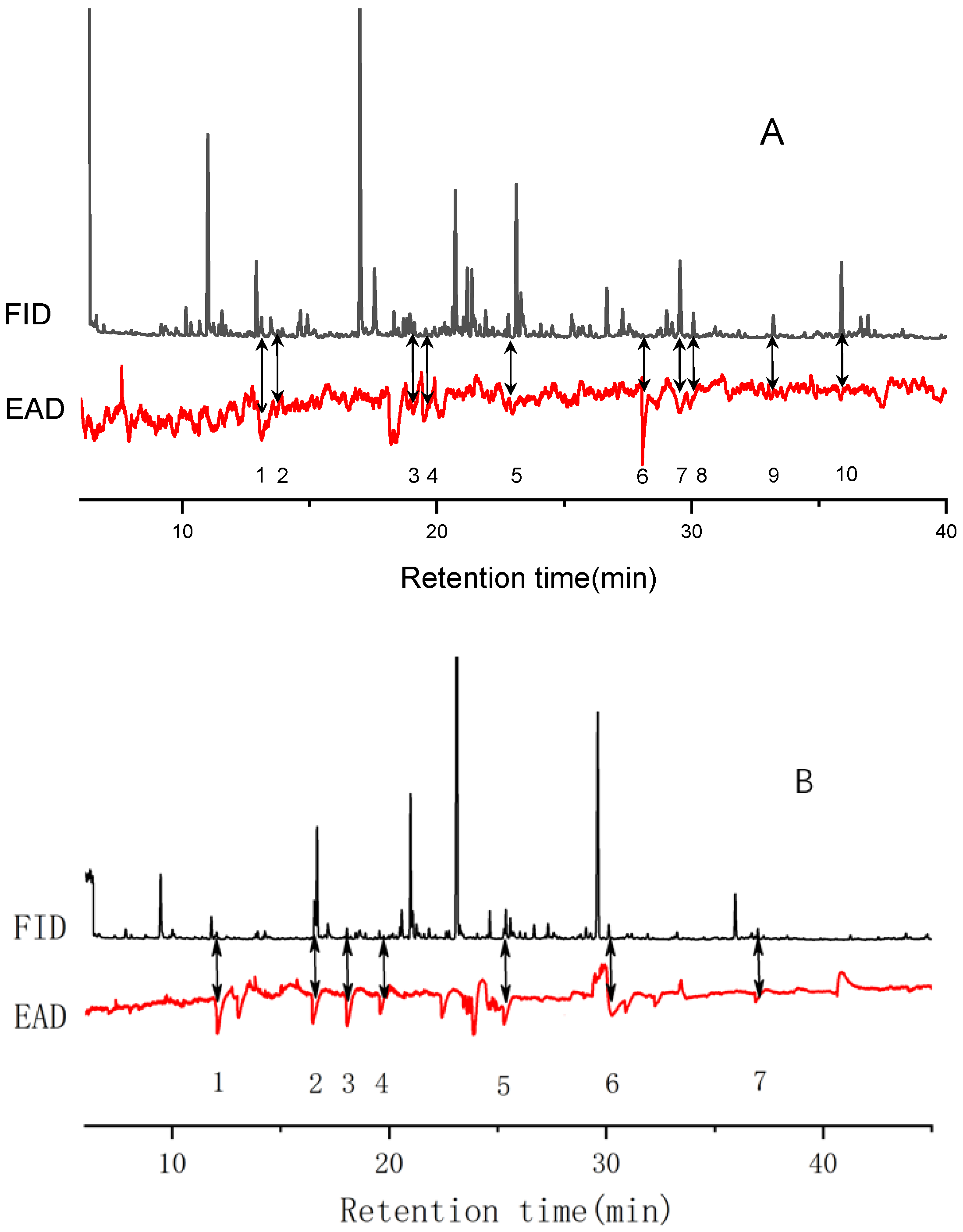
3.2. Chemical Analysis
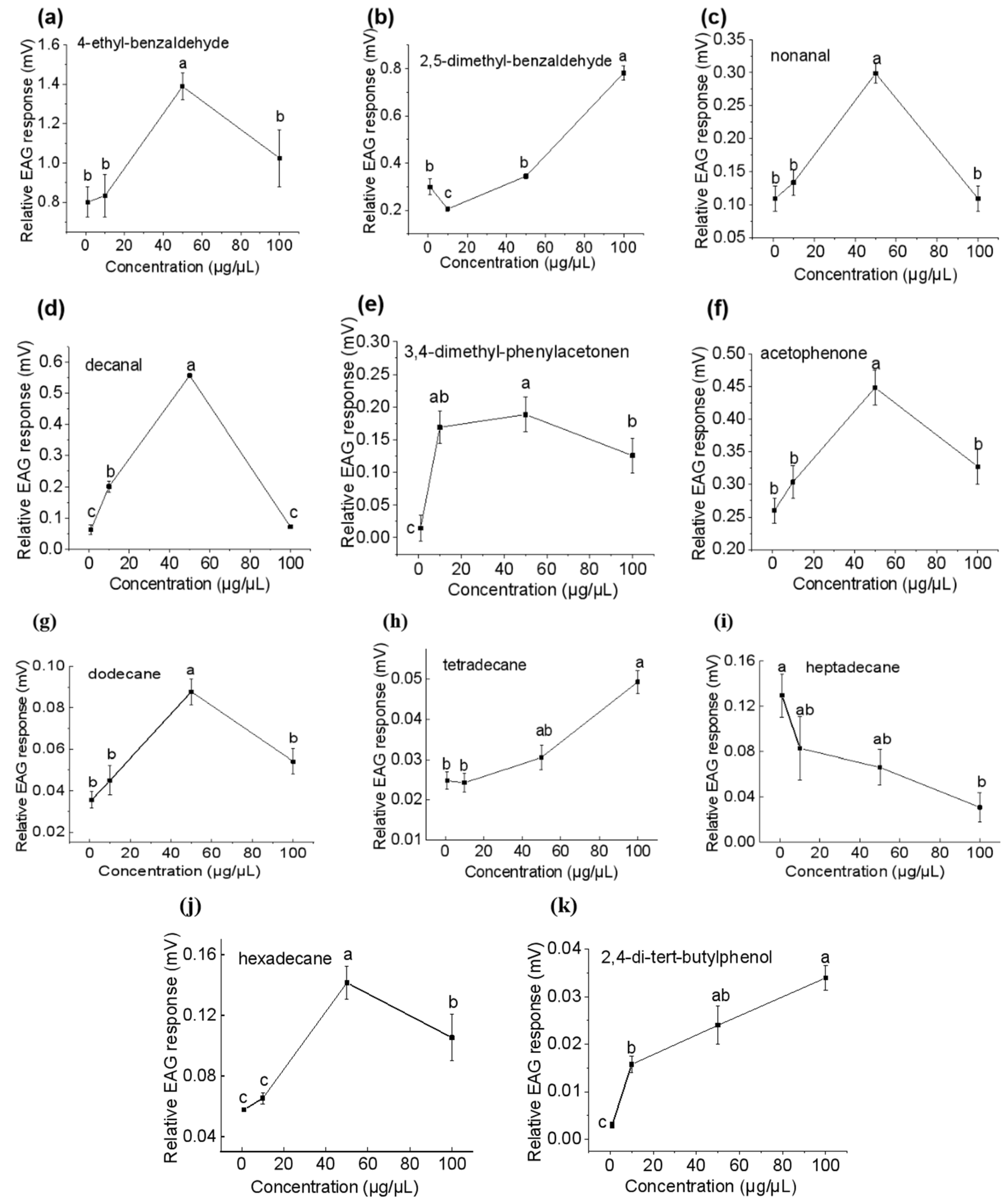
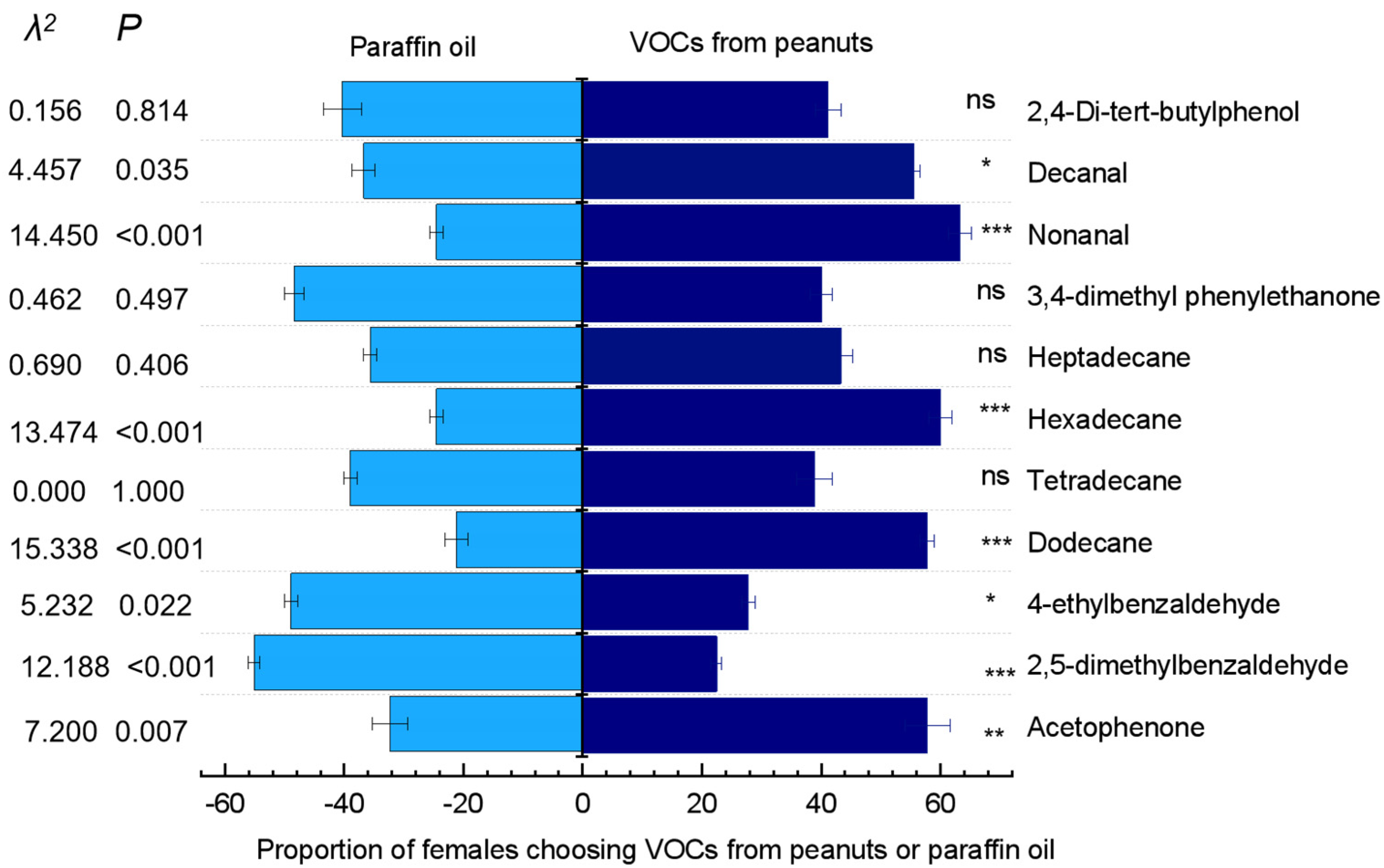
3.3. Antennal Responses of the Females to Volatile Organic Compounds
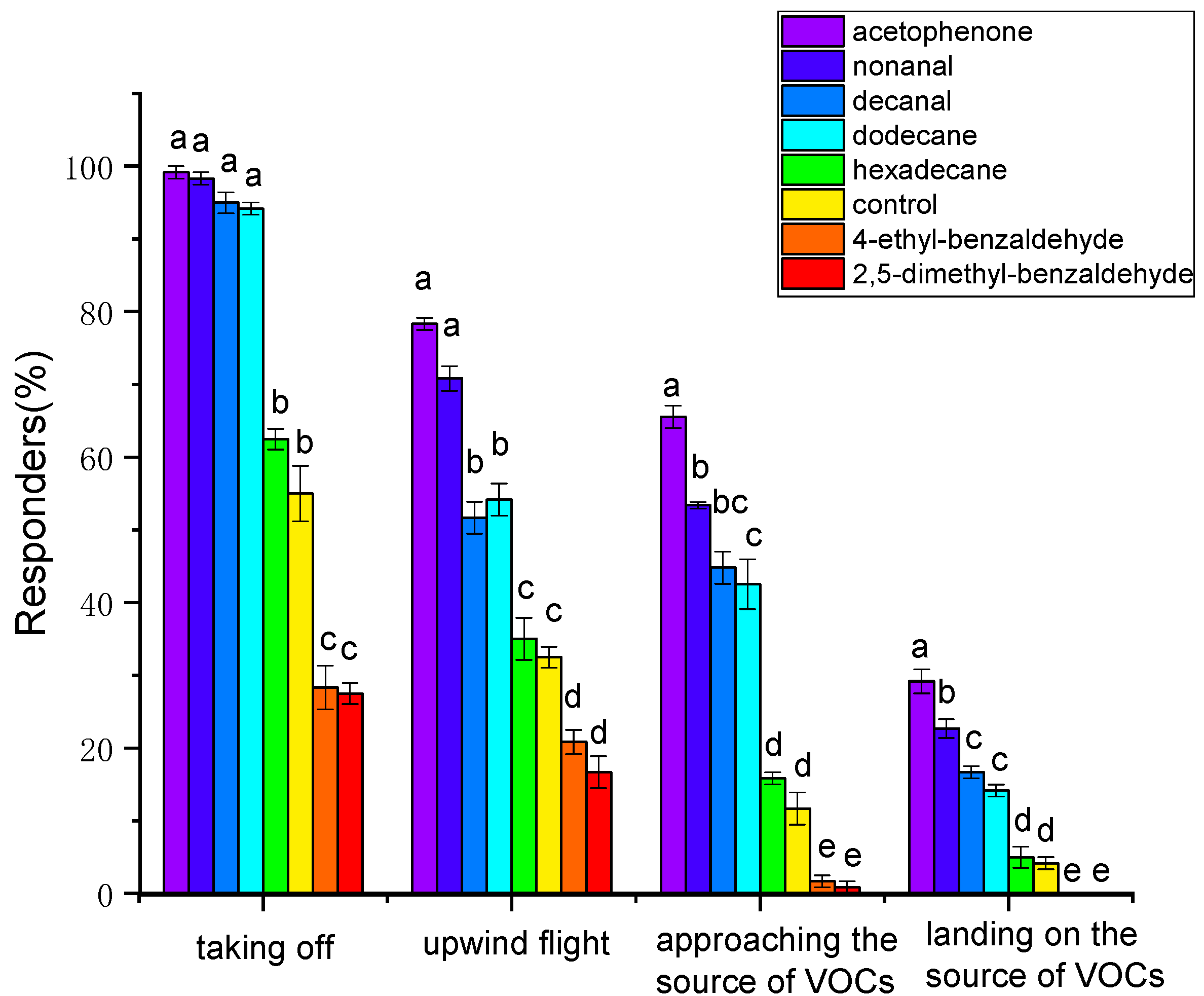
3.4. Behavioral Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wadood, S.A.; Nie, J.; Li, C.L.; Rogers, K.M.; Zhang, Y.Z.; Yuan, Y.W. Geographical origin classification of peanuts and processed fractions using stable isotopes. Food Chem.-X 2022, 16, 104456–104463. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.N.; Shi, A.M.; Liu, H.Z.; Yu, H.W.; Li, L.; Lin, W.J.; Qiang, W. Relationship of chemical properties of different peanut varieties to peanut butter storage stability. J. Integr. Agric. 2018, 17, 1003–1010. [Google Scholar] [CrossRef]
- Jastrombek, J.M.; Faguerazzi, M.M.; de Cássio Pierezan, H.; Rufato, L.; Sato, A.J.; da Silva Ricce, W.; Marques, V.V.; Leles, N.R.; Roberto, S.R. Hop: An emerging crop in subtropical areas in Brazil. Horticulturae 2022, 8, 393. [Google Scholar] [CrossRef]
- Qiang, W.C.; Zhang, J.T.; Yan, Y.S.; Wang, S. Current Situation and Future Directions of High Oleic Peanut Breeding in China. Shandong Agric. Sci. 2018, 50, 171–176. [Google Scholar]
- Ranga Rao, G.; Rameshwar Rao, V.; Nigam, S. Post-Harvest Insect Pests of Groundnut and Their Management; Information Bulletin No. 84; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2010; 20p, ISBN 978-92-9066-528-1. [Google Scholar]
- Arthur, F.H. Effects of cleaning peanuts on insect damage, insect population growth and insecticide efficacy. Peanut Sci. 2010, 16, 100–105. [Google Scholar] [CrossRef]
- Nesci, A.; Montemarani, A.; Etcheverry, M. Assessment of mycoflora and infestation of insects, vector of Aspergillus section Flavi, in stored peanut from Argentina. Mycotoxin Res. 2011, 27, 5–12. [Google Scholar] [CrossRef]
- Mohandass, S.; Arthur, F.H.; Zhu, K.Y.; Throne, J.E. Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products. J. Stored Prod. Res. 2007, 43, 302–311. [Google Scholar] [CrossRef]
- Yaman, M.; Saǧlam, T.; Ertürk, Ö. First Record, Distribution and Occurrence of A Protistan Entomopathogen, Adelina mesnili Perez (Coccidia: Adeleidae) in the Indian Meal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) Populations in Türkiye. Türkiye Parazitolojii Derg. 2023, 47, 151–155. [Google Scholar] [CrossRef]
- Mankin, R.W.; Hagstrum, D.W.; Nansen, C.; Meikle, W.G. Almond moth oviposition patterns in continuous layers of peanuts. J. Stored Prod. Res. 2014, 59, 48–54. [Google Scholar] [CrossRef]
- Warsi, S.; Mbata, G.N. Impact of Peanut Depth and Container Size on the Parasitism of Diapausing and Nondiapausing Larvae of Indian Meal Moth (Lepidoptera: Pyralidae) by Habrobracon hebetor (Hymenoptera: Braconidae). Environ. Entomol. 2018, 47, 1226–1232. [Google Scholar] [CrossRef]
- Pearson, T.; Brabec, D.L. Detection of wheat kernels with hidden insect infestations with an electrically conductive roller mill. Appl. Eng. Agric. 2007, 23, 639–645. [Google Scholar] [CrossRef]
- Perez, J.; Moore, R.; Abney, M.; Toews, M. Species Composition, Temporal Abundance and Distribution of Insect Captures Inside and Outside Commercial Peanut Shelling Facilities. Insects 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- ISO/TC34/SC4; Stored Cereal Grains and Pulses—Guidance on the Detection of Infestation by Live Invertebrates by Trapping. ISO: Geneva, Switzerland, 2004.
- Michereff, M.F.; Borges, M.; Aquino, M.F.; Laumann, R.A.; Mendes Gomes, A.C.; Blassioli-Moraes, M.C. The influence of volatile semiochemicals from stink bug eggs and oviposition-damaged plants on the foraging behaviour of the egg parasitoid Telenomus podisi. Bull. Entomol. Res. 2016, 106, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Li, X.; Yan, X.; Fan, W.; Hao, C. Electroantennogram responses of Plutella xylostella (L.) to sex pheromone components and host plant volatile semiochemicals. J. Appl. Entomol. 2020, 144, 396–406. [Google Scholar] [CrossRef]
- Zhang, Z.Z. Odorant-binding proteins and chemosensory proteins potentially involved in host plant recognition in the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2020, 76, 2609–2618. [Google Scholar] [CrossRef]
- Magalhães, D.M.; Borges, M.; Laumann, R.A.; Blassioli Moraes, M.C. Influence of multiple- and single-species infestations on herbivore-induced cotton volatiles and Anthonomus grandis behaviour. J. Pest Sci. 2018, 91, 1019–1032. [Google Scholar] [CrossRef]
- Zhao, J.Y.; Zheng, R.R.; Li, X.F.; Lyu, Z.S.; Ma, L.; Song, C.F.; Qie, X.T.; Yan, X.Z.; Hao, C. Electrophysiological and Behavioral Responses of Plodia interpunctella (Hübner) Females to Aldehyde Volatiles from Dried Fruits. J. Agric. Food Chem. 2023, 71, 17253–17262. [Google Scholar] [CrossRef]
- Ndomo-Moualeu, A.F.; Ulrichs, C.; Adler, C. Behavioral responses of Callosobruchus maculatus to volatile organic compounds found in the headspace of dried green pea seeds. J. Pest Sci. 2016, 89, 107–116. [Google Scholar] [CrossRef]
- Uechi, K.; Matsuyama, S.; Suzuki, T. Oviposition attractants for Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in the volatiles of whole wheat flour. J. Stored Prod. Res. 2007, 43, 193–201. [Google Scholar] [CrossRef]
- Būda, V.; Apšegaitė, V.; Blažytė-Čereškienė, L.; Butkienė, R.; Nedveckytė, I.; Pečiulytė, D. Response of moth Plodia interpunctella to volatiles of fungus-infected and uninfected wheat grain. J. Stored Prod. Res. 2016, 69, 152–158. [Google Scholar] [CrossRef]
- Jiang, B.R.; Wang, D.X.; Zhang, L.K.; Chen, L.; Jing, J.G.; Li, Z.H.; Tang, P.A. Comparison on Oviposition Preference of Plodia interpunctella (Hübner) on Grain and Peanut Kernels. J. Henan Univ. Technol. 2019, 40, 86–93. [Google Scholar]
- Astuti, L.P.; Rizali, A.; Tanzilia, S. Seed coat and variety of peanut inhibit host preference and development of Oryzaephilus mercator. J. Stored Prod. Res. 2018, 78, 98–104. [Google Scholar] [CrossRef]
- Huang, Y.L.; Wang, D.X.; Jian, F.J. Survival of Plodia interpunctella (Hübner) larvae treated with 98% N2 and the life history of their next generation. Bull. Entomol. Res. 2023, 113, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Zhao, N.N.; Liu, Q.Z.; Liu, Z.L.; Du, S.S.; Zhou, L.G.; Deng, Z.W. Repellent Constituents of Essential Oil of Cymbopogon distans Aerial Parts against Two Stored-Product Insects. J. Agric. Food Chem. 2011, 59, 9910–9915. [Google Scholar] [CrossRef] [PubMed]
- Dev, P.; Kennedy, J.; Srinivasan, T.; Arthanari, P.M. Dose Optimization in a Wind Tunnel to Determine the Effective Concentration preferred by Male Fall Armyworm Moths. Biol. Forum 2022, 14, 96–100. [Google Scholar]
- Cao, Y.; Li, S.; Benelli, G.; Germinara, G.S.; Yang, J.; Yang, W.J.; Li, C. Olfactory responses of stegobium paniceum to different chinese medicinal plant materials and component analysis of volatiles. J. Stored Prod. Res. 2018, 76, 122–128. [Google Scholar] [CrossRef]
- Trematerra, P.; Sciarreta, A.; Tamasi, E. Behavioural responses of Oryzaephilus surinamensis, Tribolium castaneum and Tribolium confusum to naturally and artificially damaged durum wheat kernels. Entomol. Exp. Appl. 2000, 94, 195–200. [Google Scholar] [CrossRef]
- Cao, Y.; Jian, L.; Athanassiou, C.G.; Yang, Y.; Hu, Q.; Zhang, X.Y.; Dai, F.L.; Maggi, F. Behavioral responses of Rhyzopertha dominica (F.) to volatiles of different stored grains. J. Stored Prod. Res. 2024, 105, 102235–102242. [Google Scholar] [CrossRef]
- Ukeh, D.A.; Birkett, M.A.; Bruce, T.J.; Allan, E.J.; Pickett, J.A.; Luntz, A.J. Behavioural responses of the maize weevil, Sitophilus zeamais, to host (stored-grain) and non-host plant volatiles. Pest Manag. Sci. 2010, 66, 44–50. [Google Scholar] [CrossRef]
- Tsuchida, T. Molecular basis and ecological relevance of aphid body colors. Curr. Opin. Insect Sci. 2016, 17, 74–80. [Google Scholar] [CrossRef]
- Germinara, G.S.; De Cristofaro, A.; Rotundo, G. Behavioral responses of adult Sitophilus granarius to individual cereal volatiles. J. Chem. Ecol. 2008, 34, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Olsson, P.O.; Anderbrant, O.; Lofstedt, C.; Borg-Karlson, A.K.; Liblikas, I. Electrophysiological and behavioral responses to chocolate volatiles in both sexes of the pyralid moths Ephestia cautella and Plodia interpunctella. J. Chem. Ecol. 2005, 31, 2947–2961. [Google Scholar] [CrossRef] [PubMed]
- Nansen, C.; Phillips, T.W. Ovipositional Responses of the Indianmeal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) to Oils. Ann. Entomol. Soc. Am. 2003, 96, 524–531. [Google Scholar] [CrossRef]
- Li, C.; Cao, J.; Wang, X.; Xu, P.; Wang, X.; Ren, G. Efficacy of an improved method to screen semiochemicals of insect. PeerJ 2021, 9, e11510. [Google Scholar] [CrossRef]
- Xiang, L.; Zhang, X.G.; Chun, X.; Gao, Y.L.; Dong, W.X. Behavioral responses of potato tuber moth (Phthorimaea operculella) to tobacco plant volatiles. J. Integr. Agric. 2020, 19, 325–332. [Google Scholar]
- Schirack, A.; Drake, M.; Sanders, T.; Sandeep, K. Characterization of aroma-active compounds in microwave blanched peanuts. J. Food Sci. 2006, 71, C513–C520. [Google Scholar] [CrossRef]
- Sabier, M.; Wang, J.R.; Zhang, T.; Jin, J.D.; Wang, Z.J.; Shen, B.; Deng, J.; Liu, X.; Zhou, G. The attractiveness of a food-based lure and its component volatiles to the stored-grain pest Oryzaephilus surinamensis (L.). J. Stored Prod. Res. 2022, 98, 102000–102011. [Google Scholar] [CrossRef]
- Cao, Y.; Hu, Q.; Huang, L.; Athanassiou, C.G.; Maggi, F.; D’Isita, I.; Liu, Y.Y.; Pistillo, O.M.; Miao, M.Z.; Germinara, G.S.; et al. Attraction of Sitophilus oryzae (L.) (Coleoptera: Curculionidae) to the semiochemical volatiles of stored rice materials. J. Stored Prod. Res. 2024, 97, 73–85. [Google Scholar] [CrossRef]
- Adhikary, P.; Mukherjee, A.; Barik, A. Attraction of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) to four varieties of Lathyrus sativus L. seed volatiles. Bull. Entomol. Res. 2015, 105, 187–201. [Google Scholar] [CrossRef]
- Balakrishnan, K.; Holighaus, G.; Weissbecker, B.; Schutz, S. Electroantennographic responses of red flour beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) to volatile organic compounds. J. Appl. Entomol. 2017, 141, 477–486. [Google Scholar] [CrossRef]
- Fouad, H.A.; Faroni, L.R.D.; Vilela, E.F.; de Lima, E.R. Flight responses of Sitotroga cerealella (Lepidoptera: Gelechiidae) to corn kernel volatiles in a wind tunnel. Arthropod-Plant Interact. 2013, 7, 651–658. [Google Scholar] [CrossRef]
- Kwadha, C.A.; Mutunga, J.M.; Irungu, J.; Ongamo, G.; Ndegwa, P.; Raina, S.; Fombong, A.T. Decanal as a major component of larval aggregation pheromone of the greater wax moth, Galleria mellonella. J. Appl. Entomol. 2019, 143, 417–429. [Google Scholar] [CrossRef]
- Chen, Q.; Zhu, X.; Kang, G.; Yu, Q.; Liu, Q.; Du, L.; Yang, Y.; He, X.; Zhao, Y.; Zhang, J.; et al. Identification and functional characterization of female antennae-biased odorant receptor 23 involved in acetophenone detection of the Indian meal moth Plodia interpunctella. Insect Sci. 2024, 31, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Mattiolo, E.; Licciardello, F.; Lombardo, G.M.; Muratore, G.; Anastasi, U. Volatile profiling of durum wheat kernels by HS–SPME/GC–MS. Eur. Food Res. Technol. 2017, 243, 147–155. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peanuts Variety | Oleic Acid (%) | Linoleic Acid (%) | The Oleic/Linoleic Acid Ratio | Fat (%) | Protein (%) | Water (%) |
|---|---|---|---|---|---|---|
| Yuhua 9326 | 34.7 ± 1.9 | 35.7 ± 0.7 | 1.0 ± 0.1 | 49.3 ± 0.9 | 22.9 ± 0.8 | 7.1 ± 1.0 |
| Tianfu 3 | 37.2 ± 2.7 | 38.5 ± 2.1 | 1.1 ± 0.1 | 46.9 ± 1.2 | 26.5 ± 0.9 | 6.8 ± 0.6 |
| Pukehua 5 | 37.5 ± 3.1 | 39.3 ± 0.3 | 1.1 ± 0.1 | 54.6 ± 1.9 | 21.4 ± 1.0 | 7.0 ± 0.7 |
| Puhua 28 | 37.7 ± 4.23 | 40.1 ± 1.0 | 1.2 ± 0.1 | 50.5 ± 1.0 | 23.4 ± 0.4 | 6.2 ± 0.5 |
| Yuhanghua 1 | 39.2 ± 3.5 | 37.2 ± 0.4 | 1.2 ± 0.1 | 51.7 ± 0.3 | 21.8 ± 0.2 | 6.6 ± 0.2 |
| Jihua 13 | 80.5 ± 0.3 | 3.2 ± 5.0 | 19.4 ± 0.6 | 52.8 ± 1.0 | 23.9 ± 0.7 | 6.8 ± 0.2 |
| Kainong 301 | 80.5 ± 0.3 | 6.68 ± 2.6 | 17.5 ± 0.8 | 53.0 ± 0.7 | 21.0 ± 1.5 | 6.9 ± 0.1 |
| Jihua 11 | 81.3 ± 0.3 | 3.4 ± 4.8 | 22.2 ± 13.5 | 53.3 ± 0.8 | 26.1 ± 1.8 | 6.9 ± 0.3 |
| No. | VOCs | Retention Time | RI a | RI b | Relative Content (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yuhanghua 1 | Yuhua 9326 | Puhua28 | Pukehua 5 | Tianfu 3 | Jihua 13 | Kainong 301 | Jihua 11 | |||||
| 1 | 3-butyn-1-ol | 5.03 | 650 | 659 | — | — | — | 2.1 ± 0.6 a | 4.2 ± 1.1 a | — | — | — |
| 2 | 1-hexanol | 5.23 | 849 | 860 | — | — | 0.1 ± 0.0 b | 1.4 ± 0.1 a | — | — | — | |
| 3 | benzaldehyde | 7.54 | 968 | 982 | — | — | — | 0.4 ± 0.0 | — | — | — | — |
| 4 | 1-heptanol | 8.91 | 946 | 960 | — | — | — | 0.21 ± 0.0 | — | — | — | — |
| 5 | decane | 8.94 | 1004 | 1015 | — | — | — | 5.3 ± 1.4 | — | — | — | — |
| 6 | 6-methyl-5-hepten-2-one | 9.02 | 935 | 938 | — | — | — | 1.7 ± 0.2 | — | — | — | — |
| 7 | 1-ethyl-3-methylbenzene | 9.16 | 984 | 964 | 0.1 ± 0.0 | — | — | — | — | — | — | — |
| 8 | 1-octen-3-ol | 9.94 | 967 | 969 | — | — | — | — | 0.6 ± 0.0 | — | — | — |
| 9 | octanal | 9.93 | 1001 | 1005 | — | — | 2.0 ± 0.7 a | 0.3 ± 0.1 b | — | — | — | — |
| 10 | limonene | 10.04 | 1010 | 1018 | — | — | — | 1.2 ± 0.1 | — | — | — | — |
| 11 | hexanoic acid | 10.22 | 964 | 974 | — | — | — | 3.4 ± 1.0 b | 5.4 ± 1.8 a | — | — | — |
| 12 | 2,2,4,6,6—pentamethylheptane | 10.14 | 1004 | 997 | 0.8 ± 0.0 a | — | — | — | — | — | — | — |
| 13 | 4-methyldecane | 11.54 | 1033 | 1059 | 0.3 ± 0.0 a | — | — | — | — | — | — | 0.3 ± 0.0 a |
| 14 | 3,7-dimethyl-decane | 12.06 | 1043 | 1150 | 0.3 ± 0.1 f | 0.6 ± 0.1 e | 0.5 ± 0.0 e | 2.1 ± 0.1 c | 2.2 ± 1.0 b | — | 1.0 ± 0.1 d | 3.0 ± 0.5 a |
| 15 | undecane | 12.91 | 1061 | 1154 | 0.3 ± 0.0 b | — | — | — | — | 4.0 ± 0.6 a | — | 0.5 ± 0.1 b |
| 16 | acetophenone | 13.47 | 1072 | 1078 | 0.6 ± 0.2 a | 0.2 ± 0.0 b | — | — | — | — | — | — |
| 17 | nonanal | 14.91 | 1102 | 1102 | 0.5 ± 0.1 d | 0.9 ± 0.1 c | 0.4 ± 0.0 de | 5.7 ± 0.3 a | 1.5 ± 0.1 b | 0.3 ± 0.4 de | 0.2 ± 0.0 ef | 0.3 ± 0.0 e |
| 18 | benzoic acid | 17.03 | 1147 | 1150 | — | — | — | — | 1.4 ± 0.30 a | — | — | — |
| 19 | 4-ethyl-benzaldehyde | 16.55 | 1105 | 1108 | — | 2.7 ± 0.4 b | — | — | — | 3.3 ± 0.2 b | 25.3 ± 3.2 a | 2.8 ± 0.1 b |
| 20 | 2,5-dimethyl-benzaldehyde | 18.06 | 1122 | 1137 | — | — | — | — | — | — | — | 0.9 ± 0.2 a |
| 21 | butanedioic acid, diethyl ester | 18.09 | 1143 | 1151 | — | — | — | — | — | 1.5 ± 0.0 | — | — |
| 22 | dodecane | 18.31 | 1167 | 1214 | 1.8 ± 0.1 d | 2.5 ± 0.1 c | 9.1 ± 0.3 a | 7.3 ± 0.3 b | 1.6 ± 0.2 d | 1.2 ± 0.1 e | 0.9 ± 0.1 e | 1.0 ± 0.1 e |
| 23 | decanal | 18.69 | 1173 | 1161 | 0.3 ± 0.0 d | 0.5 ± 0.0 c | — | 1.8 ± 0.1 b | 2.2 ± 0.1 a | — | 0.1 ± 0.0 d | 0.2 ± 0.0 d |
| 24 | 4,6-dimethyl-undecane | 18.82 | 1176 | 1193 | 0.9 ± 0.2 | — | — | — | — | — | — | — |
| 25 | 4-methyl-dodecane | 18.91 | 1178 | 1249 | 1.1 ± 0.1 a | — | 0.3 ± 0.1 c | — | — | 0.5 ± 0.1 b | — | 0.45 ± 0.03 b |
| 26 | 2,4-dimethyl-benzaldehyde | 18.94 | 1179 | 1180 | 0.5 ± 0.2 a | 0.7 ± 0.1 a | — | — | — | — | — | — |
| 27 | isophthalaldehyde | 19.38 | 1210 | 1284 | — | 0.3 ± 0.0 a | — | — | — | — | — | — |
| 28 | tridecane | 20.64 | 1302 | 1313 | — | 0.7 ± 0.1 a | — | 2.6 ± 0.5 a | — | — | — | — |
| 29 | 5-methyl tetradecane | 20.62 | 1213 | 1454 | 0.3 ± 0.0 c | — | — | — | — | 1.0 ± 0.1 a | 0.7 ± 0.1 b | 0.8 ± 0.2 b |
| 30 | 3,4-dimethyl-phenylethanone | 20.72 | 1215 | 1255 | 3.0 ± 0.2 d | 5.1 ± 0.1 c | — | — | — | 8.4 ± 0.6 ab | 10.2 ± 0.9 a | 9.4 ± 0.5 ab |
| 31 | 4-ethylphenyl-ethanone | 21.37 | 1231 | 1242 | 1.5 ± 0.3 c | — | — | — | — | 4.6 ± 0.1 b | 7.6 ± 0.9 a | — |
| 32 | 5-methy-5-propylnonane | 21.49 | 1233 | 1229 | 0.1 ± 0.0 | — | — | — | — | — | — | — |
| 33 | 4,6-dimethyl-dodecane | 22.80 | 1264 | 1285 | 0.2 ± 0.0 d | 0.8 ± 0.1 c | 1.0 ± 0.1 bc | — | — | 2.7 ± 0.4 a | 1.3 ± 0.1 b | 1.3 ± 0.1 b |
| 34 | tetradecane | 25.29 | 1383 | 1413 | 0.3 ± 0.0 d | 0.6 ± 0.1 b | 0.5 ± 0.0 c | — | — | — | 1.0 ± 0.1 a | 0.4 ± 0.1 cd |
| 35 | 1,1′-(1,4-phenylene)bisethanone | 26.67 | 1400 | 1378 | 0.4 ± 0.2 e | 3.2 ± 0.1 b | — | — | — | 4.4 ± 0.1 a | 1.9 ± 0.1 d | 2.3 ± 0.1 c |
| 36 | 2,4-di-tert-butylphenol | 30.07 | 1455 | 1494 | 0.7 ± 0.1 c | 2.1 ± 0.1 b | — | — | — | 3.2 ± 0.3 a | 2.1 ± 0.1 b | 0.7 ± 0.1 c |
| 37 | hexadecane | 33.21 | 1560 | 1612 | 0.7 ± 0.1 d | 0.5 ± 0.1 d | 11.1 ± 1.3 a | — | 1.7 ± 0.2 bc | 1.2 ± 0.1 cd | 0.5 ± 0.1 d | 2.3 ± 0.2 b |
| 38 | heptadecane | 36.93 | 1685 | 1711 | 1.0 ± 0.1 c | 0.4 ± 0.1 e | 0.7 ± 0.1 d | — | — | 2.3 ± 0.1 b | 2.4 ± 0.1 a | 0.5 ± 0.1 e |
| 39 | eicosane | 37.02 | 2002 | 2009 | 0.3 ± 0.0 e | 0.4 ± 0.1 d | — | — | — | 2.6 ± 0.1 a | 1.5 ± 0.1 b | 1.0 ± 0.1 c |
| 40 | dibutyl phthalate | 44.80 | 2168 | 2037 | 0.1 ± 0.0 d | — | — | — | 0.4 ± 0.1 c | — | 1.2 ± 0.2 a | 0.9 ± 0.2 b |
| 41 | 2-methyl-hexacosane | 51.43 | 2617 | 2656 | 0.1 ± 0.0 | — | — | — | — | — | — | — |
| 42 | phthalic acid, di(2-propylpentyl) ester | 52.23 | 2689 | 2704 | — | — | — | 0.1 ± 0.0 b | — | — | — | 0.6 ± 0.1 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Wang, D.; Zeng, F.; Chen, L.; Zhao, X.; Zhu, X.; Yao, J.; Li, Y. Identification on Key Volatiles Contributed to Oviposition Preference of Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae) from High and Normal Oleic Varieties of Peanut. Insects 2024, 15, 866. https://doi.org/10.3390/insects15110866
Wang C, Wang D, Zeng F, Chen L, Zhao X, Zhu X, Yao J, Li Y. Identification on Key Volatiles Contributed to Oviposition Preference of Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae) from High and Normal Oleic Varieties of Peanut. Insects. 2024; 15(11):866. https://doi.org/10.3390/insects15110866
Chicago/Turabian StyleWang, Chen, Dianxuan Wang, Fangfang Zeng, Liang Chen, Xinxin Zhao, Xi Zhu, Junji Yao, and Yihan Li. 2024. "Identification on Key Volatiles Contributed to Oviposition Preference of Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae) from High and Normal Oleic Varieties of Peanut" Insects 15, no. 11: 866. https://doi.org/10.3390/insects15110866
APA StyleWang, C., Wang, D., Zeng, F., Chen, L., Zhao, X., Zhu, X., Yao, J., & Li, Y. (2024). Identification on Key Volatiles Contributed to Oviposition Preference of Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae) from High and Normal Oleic Varieties of Peanut. Insects, 15(11), 866. https://doi.org/10.3390/insects15110866