
Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae)

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
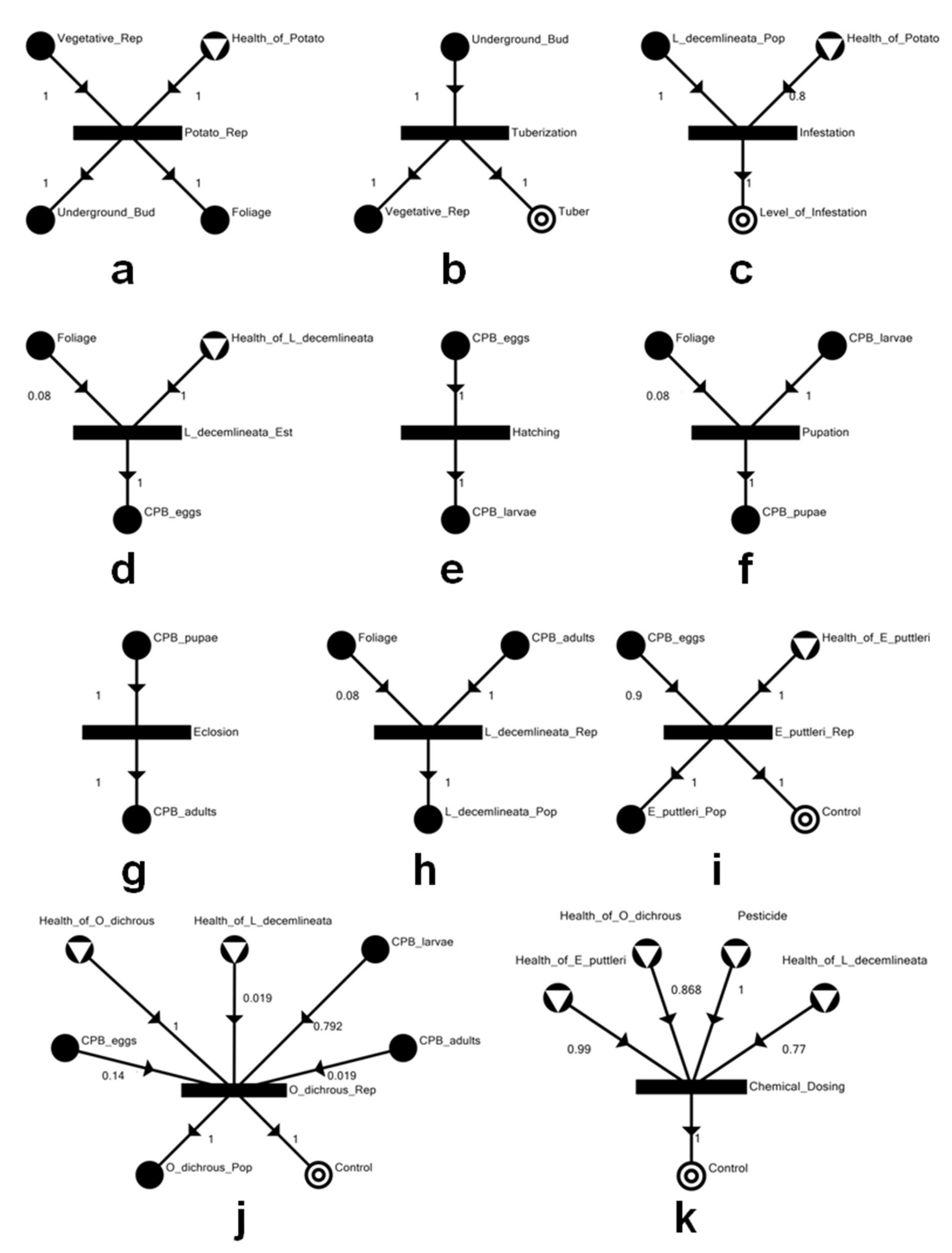
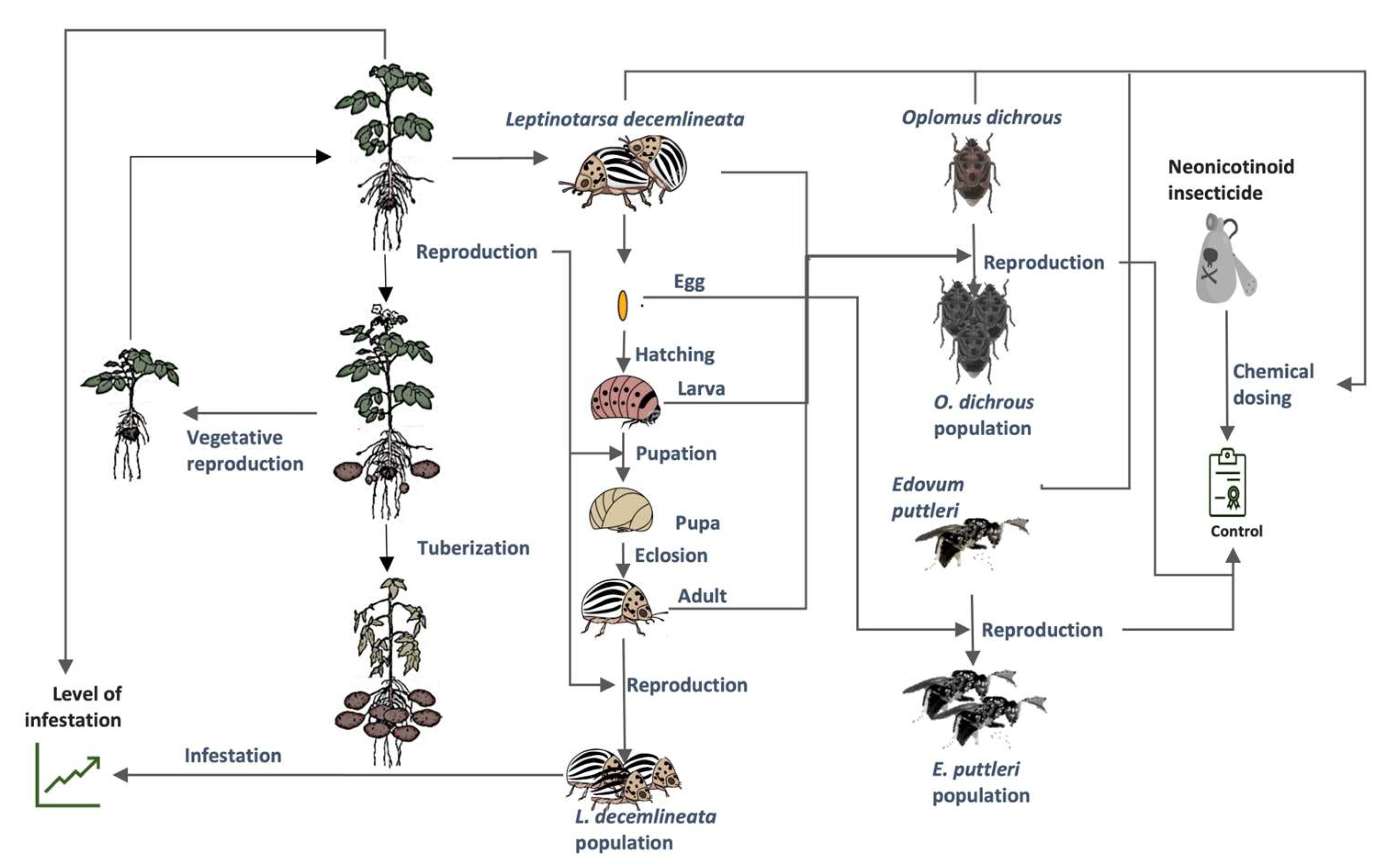
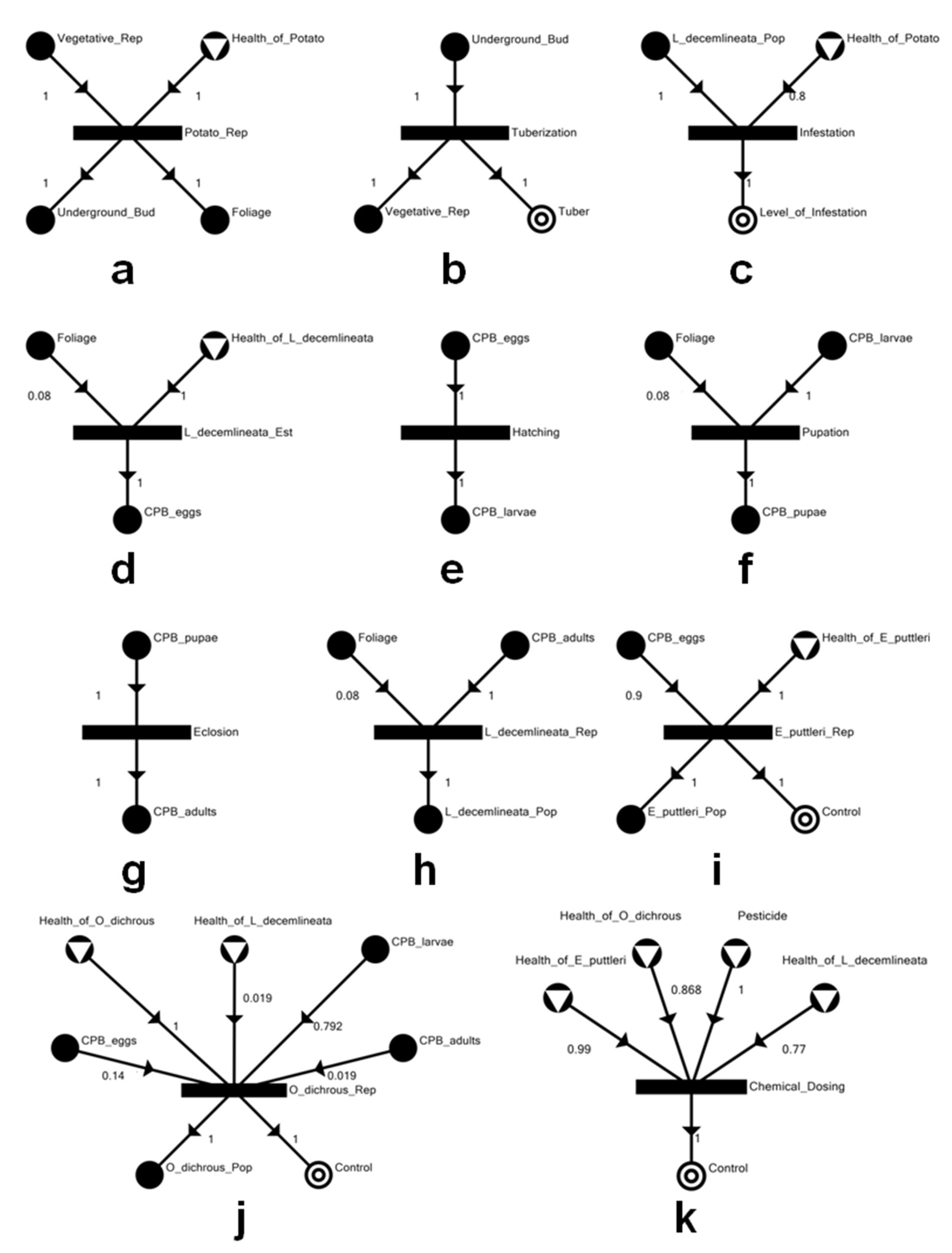
2.1. Model Development
2.2. Network Analysis
3. Results
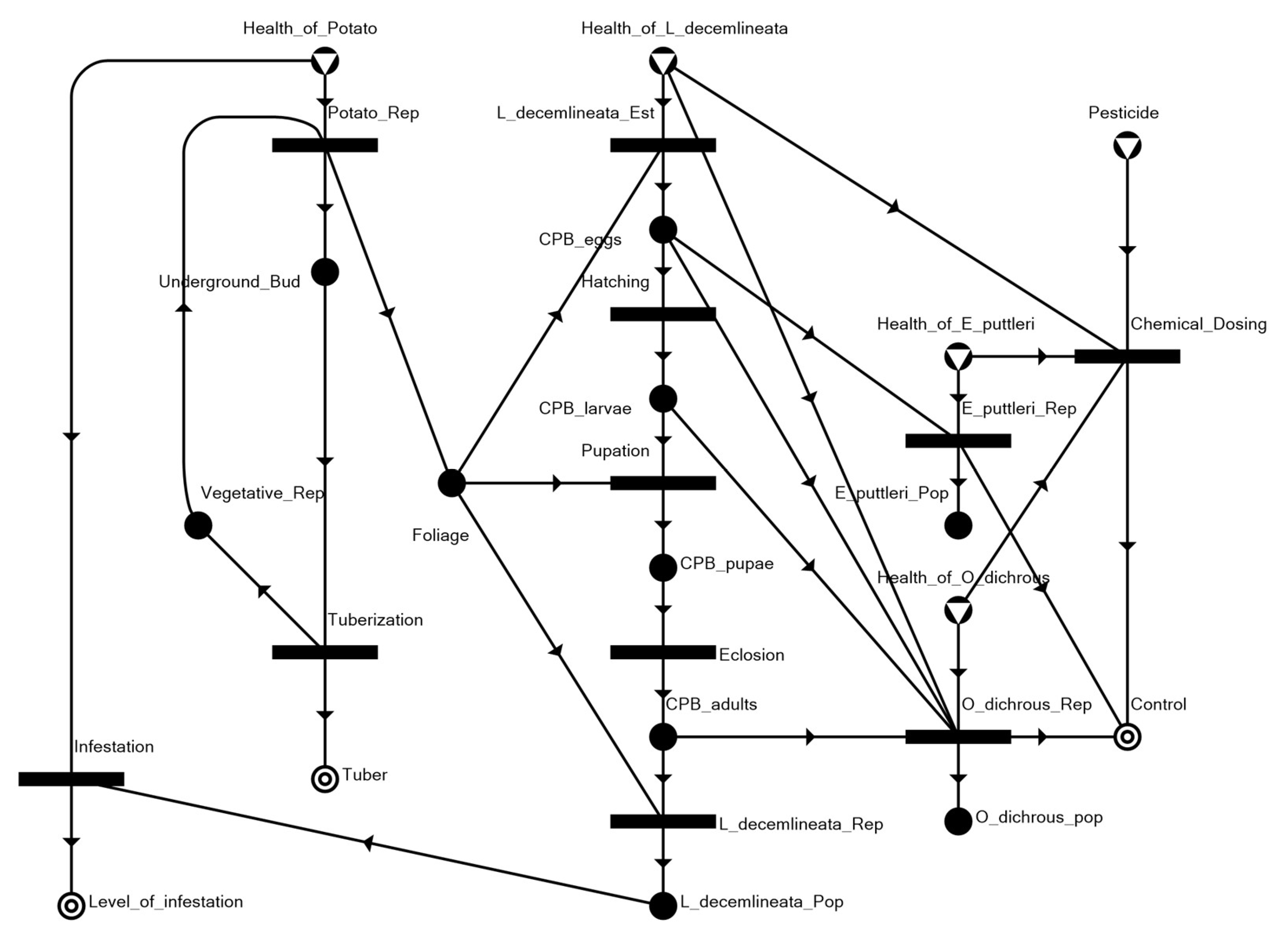
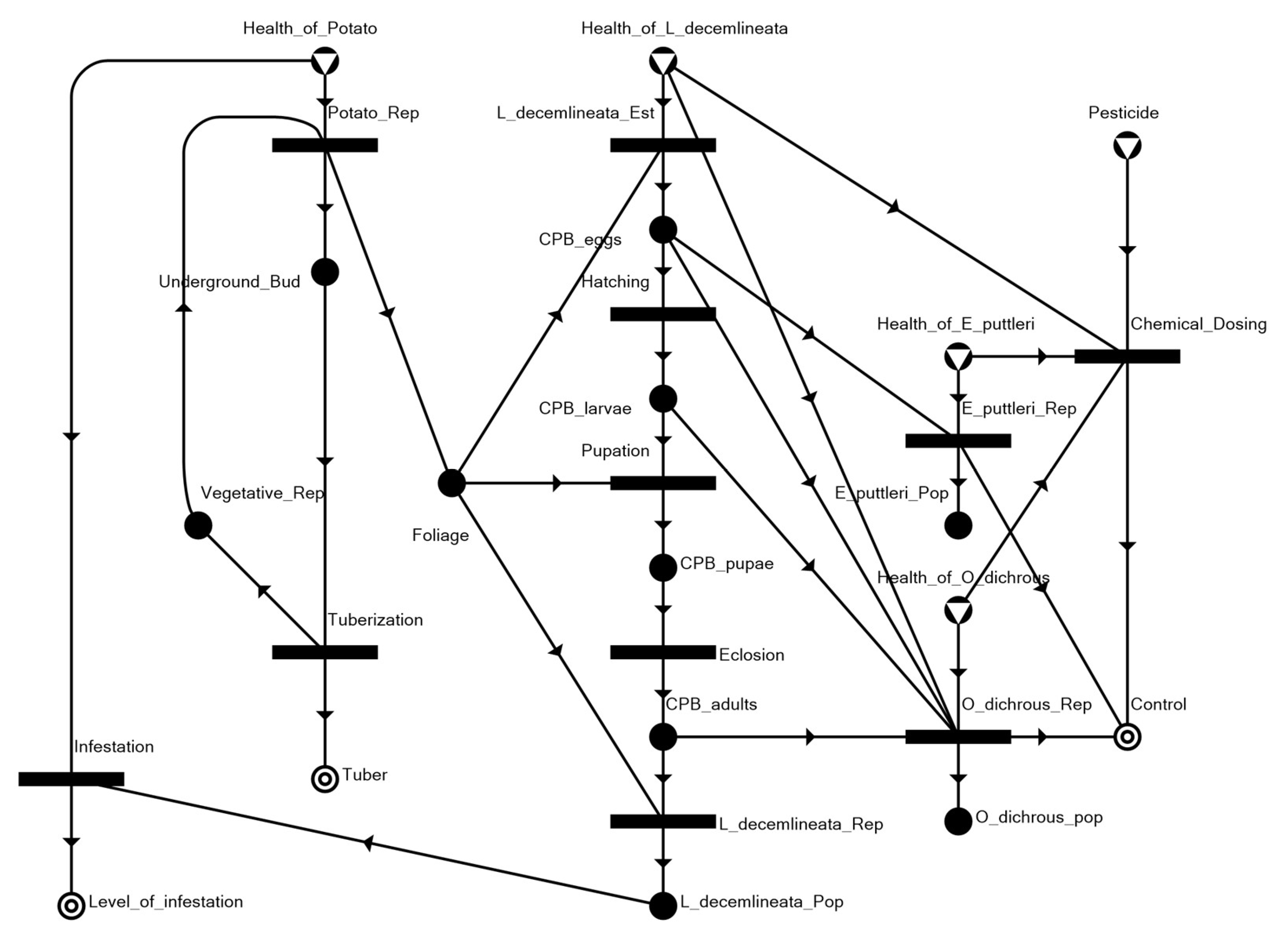
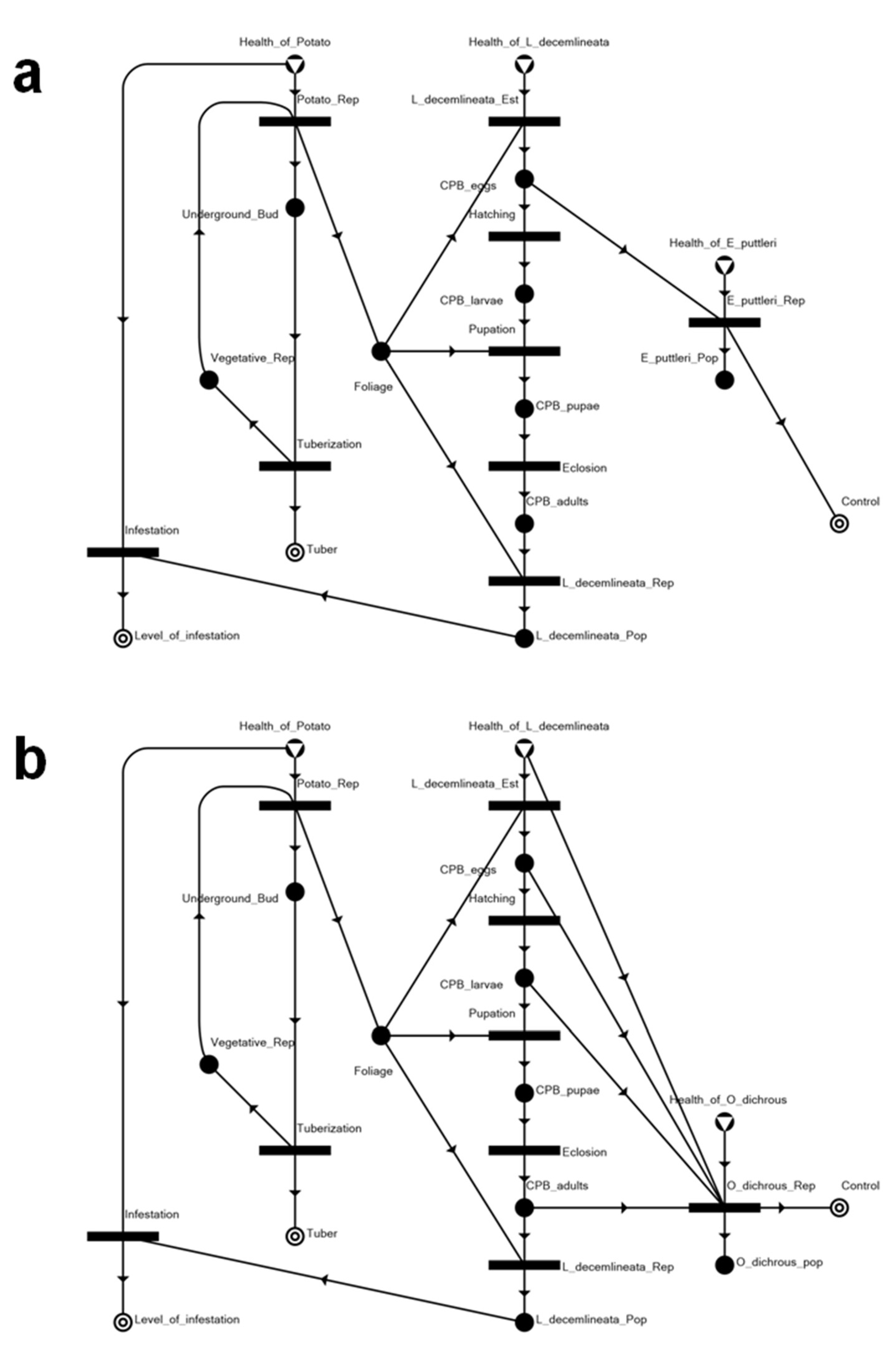
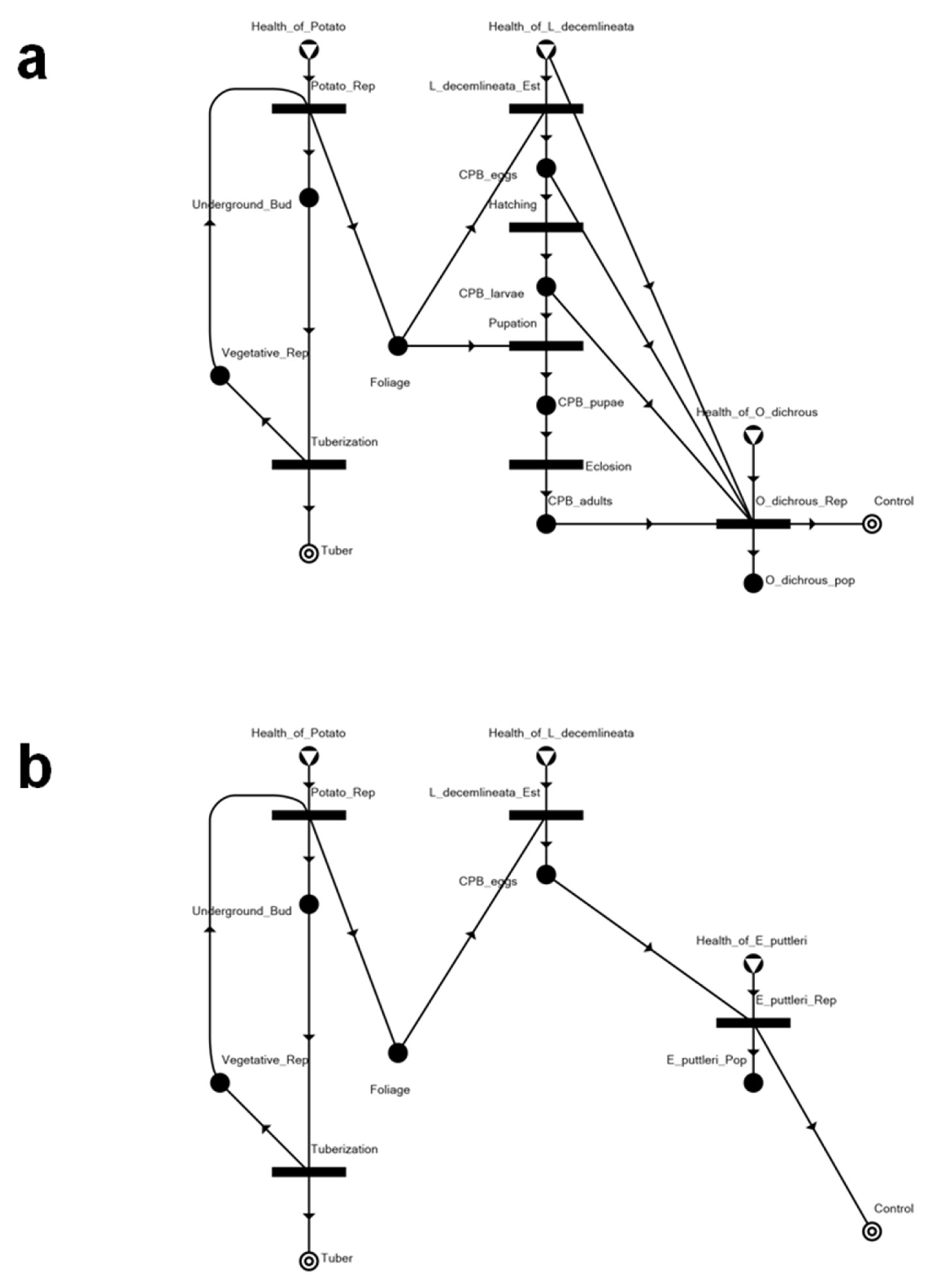
3.1. The Potato–CPB Agroecosystem Model
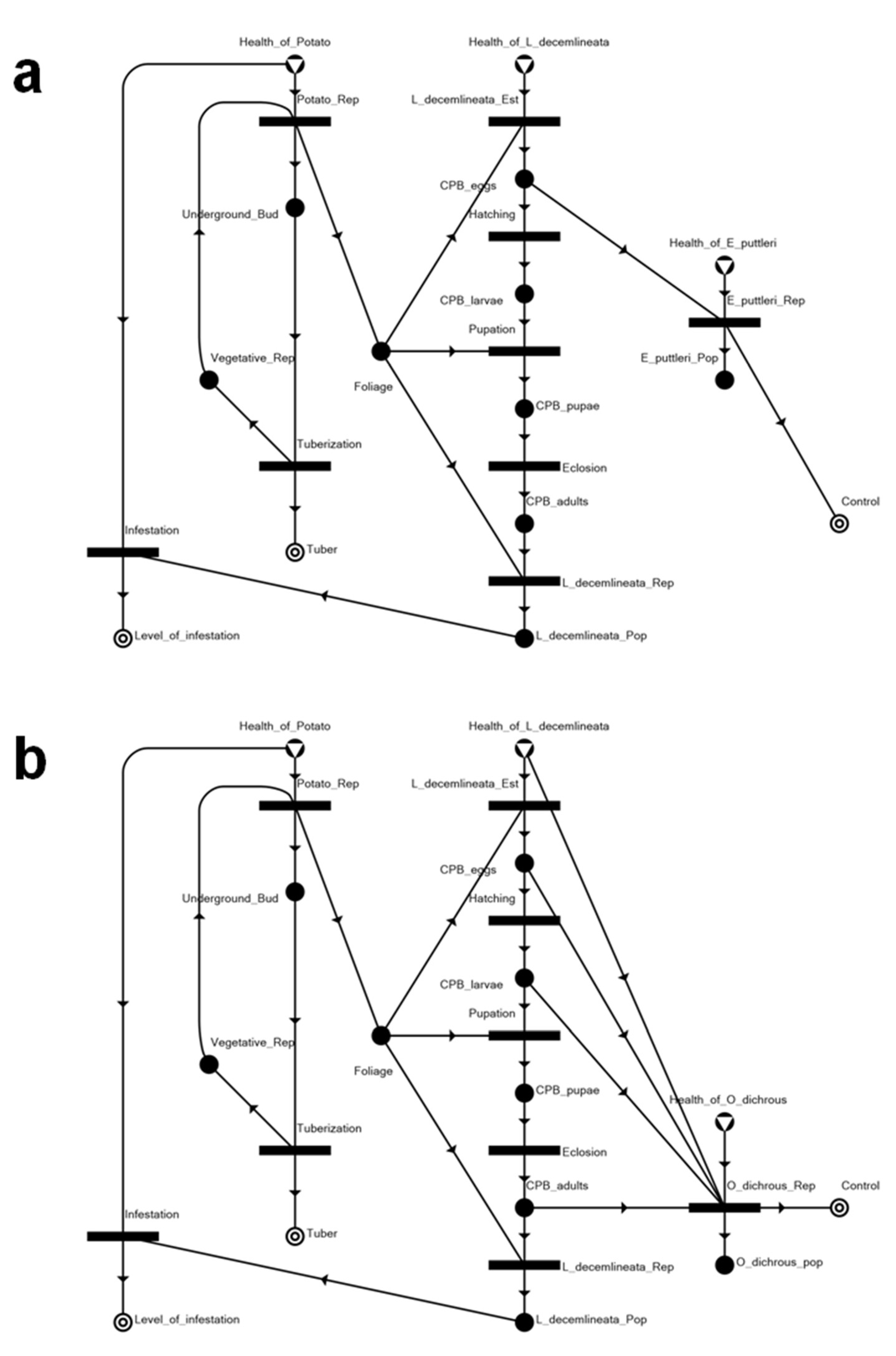
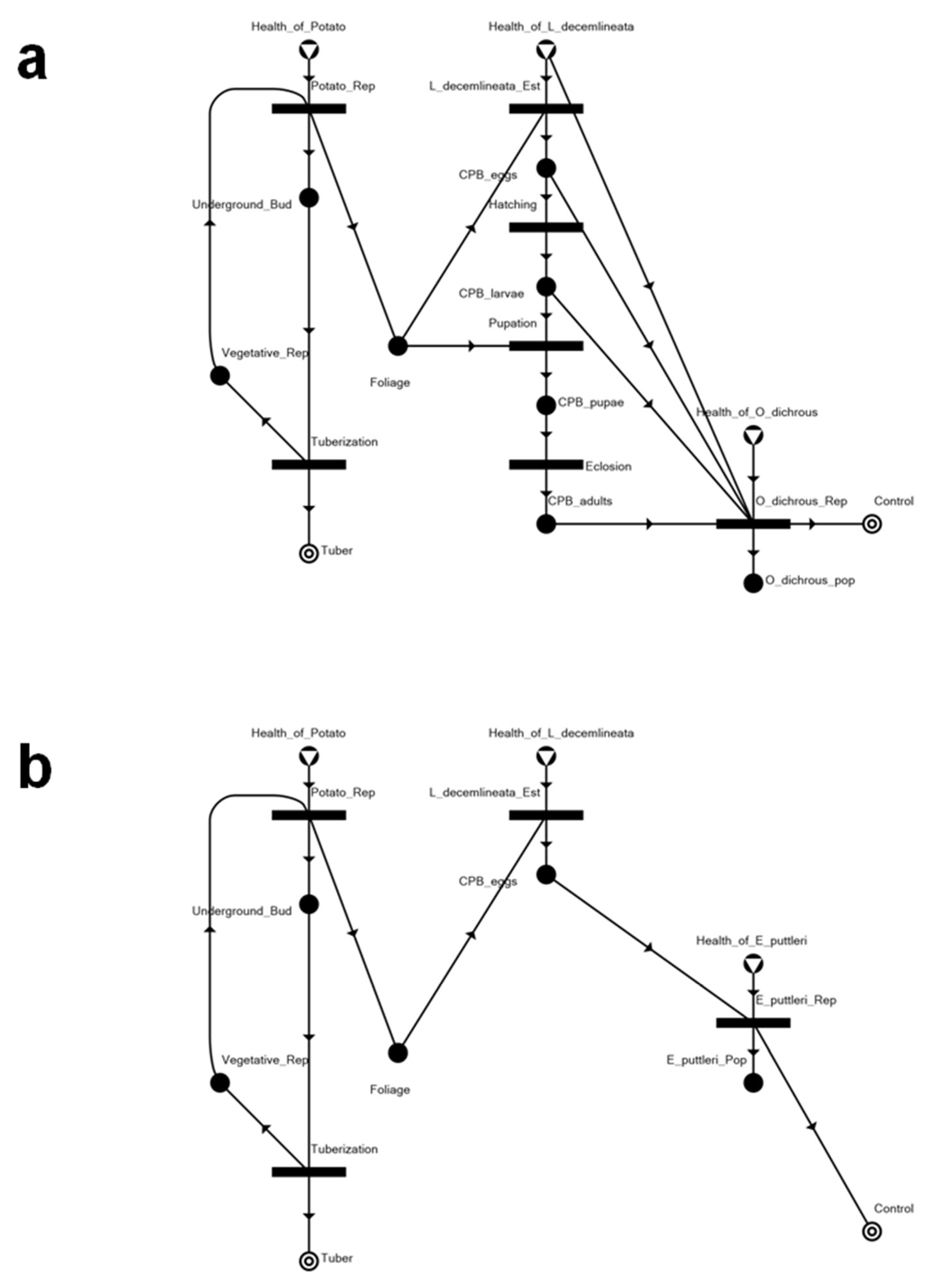
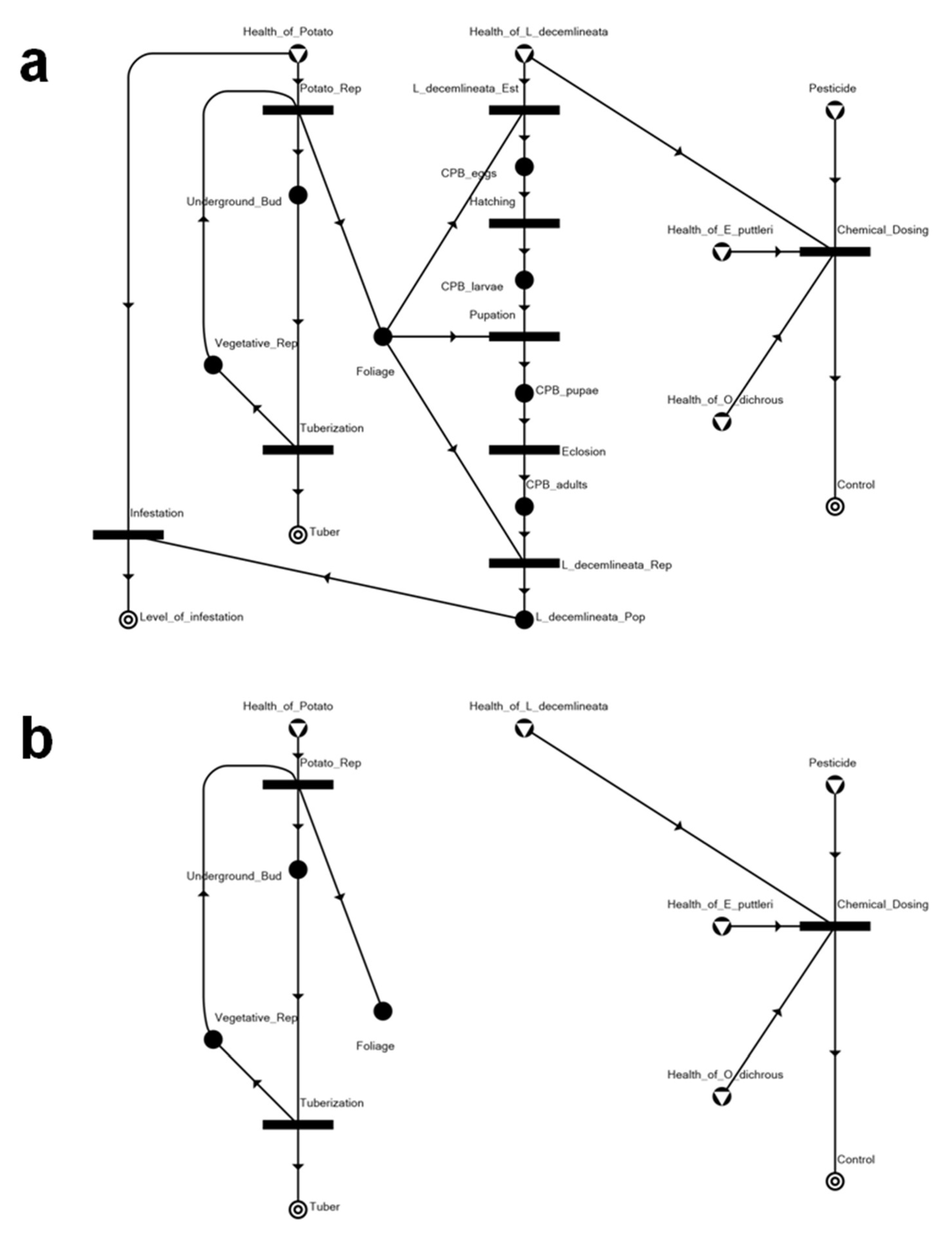
3.2. Solution Structures and Represented Scenarios
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bezner Kerr, R.; Hasegawa, T.; Lasco, R.; Bhatt, I.; Deryng, D.; Farrell, A.; Gurney-Smith, H.; Ju, H.; Lluch-Cota, S.; Meza, F.; et al. Food, fibre, and other ecosystem products. In Climate Change 2022: Impacts, Adaptation and Vulnerability: Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; pp. 713–906. [Google Scholar] [CrossRef]
- Fu, B.; Gasser, T.; Li, B.; Tao, S.; Ciais, P.; Piao, S.; Balkanski, Y.; Li, W.; Yin, T.; Han, L.; et al. Short-lived climate forcers have long-term climate impacts via the carbon–climate feedback. Nat. Clim. Chang. 2020, 10, 851–855. [Google Scholar] [CrossRef]
- Lenton, T.M.; Rockström, J.; Gaffney, O.; Rahmstorf, S.; Richardson, K.; Steffen, W.; Schellnhuber, H.J. Climate tipping points—Too risky to bet against. Nature 2019, 575, 592–595. [Google Scholar] [CrossRef] [PubMed]
- Brovkin, V.; Brook, E.; Williams, J.W.; Bathiany, S.; Lenton, T.M.; Barton, M.; DeConto, R.M.; Donges, J.F.; Ganopolski, A.; McManus, J.; et al. Past abrupt changes, tipping points and cascading impacts in the Earth system. Nat. Geosci. 2021, 14, 550–558. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Leši, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Senate of the Philippines. Press Release—Pangilinan: Use Anti-Cocolisap Template to Solve ‘Harabas’ Onion Pest Problem. Available online: https://legacy.senate.gov.ph/press_release/2022/0412_pangilinan1.asp (accessed on 10 October 2022).
- Wang, C.; Hawthorne, D.; Qin, Y.; Pan, X.; Li, Z.; Zhu, S. Impact of climate and host availability on future distribution of Colorado potato beetle. Sci. Rep. 2017, 7, 4489. [Google Scholar] [CrossRef] [PubMed]
- Co’s Digital Flora of the Philippines. Solanaceae. Available online: https://www.philippineplants.org/Families/Solanaceae.html (accessed on 8 February 2023).
- CABI International. ‘Leptinotarsa decemlineata (Colorado Potato Beetle)’, CABI Compendium. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.30380 (accessed on 6 October 2022).
- Gonzales, I.C.; Kiswa, C.G.; Bautista, A.B. Sustainable potato production in the Philippine Cordillera Region. Int. J. Eng. App. Sci. 2016, 3, 29–37. [Google Scholar]
- National Tobacco Association. Frequently Asked Questions. Available online: https://nta.da.gov.ph/faq.html (accessed on 8 February 2023).
- International Potato Center. Regional Congress Highlights Food Security and Resilience Benefits of Root and Tuber Crops. Available online: https://cipotato.org/pressreleases/dost-pcaarrd-cip-host-regional-congress-root-tuber-crops/ (accessed on 8 February 2023).
- Friedler, F.; Orosz, Á.; Pimentel Losada, J. P-graphs for Process Systems Engineering: Mathematical Models and Algorithms; Springer Nature: Cham, Switzerland, 2022; p. 261. [Google Scholar] [CrossRef]
- Friedler, F.; Tarjan, K.; Huang, Y.W.; Fan, L.T. Graph-theoretic approach to process synthesis: Axioms and theorems. Chem. Eng. Sci. 1992, 47, 1973–1988. [Google Scholar] [CrossRef]
- Almarinez, B.J.M.; Amalin, D.M.; Aviso, K.B.; Cabezas, H.; Lao, A.R.; Tan, R.R. Modelling network effects of biological control: The case of the Philippine coconut scale insect outbreak. BioControl 2023, 68, 117–130. [Google Scholar] [CrossRef]
- Lao, A.R.; Aviso, K.B.; Cabezas, H.; Tan, R.R. Maintaining the productivity of co-culture systems in the face of environmental change. Nat. Sustain. 2022, 5, 749–752. [Google Scholar] [CrossRef]
- Lao, A.; Cabezas, H.; Orosz, A.; Friedler, F.; Tan, R. Socio-ecological network structures from process graphs. PLoS ONE 2020, 15, e0232384. [Google Scholar] [CrossRef]
- Thornton, M. Potato Growth and Development. In Potato Production Systems; Stark, J.C., Thornton, M., Nolte, P., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 19–33. [Google Scholar]
- CABI International. ‘Edovum puttleri’, CABI Compendium. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.20479 (accessed on 6 October 2022).
- CABI International. ‘Oplomus dichrous’, CABI Compendium. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.37669 (accessed on 6 October 2022).
- Lashomb, J.; Krainacker, D.; Jansson, R.K.; Ng, Y.S.; Chianese, R. Parasitism of Leptinotarsa decemlineata (Say) eggs by Edovum puttleri Grisell (Hymenoptera: Eulophidae): Effects of host age, parasitoid age, and temperature. Can. Entomol. 1987, 119, 75–82. [Google Scholar] [CrossRef]
- Drummond, F.A.; Casagrande, R.A.; Groden, E. Biology of Oplomus dichrous (Heteroptera: Pentatomidae) and its potential to control Colorado potato beetle (Coleoptera: Chrysomelidae). Environ. Entomol. 1987, 16, 633–638. [Google Scholar] [CrossRef]
- Scott, I.M.; Tolman, J.H.; MacArthur, D.C. Insecticide resistance and cross-resistance development in Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) populations in Canada 2008–2011. Pest Manag. Sci. 2015, 71, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.B.; Silva-Torres, C.S.A.; Barros, R. Relative effects of the insecticide thiamethoxam on the predator Podisus nigrispinus and the tobacco whitefly Bemisia tabaci in nectaried and nectariless cotton. Pest Manag. Sci. 2003, 59, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Maharijaya, A.; Vosman, B. Managing the Colorado potato beetle; the need for resistance breeding. Euphytica 2015, 204, 487–501. [Google Scholar] [CrossRef]
- Varenhorst, A.; Bachmann, A.; Rozeboom, P.; Wagner, P.; South Dakota State University Extension. Colorado Potato Beetles Causing Defoliation on Potatoes. Available online: https://extension.sdstate.edu/colorado-potato-beetles-causing-defoliation-potatoes (accessed on 6 October 2022).
- Ragsdale, D.; Radcliffe, E.; VegEdge, University of Minnesota. Colorado Potato Beetle. Available online: https://vegedge.umn.edu/insect-pest-profiles/colorado-potato-beetle (accessed on 6 October 2022).
- Friedler, F.; Tarjan, K.; Huang, Y.W.; Fan, L.T. Graph-theoretic approach to process synthesis: Polynomial algorithm for maximal structure generation. Comput. Chem. Eng. 1993, 17, 929–942. [Google Scholar] [CrossRef]
- Alyokhin, A.; Dively, G.; Patterson, M.; Castaldo, C.; Rogers, D.; Mahoney, M.; Wollam, J. Resistance and cross-resistance to imidacloprid and thiamethoxam in the Colorado potato beetle Leptinotarsa decemlineata. Pest Manag. Sci. 2007, 63, 32–41. [Google Scholar] [CrossRef]
- Huseth, A.S.; Groves, R.L.; Chapman, S.A.; Alyokhin, A.; Kuhar, T.P.; Macrae, I.V.; Szendrei, Z.; Nault, B.A. Managing Colorado potato beetle insecticide resistance: New tools and strategies for the next decade of pest control in potato. J. Integr. Pest Manag. 2014, 5, A1–A8. [Google Scholar] [CrossRef]
- Clements, J.; Sanchez-Sedillo, B.; Bradfield, C.A.; Groves, R.L. Transcriptomic analysis reveals similarities in genetic activation of detoxification mechanisms resulting from imidacloprid and chlorothalonil exposure. PLoS ONE 2018, 13, e0205881. [Google Scholar] [CrossRef]
- Li, Z.; Li, M.; He, J.; Zhao, X.; Chaimanee, V.; Huang, W.F.; Nie, H.; Zhao, Y.; Su, S. Differential physiological effects of neonicotinoid insecticides on honey bees: A comparison between Apis mellifera and Apis cerana. Pestic. Biochem. Physiol. 2017, 140, 1–8. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Nasr, H.M.; Rabea, E.I. Toxicity and biochemical changes in the honey bee Apis mellifera exposed to four insecticides under laboratory conditions. Apidologie 2015, 46, 177–193. [Google Scholar] [CrossRef]
- Laurino, D.; Porporato, M.; Patetta, A.; Manino, A. Toxicity of neonicotinoid insecticides to honey bees: Laboratory tests. Bull. Insectol. 2011, 64, 107–113. [Google Scholar]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Michael, R. Mechanism for the differential toxicity of neonicotinoid insecticides in the honey bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- BRC—Database of Insects and their Food Plants. Available online: http://dbif.brc.ac.uk/invertebratesresults.aspx?insectid=4742 (accessed on 6 October 2022).
- Miller, K.E.; Polaszek, A.; Evans, D.M. A dearth of data: Fitting parasitoids into ecological networks. Trends Parasitol. 2021, 37, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Kosko, B. Fuzzy cognitive maps. Int. J. Man-Mach. Stud. 1986, 24, 65–75. [Google Scholar] [CrossRef]
- Ramsey, D.S.L.; Norbury, G.L. Predicting the unexpected: Using a qualitative model of a New Zealand dryland ecosystem to anticipate pest management outcomes. Austral. Ecol. 2009, 34, 409–421. [Google Scholar] [CrossRef]
- Vasslides, J.M.; Jensen, O.P. Quantitative vs. semiquantitative ecosystem models: Comparing alternate representations of an estuarine ecosystem. J. Coast. Res. 2017, 78, 287–296. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process Engineering | Socio-Ecological Systems |
|---|---|
| (S1) Every final product is represented in the structure | (SE1) There should be at least one well-defined terminal ecosystem service in the ecosystem structure. |
| (S2) A material represented in the structure is a raw material if and only if it is not an output of any operating unit represented in the structure | (SE2) An ecosystem service represented in the structure is exogenous if and only if it is not an output of a functional unit defined in the ecosystem structure. |
| (S3) Every operating unit represented in the structure is defined in the synthesis problem | (SE3) Every ecosystem functional unit in the ecosystem structure is well defined. |
| (S4) Any operating unit represented in the structure has at least one path leading to a product | (SE4) Any ecosystem functional unit has at least one path leading to a terminal ecosystem service. |
| (S5) If a material belongs to the structure, it must be an input to or output from at least one operating unit represented in the structure | (SE5) If an ecosystem service belongs to the ecosystem structure, it must be an input to or output from at least one ecosystem functional unit represented in the structure. |
| Assumption: capital is available to pay for operating units and operating costs. | Assumption: energy is available to keep the ecosystem structure functioning |
| Goals: (1) meet production goal at (2) minimum cost for the structure and operation | Goals: (1) meet ecosystem services goal; and (2) minimize a cost metric (e.g., money, ecological footprint, energy, etc.) for the structure, management, functioning, and operations of the ecosystem services |
| Material/Unit | Description |
|---|---|
| Chemical_Dosing | Chemical control strategy against Leptinotarsa decemlineata (CPB) |
| Control | Overall pest control |
| CPB_adults | Adult CPB |
| CPB_eggs | Eggs of CPB |
| CPB_larvae | Larvae of CPB |
| CPB_pupae | Pupae of CPB |
| E_puttleri_Pop | Edovum puttleri population |
| E_puttleri_Rep | Reproductive capacity of E. puttleri |
| Eclosion | Eclosion of CPB pupae |
| Foliage | Foliage of the potato plant |
| Hatching | Hatching of CPB eggs |
| Health_of_E_puttleri | Overall health of E. puttleri |
| Health_of_L_decemlineata | Overall health of CPB |
| Health_of_O_dichrous | Overall health of Oplomus dichrous |
| Health_of_Potato | Overall health of the potato plant |
| Infestation | Infestation capacity of CPB on the potato plant |
| L_decemlineata_Est | Establishment of invading CPB |
| L_decemlineata_Pop | CPB population |
| L_decemlineata_Rep | Reproductive capacity of CPB on the host potato plant |
| Level_of_Infestation | Observable level of infestation by CPB on the host potato plant |
| O_dichrous_Pop | O. dichrous population |
| O_dichrous_Rep | Reproductive capacity of O. dichrous |
| Pesticide | Thiamethoxam |
| Potato_Rep | Reproduction of the potato plant |
| Pupation | Pupation of CPB larvae |
| Tuber | Yield of consumable potato tubers |
| Tuberization | Formation of tubers by the potato plant |
| Underground_Bud | Underground buds of the potato plant |
| Vegetative_Rep | Vegetative reproduction of the potato plant via tubers not for human consumption |
| Solution Structure | Represented Scenario | Tuber Production | Level of Infestation | Control |
|---|---|---|---|---|
| SS1 | Chemically controlled system, with two biological control agents eradicated as well | 0.816 | 0.23 | 1 |
| SS2 | Biologically controlled system with Edovum puttleri | 0.920 | 0.1 | 1 |
| SS3 | Biologically controlled system with Oplomus dichrous | 0.976 | 0.03 | 1 |
| SS4 | Insecticide applied prior to Leptinotarsa decemlineata (CPB) establishment, eradicating either biological control agent | 1 | N.A. | 1 |
| SS5 | Biologically controlled system with release of E. puttleri upon detection of eggs from invading CPB | 1 | N.A. | 1 |
| SS6 | Biologically controlled system with release of O. dichrous upon the occurrence of immature and mature progeny of invading CPB | 1 | N.A. | 1 |
| SS7 | System with uncontrolled CPB infestation | 0.231 | 0.961538 | N.A. |
| SS8 | Completely healthy system, without CPB | 1 | N.A. | N.A. |
| SS9 | System without potato, but with invading CPB and occurring natural enemies killed by insecticide | N.A. | N.A. | 1 |
| SS10 | Null | N.A. | N.A. | N.A. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almarinez, B.J.M.; Amalin, D.M.; Aviso, K.B.; Cabezas, H.; Lao, A.R.; Tan, R.R. Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae). Insects 2023, 14, 731. https://doi.org/10.3390/insects14090731
Almarinez BJM, Amalin DM, Aviso KB, Cabezas H, Lao AR, Tan RR. Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae). Insects. 2023; 14(9):731. https://doi.org/10.3390/insects14090731
Chicago/Turabian StyleAlmarinez, Billy Joel M., Divina M. Amalin, Kathleen B. Aviso, Heriberto Cabezas, Angelyn R. Lao, and Raymond R. Tan. 2023. "Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae)" Insects 14, no. 9: 731. https://doi.org/10.3390/insects14090731
APA StyleAlmarinez, B. J. M., Amalin, D. M., Aviso, K. B., Cabezas, H., Lao, A. R., & Tan, R. R. (2023). Network Modeling for Post-Entry Management of Invasive Pest Species in the Philippines: The Case of the Colorado Potato Beetle, Leptinotarsa decemlineata (Say, 1824) (Coleoptera: Chrysomelidae). Insects, 14(9), 731. https://doi.org/10.3390/insects14090731