
Reproductive Behavior and Development of the Global Insect Pest, Cotton Seed Bug Oxycarenus hyalinipennis

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Experimental Design and Procedure
2.2.1. Oxycarenus hyalinipennis Copulation: Frequency, Latency, and Duration Analysis
2.2.2. Egg-Laying Patterns in Virgin and Mated Female O. hyalinipennis
2.2.3. Development of O. hyalinipennis Life Stages after Egg-Laying
2.2.4. Longevity of Adult Male and Female O. hyalinipennis
2.3. Data Analysis
3. Results
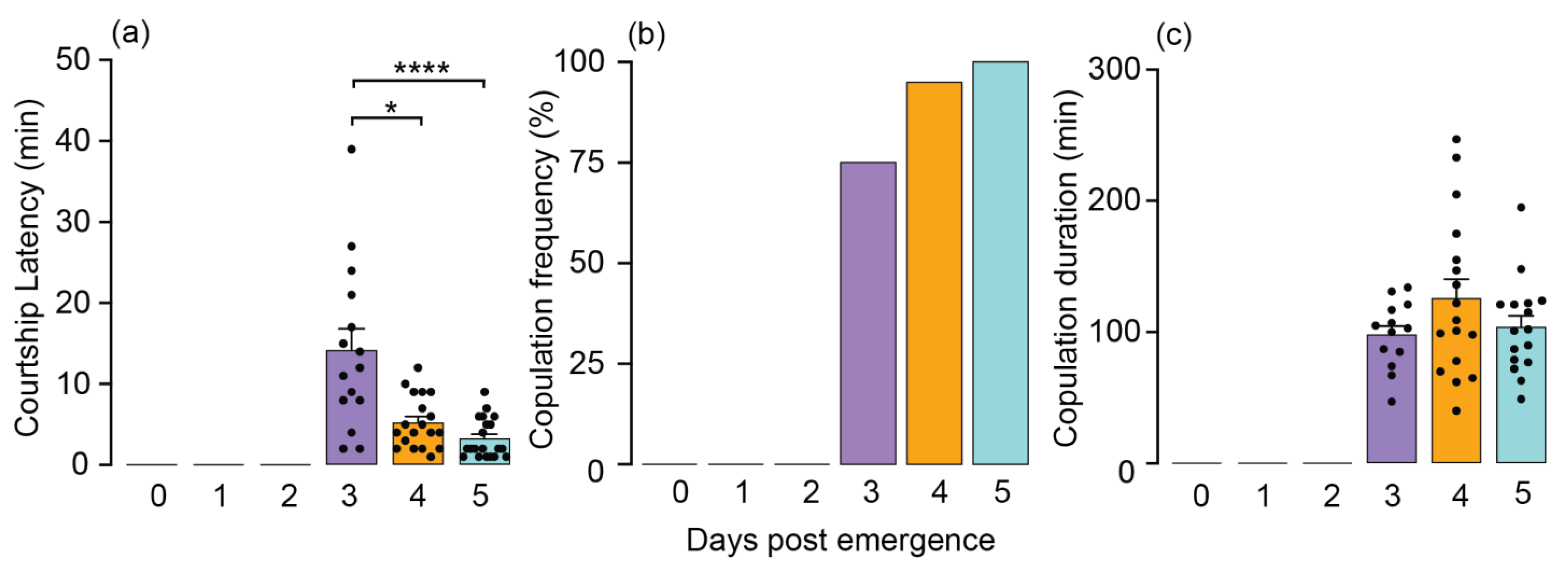
3.1. Age-Dependent Effects on O. hyalinipennis Copulation Behavior
3.2. Egg-Laying Patterns in Unmated and Mated Females
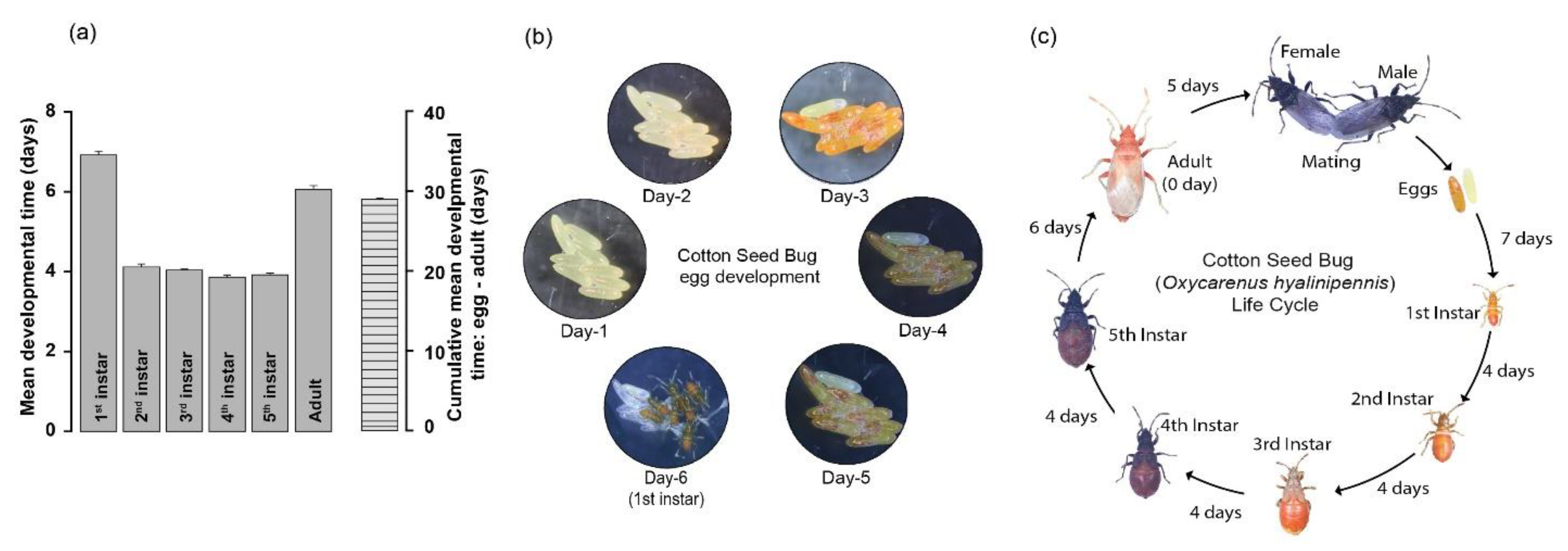
3.3. Development of O. hyalinipennis Life Strategy
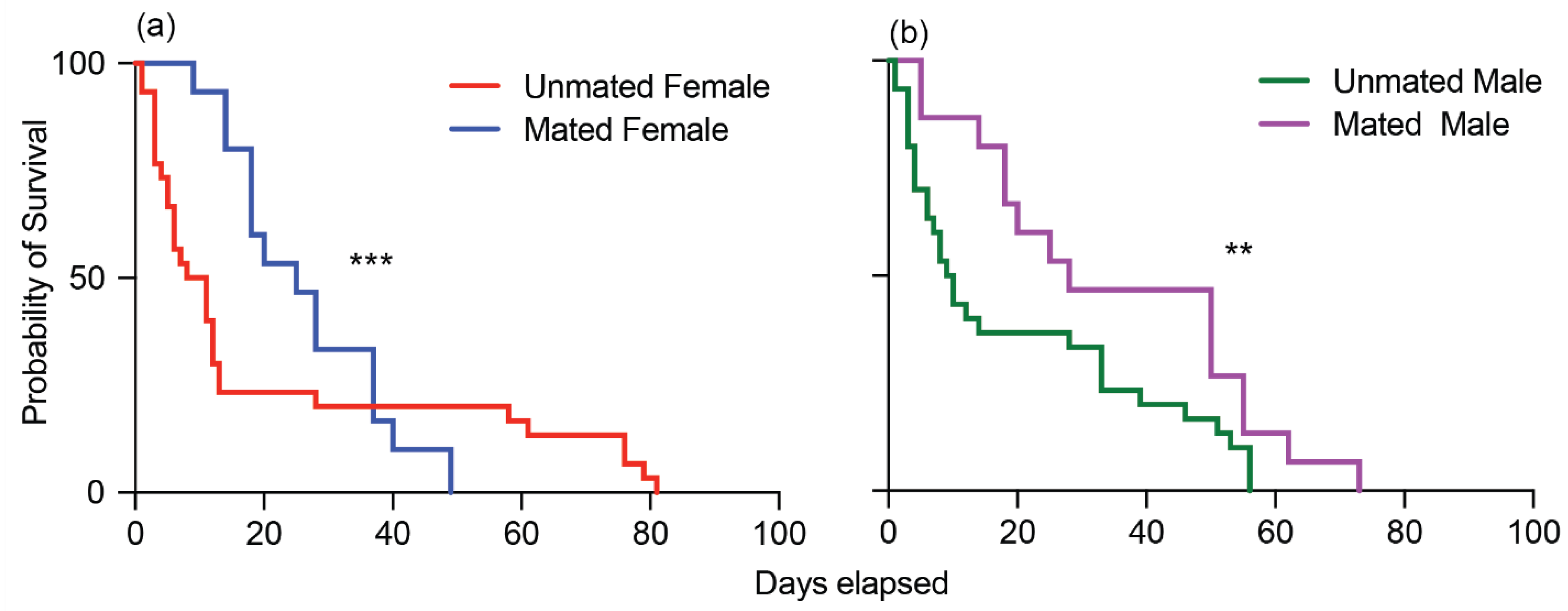
3.4. Longevity of Adult Male and Female Cotton Seed Bugs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Renou, M.; Guerrero, A. Insect Pheromones in Olfaction Research and Semiochemical-Based Pest Control Strategies. Annu. Rev. Entomol. 2000, 48, 605–630. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, M.L.; Sunil, V. Basic Behavioural Patterns in Insects and Applications of Behavioural Manipulation in Insect Pest Management. J. Entomol. Zool. Stud. 2020, 8, 991–996. [Google Scholar]
- Foster, S.P.; Harris, M.O. Behavioral Manipulation Methods for Insect Pest-Management. Annu. Rev. Entomol. 1997, 42, 123–169. [Google Scholar] [CrossRef] [PubMed]
- Slater, A.J.; Baranowski, M.R. The Occurrence of Oxycarenus hyalinipennis (Costa) (Hemiptera: Lygaeidae) in the West Indies and New Lygaeidae Records for the Turks and Caicos Islands of Providenciales and North Caicos. Fla. Entomol. 1994, 77, 495–497. [Google Scholar] [CrossRef]
- Smith, T.R.; Brambila, J. A Major Pest of Cotton, Oxycarenus hyalinipennis (Heteroptera: Oxycarenidae) in the Bahamas. Fla. Entomol. 2008, 91, 479–482. [Google Scholar] [CrossRef]
- Bolu, H.; Dioli, P.; Çelik, H. Various Observations on Some Biological Character of Oxycarenus hyalinipennis (A. Costa, 1843) (Hemiptera: Lygaeoidea: Oxycarenidae) in South-Eastern Turkey. Munis Entomol. Zool. J. 2020, 15, 481–488. [Google Scholar]
- Ananthakrishnanan, T.N.; Raman, K.; Sanjayan, K.P. Comparative Growth Rate, Fecundity and Behavioral Diversity of the Dusky Cotton Bug, Oxycarenus hyalinipennis (Costa) (Hemiptera; Lygaeidae) on Certain Malvaceous Host Plants. Proc. Indian Nat. Sci. Acad. 1982, 48, 577–584. [Google Scholar]
- Ullah, S.; Shad, S.A. Toxicity of Insecticides, Cross-Resistance and Stability of Chlorfenapyr Resistance in Different Strains of Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae). Crop Prot. 2017, 99, 132–136. [Google Scholar] [CrossRef]
- Atta, B.; Gogi, M.D.; Arif, M.J.; Mustafa, F.; Raza, M.F.; Hussain, M.J.; Farooq, M.A.; Nisar, M.J.; Iqbal, M. Toxicity of Come Insect Growth Regulators (IGRS) against different Life Stages of Dusky Cotton Bugs Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae: Oxycareninae). Bulg. J. Agric. Sci. 2015, 21, 367–371. [Google Scholar]
- Adu-Mensah, K.; Kumar, R. Ecology of Oxycarenus Species (Heteroptera: Lygaeidae) in Southern Ghana. Biol. J. Linn. Soc. 1977, 9, 349–377. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Francis Borgio, J. Virulence of Entomopathogenic Fungus Metarhizium anisopliae (Metsch.) Sorokin on Seven Insect Pests. Indian J. Agric. Res. 2010, 44, 195–200. [Google Scholar]
- Brent, C.S. Reproduction of the Western Tarnished Plant Bug, Lygus hesperus, in Relation to Age, Gonadal Activity and Mating Status. J. Insect Physiol. 2010, 56, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, L.; Burnep, B. Courtship Latency in Male Drosophila melanogaster. Behav. Genet. 1977, 7, 359–372. [Google Scholar] [CrossRef]
- Ridley, M. Mating Frequency and Fecundity in Insects. Bological Rev. 1988, 63, 509–549. [Google Scholar] [CrossRef]
- Gillott, C.; Langley, P.A. The Control of Receptivity and Ovulation in the Tsetse Fly, Glossina morsitans. Physiol. Entomol. 1981, 6, 269–281. [Google Scholar] [CrossRef]
- Melo, G.A.R.; Buschini, M.L.T.; Campos, L.A.O. Ovarian Activation in Melipona quadrifasciata Queens by Mating Plug Stimulation (Hymenoptera, Apidae). Apidologie 2001, 32, 355–361. [Google Scholar] [CrossRef]
- Heifetz, Y.; Lung, O.; Frongillo, E.A.; Wolfner, M.F. The Drosophila Seminal Fluid Protein Acp26Aa Stimulates Release of Oocytes by the Ovary. Curr. Biol. 2000, 10, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.Y.; Gong, H. Male Accessory Gland Derived Factors Can Stimulate Oogenesis and Enhance Oviposition in Helicoverpa armigera (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2001, 46, 175–185. [Google Scholar] [CrossRef]
- Leahy, M.G. Qviposition of Schistocerca gregaria (Forskal) (Orthoptera: Acrididae) Mated with Males Unable to Transfer Spermatophores. J. Entomol. Ser. A Gen. Entomol. 1973, 48, 79–84. [Google Scholar] [CrossRef]
- Ugine, T.A. Developmental Times and Age-Specific Life Tables for Lygus lineolaris (Heteroptera: Miridae), Reared at Multiple Constant Temperatures. Environ. Entomol. 2012, 41, 1–10. [Google Scholar] [CrossRef]
- Arnqvist, G.; Nilsson, T. The Evolution of Polyandry: Multiple Mating and Female Fitness in Insects. Anim. Behav. 2000, 60, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Wolfner, M.F. Tokens of Love: Functions and Regulation of Drosophila Male Accessory Gland Products. Insect Biochem. Mol. Biol. 1997, 27, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.; Liddle, L.F.; Kalb, J.M.; Wolfner, M.F.; Partridge, L. Cost of Mating in Drosophila melanogaster Females Is Mediated by Male Accessory Gland Products. Nature 1995, 373, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Saveer, A.M.; Becher, P.G.; Birgersson, G.; Hansson, B.S.; Witzgall, P.; Bengtsson, M. Mate Recognition and Reproductive Isolation in the Sibling Species Spodoptera littoralis and Spodoptera litura. Front. Ecol. Evol. 2014, 2, 18. [Google Scholar] [CrossRef]
- Villarreal, S.M.; Pitcher, S.; Helinski, M.E.H.; Johnson, L.; Wolfner, M.F.; Harrington, L.C. Male Contributions during Mating Increase Female Survival in the Disease Vector Mosquito Aedes aegypti. J. Insect. Physiol. 2018, 108, 1–9. [Google Scholar] [CrossRef]
- Saveer, A.M.; DeVries, Z.C.; Santangelo, R.G.; Schal, C. Mating and Starvation Modulate Feeding and Host-Seeking Responses in Female Bed Bugs, Cimex lectularius. Sci. Rep. 2021, 11, 1915. [Google Scholar] [CrossRef]
- Cook, J.C.; Funaro, C.F.; Harding, T.C.; McPhie, D. Technical Bulletin—Oxycarenus hyalinipennis (COSTA) (Hemiptera: Oxycarenidae) Cotton Seed Bug; Animal and Plant Health Inspection Service Plant Protection and Quarantine; United States Department of Agriculture: Washington, DC, USA, 2021. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saveer, A.M.; Hu, J.; Strickland, J.; Krueger, R.; Clafford, S.; Zhang, A. Reproductive Behavior and Development of the Global Insect Pest, Cotton Seed Bug Oxycarenus hyalinipennis. Insects 2024, 15, 65. https://doi.org/10.3390/insects15010065
Saveer AM, Hu J, Strickland J, Krueger R, Clafford S, Zhang A. Reproductive Behavior and Development of the Global Insect Pest, Cotton Seed Bug Oxycarenus hyalinipennis. Insects. 2024; 15(1):65. https://doi.org/10.3390/insects15010065
Chicago/Turabian StyleSaveer, Ahmed M., Jing Hu, Jaime Strickland, Robert Krueger, Shannon Clafford, and Aijun Zhang. 2024. "Reproductive Behavior and Development of the Global Insect Pest, Cotton Seed Bug Oxycarenus hyalinipennis" Insects 15, no. 1: 65. https://doi.org/10.3390/insects15010065
APA StyleSaveer, A. M., Hu, J., Strickland, J., Krueger, R., Clafford, S., & Zhang, A. (2024). Reproductive Behavior and Development of the Global Insect Pest, Cotton Seed Bug Oxycarenus hyalinipennis. Insects, 15(1), 65. https://doi.org/10.3390/insects15010065