
The Introduction and Establishment of Four Invasive Insect Species in Serbia



Abstract
Simple Summary
Abstract
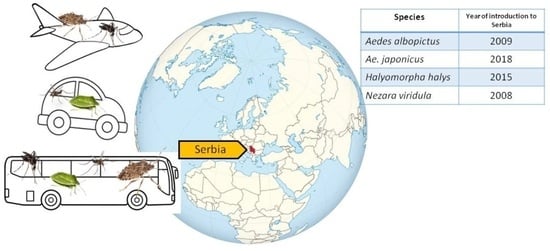
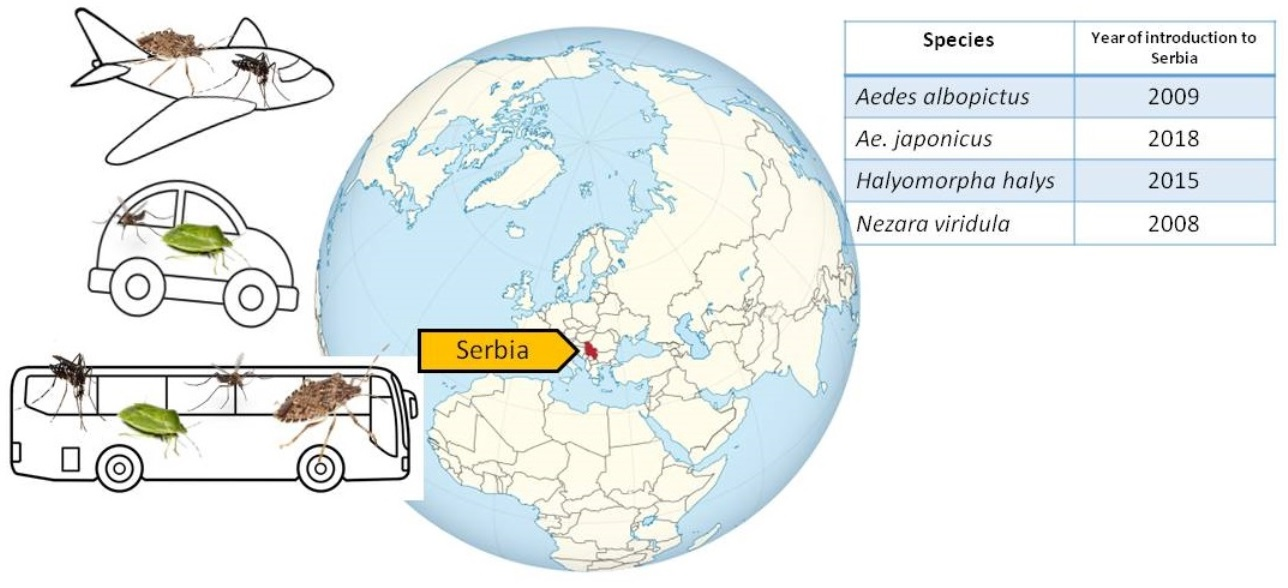
1. Introduction
2. The Asian Tiger Mosquito, Aedes albopictus Skuse (Diptera, Culicidae)
2.1. Distribution
2.2. Significance
2.3. First Record in Serbia
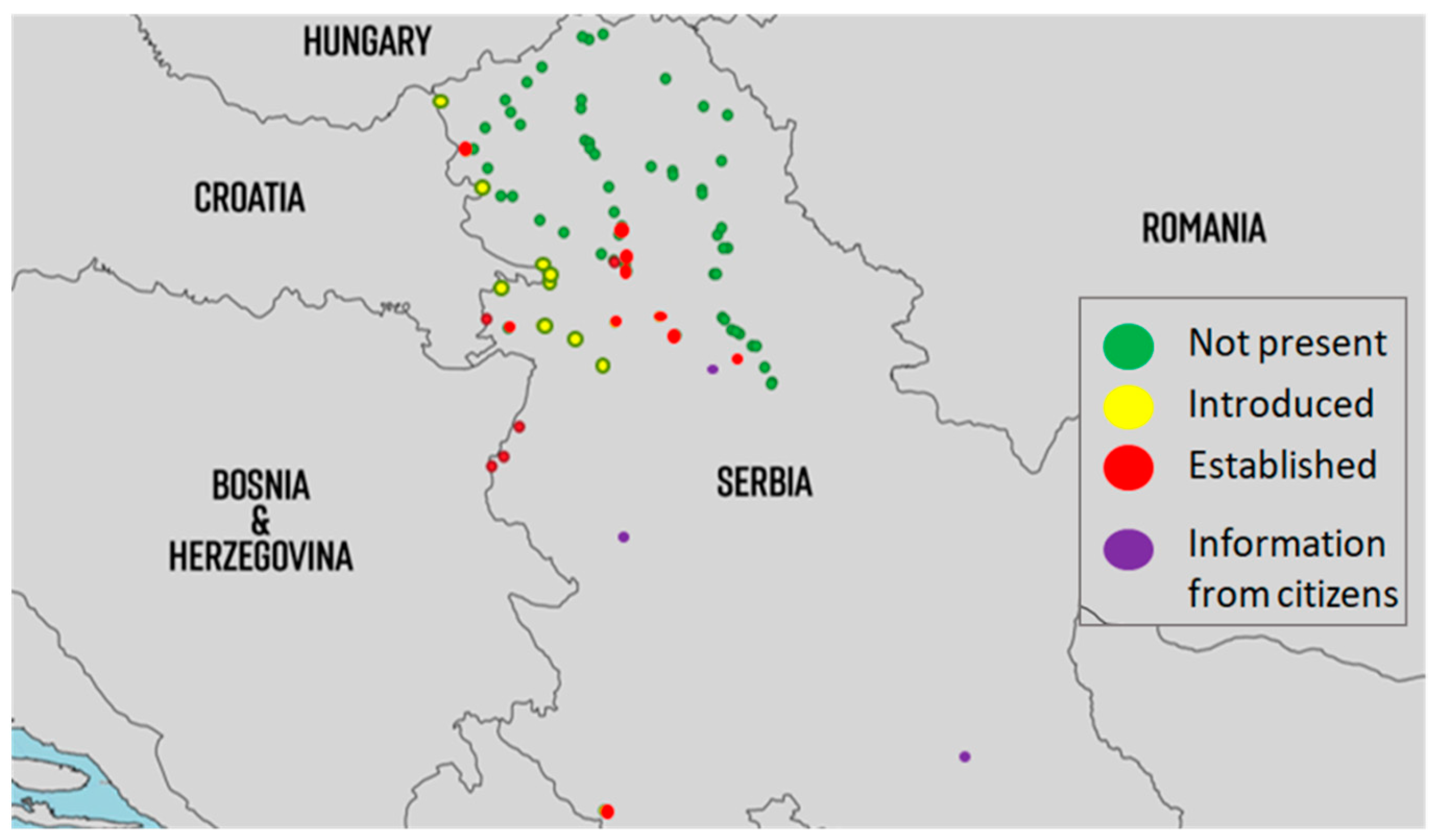
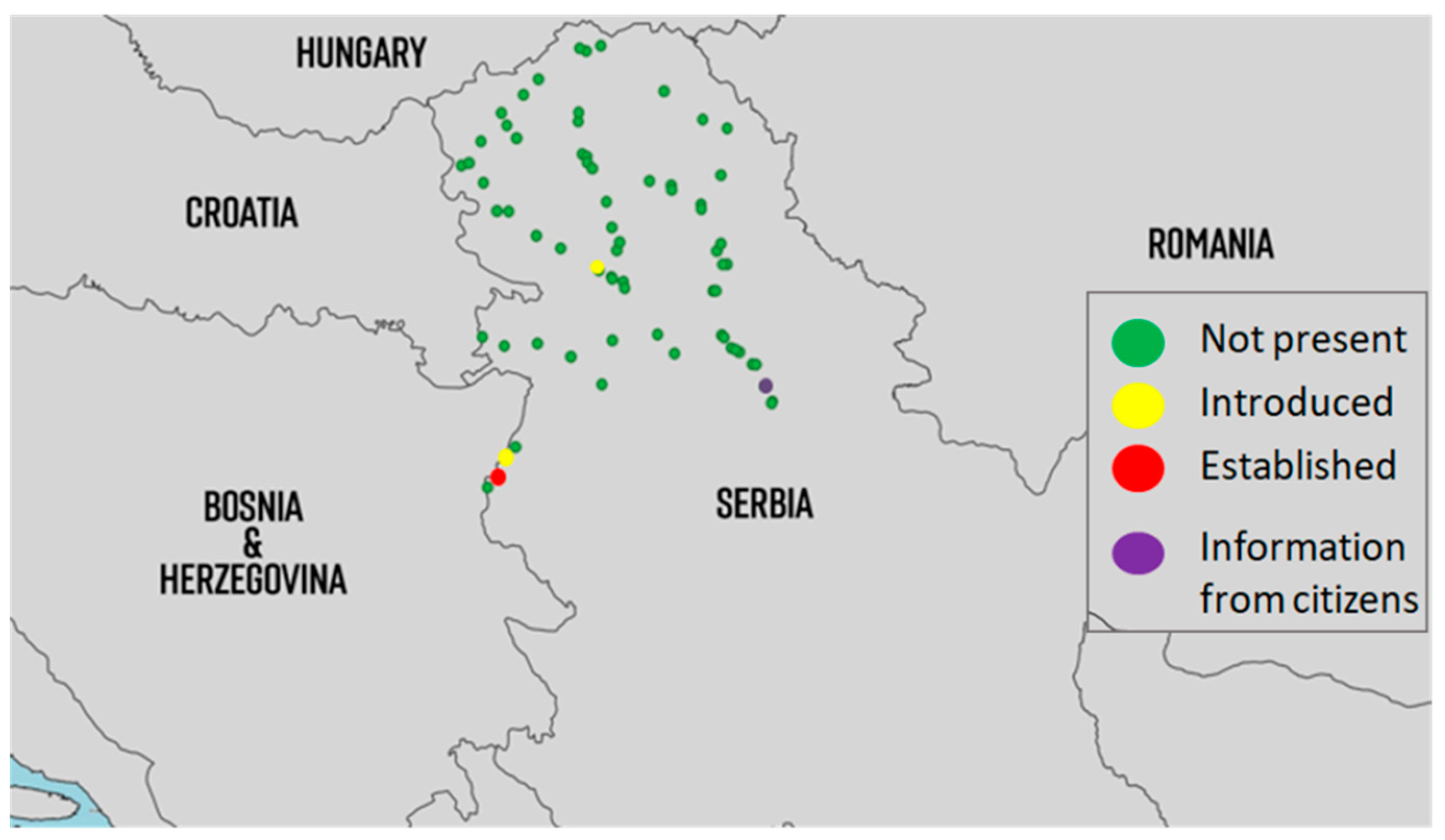
2.4. Monitoring
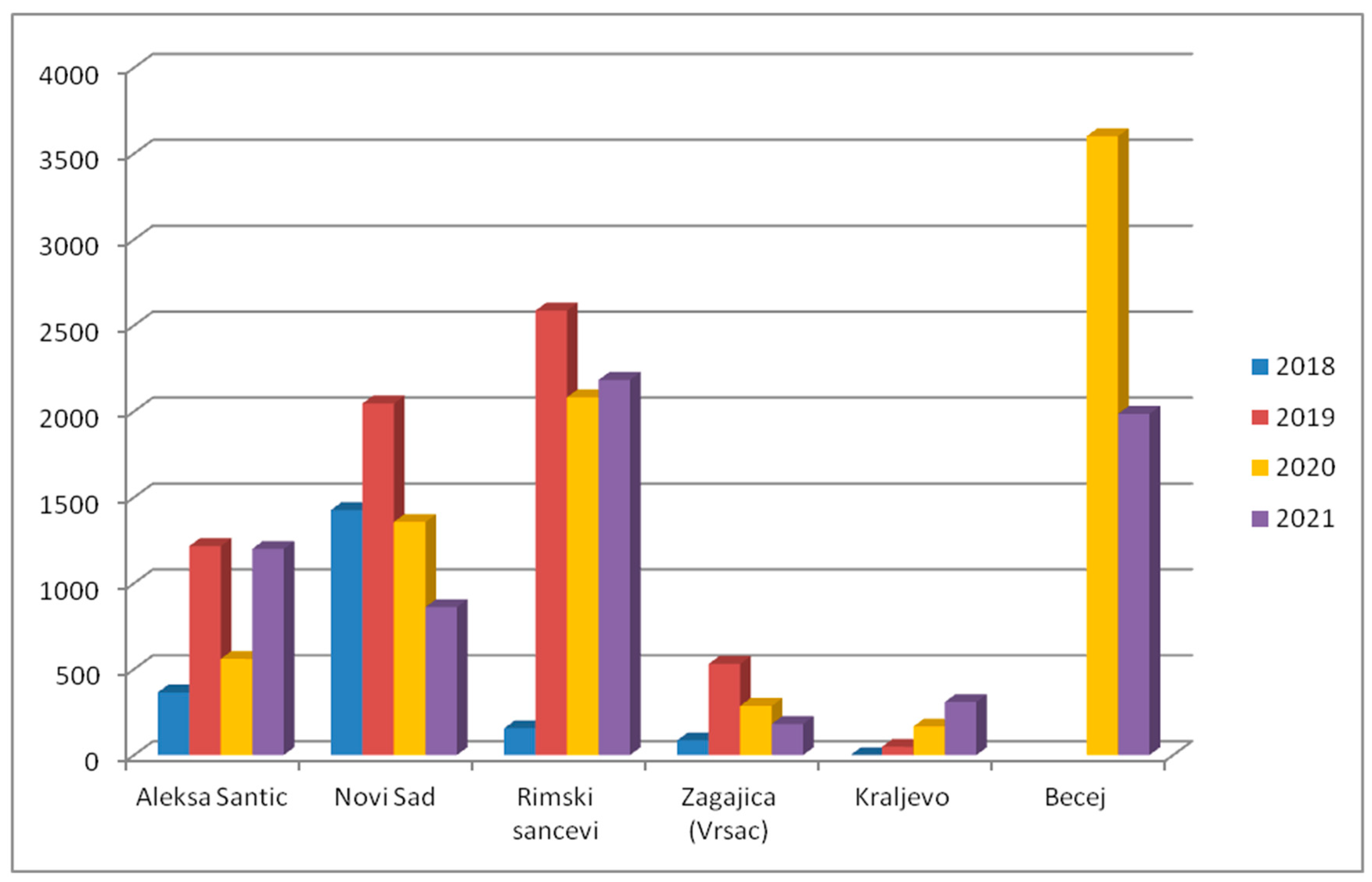
2.5. Distribution
2.6. Applied Control Measures
3. Japanese Bush Mosquito, Aedes japonicus (Theobald) (Diptera, Culicidae)
3.1. Distribution
3.2. Significance
3.3. First Record in Serbia
3.4. Monitoring
3.5. Distribution
3.6. Applied Control Measures
4. Brown Marmorated Stink Bug, Halyomorpha halys Stål (Hemiptera, Pentatomidae)
4.1. Distribution
4.2. Significance
4.3. First Record in Serbia
4.4. Monitoring
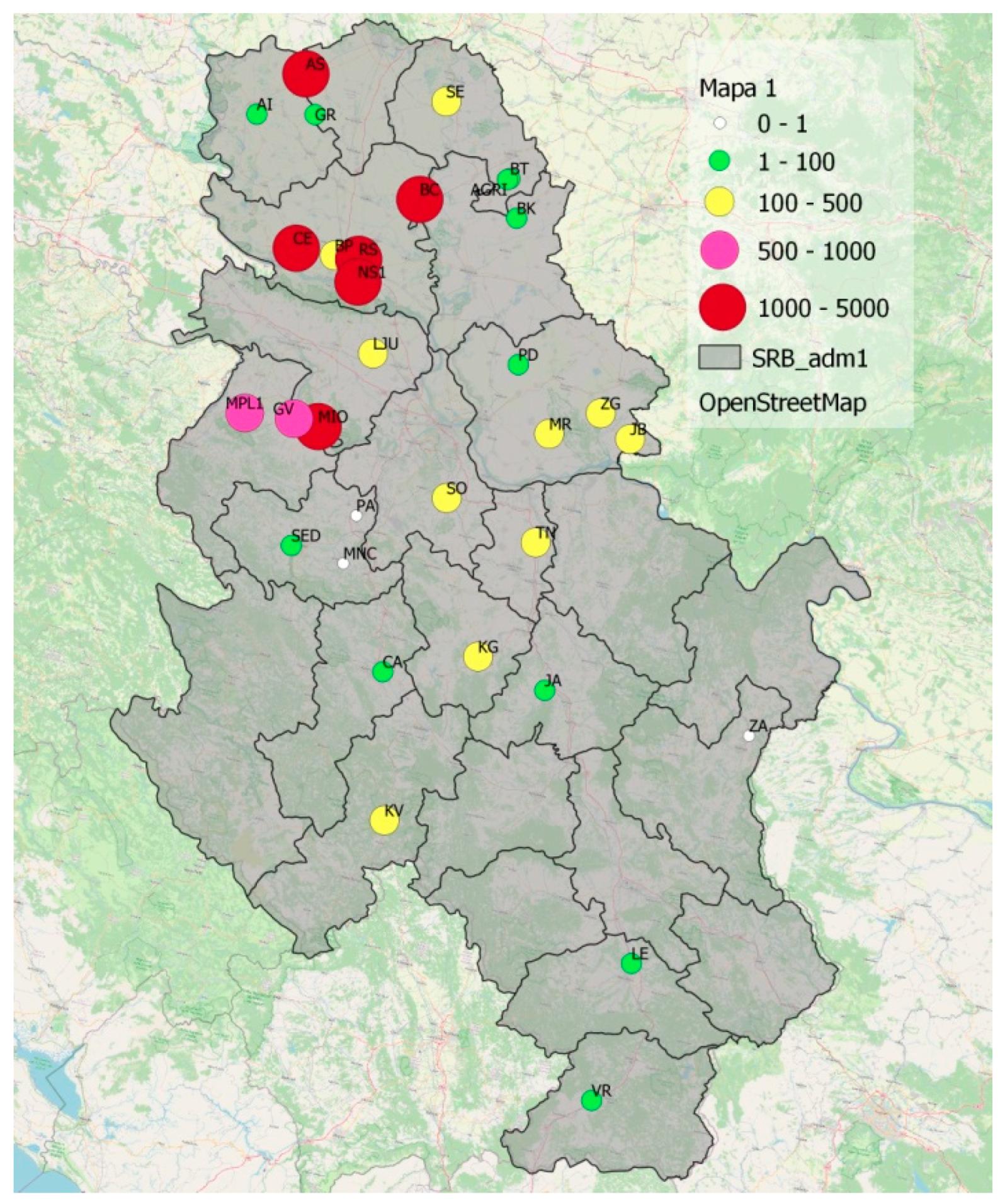
4.5. Distribution
4.6. Applied Control Measures
5. Southern Green Stink Bug, Nezara viridula (Linnaeus), (Hemiptera, Pentatomidae)
5.1. Distribution
5.2. Significance
5.3. First Record in Serbia
5.4. Monitoring
5.5. Distribution
5.6. Applied Control Measures
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Centre for Disease Prevention and Control, ECDC. The Climatic Suitability for Dengue Transmission in Continental Europe; ECDC: Stockholm, Sweden, 2012.
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Guidelines for the Surveillance of Invasive Mosquitoes in Europe; ECDC: Stockholm, Sweden, 2012. Available online: http://ecdc.europa.eu/en/publications/Publications/TER-Mosquitosurveillance-guidelines.pdf (accessed on 4 June 2023).
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souarès, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, southeastern France. Emerg. Infect. Dis. 2011, 17, 910. [Google Scholar] [CrossRef]
- Rocklöv, J.; Dubrow, R. Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar] [CrossRef]
- Caldwell, J.M.; LaBeaud, A.D.; Lambin, E.F.; Stewart-Ibarra, A.M.; Ndenga, B.A.; Mutuku, F.M.; Krystosik, A.R.; Ayala, E.B.; Anyamba, A.; Borbor-Cordova, M.J.; et al. Climate predicts geographic and temporal variation in mosquito-borne disease dynamics on two continents. Nat. Commun. 2021, 12, 1233. [Google Scholar] [CrossRef]
- Haye, T.; Wyniger, D.; Gariepy, T. Recent range expansion of brown marmorated stink bug in Europe. In Proceedings of the 8th International Conference on Urban Pests, Zurich, Switzerland, 20–23 July 2014; Executive Committee of the International Conference on Urban Pests, 2014; pp. 309–314. [Google Scholar]
- Konjević, A. First records of the brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Hemiptera: Pentatomidae) in Republic of North Macedonia. Acta Zool. Bulg. 2020, 72, 687–690. [Google Scholar]
- Adhami, J.; Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus Skuse (Diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 1998, 14, 340–343. [Google Scholar]
- Sabatini, A.; Raineri, V.; Trovato, G.; Coluzzi, M. Aedes albopictus in Italy and possible diffusion of the species into the Mediterranean area. Parassitologia 1990, 32, 301–304. [Google Scholar]
- Hawley, W.A.; Reiter, P.; Copeland, R.S.; Pumpuni, C.B.; Craig, G.B., Jr. Aedes albopictus in North America: Probable introduction in used tires from northern Asia. Science 1987, 236, 1114–1116. [Google Scholar] [CrossRef]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control Assoc. Suppl. 1988, 1, 1–39. [Google Scholar]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Invasive Species Specialist Group, ISSG. Global Invasive Species Database—Aedes albopictus 2009. Available online: http://www.issg.org/database/species/ecology.asp?si=109&fr=1&sts=sss&lang=EN (accessed on 16 February 2023).
- European Centre for Disease Prevention and Control, ECDC. Development of Aedes albopictus Risk Maps; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2009.
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’guinn, M.L.; Andreadis, T.G.; Blow, J.A. An update on the potential of North American mosquitoes (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef]
- Eritja, R.; Escosa, R.; Lucientes, J.; Marques, E.; Roiz, D.; Ruiz, S. Worldwide invasion of vector mosquitoes: Present European distribution and challenges for Spain. Biol. Invasions 2005, 7, 87–97. [Google Scholar] [CrossRef]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector-Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef]
- Schaffner, F.; Chouin, S.; Guilloteau, J. First record of Ochlerotatus (Finlaya) japonicus japonicus (Theobald, 1901) in metropolitan France. J. Am. Mosq. Control Assoc. 2003, 19, 1–5. [Google Scholar]
- Medlock, J.M.; Hansford, K.M.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entomol. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef]
- Petrović, V.; Turkulov, V.; Ilić, S.; Milošević, V.; Petrović, M.; Petrić, D.; Potkonjak, A. First report of imported case of dengue fever in Republic of Serbia. Travel Med. Infect. Dis. 2016, 1, 60–61. [Google Scholar] [CrossRef]
- Petric, D. Monitoring of Invasive Vector Mosquitoes and Vectorborne Diseases; Report to Administration for Environmental Protection; Novi Sad City, Serbia, 2009; pp. 1–9. [Google Scholar]
- Tanaka, K.; Mizusawa, K.; Saugstad, E.S. A revision of the Adult and Larval Mosquitoes of Japan (Including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae); Contributions of the American Entomological Institute: San Francisco, CA, USA, 1979; Volume 16, pp. 1–987. [Google Scholar]
- Versteirt, V.; Schaffner, F.; Garros, C.; Dekoninck, W.; Coosemans, M.; Van Bortel, W. Introduction and establishment of the exotic mosquito species Aedes japonicus japonicus (Diptera: Culicidae) in Belgium. J. Med. Entomol. 2009, 46, 1464–1467. [Google Scholar] [CrossRef]
- Schaffner, F.; Kaufmann, C.; Hegglin, D.; Mathis, A. The invasive mosquito Aedes japonicus in Central Europe. Med. Vet. Entomol. 2009, 23, 448–451. [Google Scholar] [CrossRef]
- Seidel, B.; Duh, D.; Nowotny, N.; Allerberger, F. First record of the mosquitoes Aedes (Ochlerotatus) japonicus japonicus (Theobald, 1901) in Austria and Slovenia 2011 and for Aedes (Stegomyia) albopictus (Skuse, 1895) in Austria. Entomol Zeitschr 2012, 122, 223–226. [Google Scholar]
- Ibáñez-Justicia, A.; Kampen, H.; Braks, M.; Schaffner, F.; Steeghs, M.; Werner, D.; Zielke, D.; Den Hartog, W.; Brooks, M.; Dik, M.; et al. First report of established population of Aedes japonicus japonicus (Theobald, 1901) (Diptera, Culicidae) in the Netherlands. J. Eur. Mosq. Control Assoc. 2014, 32, 9–13. [Google Scholar]
- Klobučar, A.; Lipovac, I.; Žagar, N.; Mitrović-Hamzić, S.; Tešić, V.; Vilibić-Čavlek, T.; Merdić, E. First record and spreading of the invasive mosquito Aedes japonicus japonicus (Theobald, 1901) in Croatia. Med. Vet. Entomol. 2019, 33, 171–176. [Google Scholar] [CrossRef]
- Seidel, B.; Montarsi, F.; Huemer, H.P.; Indra, A.; Capelli, G.; Allerberger, F.; Nowotny, N. First record of the Asian bush mosquito, Aedes japonicus japonicus, in Italy: Invasion from an established Austrian population. Parasites Vectors 2016, 9, 284. [Google Scholar] [CrossRef]
- Eritja, R.; Ruiz-Arrondo, I.; Delacour-Estrella, S.; Schaffner, F.; Álvarez-Chachero, J.; Bengoa, M.; Puig, M.Á.; Melero-Alcíbar, R.; Oltra, A.; Bartumeus, F. First detection of Aedes japonicus in Spain: An unexpected finding triggered by citizen science. Parasites Vectors 2019, 12, 53. [Google Scholar] [CrossRef]
- Schaffner, F.; Ries, C. First evidence and distribution of the invasive alien mosquito Aedes japonicus (Theobald, 1901) in Luxembourg. Bull. Soc. Nat. Luxemb. 2019, 121, 169–183. [Google Scholar]
- Kavran, M.; Lučić, D.; Ignjatović-Ćupina, A.; Zgomba, M.; Petrić, D. The first record of Aedes japonicus in Posavina region, Bosnia and Herzegovina. In Proceedings of the E-SOVE, European Society for Vector Ecology Conference, Palermo, Italy, 22–26 October 2018; pp. 22–26. [Google Scholar]
- Kavran, M.; Ignjatović Ćupina, A.; Zgomba, M.; Žunić, A.; Bogdanović, S.; Srdić, V.; Dondur, D.; Pudar, D.; Marinković, D.; Petrić, D. Invasive mosquito surveillance and the first record of Aedes japonicus in Serbia. In Proceedings of the IXth International EMCA Conference, La Rochelle, France, 11–14 March 2019; pp. 10–14. [Google Scholar]
- Horváth, C.; Cazan, C.D.; Mihalca, A.D. Emergence of the invasive Asian bush mosquito, Aedes (Finlaya) japonicus japonicus, in an urban area, Romania. Parasites Vectors 2021, 14, 1–7. [Google Scholar]
- Vojtíšek, J.; Janssen, N.; Šikutová, S.; Šebesta, O.; Kampen, H.; Rudolf, I. Emergence of the invasive Asian bush mosquito Aedes (Hulecoeteomyia) japonicus (Theobald, 1901) in the Czech Republic. Parasites Vectors 2022, 15, 250. [Google Scholar] [CrossRef]
- Becker, N.; Huber, K.; Pluskota, B.; Kaiser, A. Ochlerotatus japonicus japonicus–a newly established neozoan in Germany and a revised list of the German mosquito fauna. Eur. Mosq. Bull. 2011, 29, 102. [Google Scholar]
- Kaufman, M.G.; Stanuszek, W.W.; Brouhard, E.A.; Knepper, R.G.; Walker, E.D. Establishment of Aedes japonicus japonicus and its colonization of container habitats in Michigan. J. Med. Entomol. 2014, 49, 1307–1317. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Campbell, S.; Crans, W.J.; Mogi, M.; Miyagi, I.; Toma, T.; Bullians, M.; Andreadis, T.G.; Berry, R.L.; Pagac, B.; et al. Aedes (Finlaya) japonicus (Diptera: Culicidae), a newly recognized mosquito in the United States: Analyses of genetic variation in the United States and putative source populations. J. Med. Entomol. 2001, 38, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, T.G.; Anderson, J.F.; Munstermann, L.E.; Wolfe, R.J.; Florin, D.A. Discovery, distribution, and abundance of the newly introduced mosquito Ochlerotatus japonicus (Diptera: Culicidae) in Connecticut, USA. J. Med. Entomol. 2001, 38, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.L. Contributions to the Mosquito Fauna of Southeast Asia IV: Species of the Subgroup Chrysolineatus of Group D, Genus Aedes, Subgenus Finlaya Theobald; American Entomological Institute: Logan, UT, USA, 1968. [Google Scholar]
- Kamimura, K. The distribution and habit of medically important mosquitoes of Japan. Jpn. J. Sanit. Zool. 1968, 1. [Google Scholar]
- Sardelis, M.R.; Dohm, D.J.; Pagac, B.; Andre, R.G.; Turell, M.J. Experimental transmission of eastern equine encephalitis virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 480–484. [Google Scholar] [CrossRef]
- Sardelis, M.R.; Turell, M.J.; Andre, R.G. Laboratory transmission of La Crosse virus by Ochlerotatus j. japonicus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 635–639. [Google Scholar]
- Sardelis, M.R.; Turell, M.J.; Andre, R.G. Experimental transmission of St. Louis encephalitis virus by Ochlerotatus j. japonicus. J. Am. Mosq. Control Assoc. 2003, 19, 159–162. [Google Scholar]
- Sardelis, M.R.; Turell, M.J. Ochlerotatus j. japonicus in Frederick County, Maryland: Discovery, distribution, and vector competence for West Nile virus. J. Am. Mosq. Control Assoc.-Mosq. News 2001, 17, 137–141. [Google Scholar]
- Takashima, I.; Rosen, L. Horizontal and vertical transmission of Japanese encephalitis virus by Aedes japonicus (Diptera: Culicidae). J. Med. Entomol. 1989, 26, 454–458. [Google Scholar] [CrossRef]
- Schaffner, F.; Vazeille, M.; Kaufmann, C.; Failloux, A.B.; Mathis, A. Vector competence of Aedes japonicus for Chikungunya and dengue viruses. J. Eur. Mosq. Control Assoc. 2011, 29, 141–142. [Google Scholar]
- Turell, M.J.; Byrd, B.D.; Harrison, B.A. Potential for Populations of Aedes j. japonicus to Transmit Rift Valley Fever Virus in the USA1. J. Am. Mosq. Control Assoc. 2013, 29, 133–137. [Google Scholar]
- Josifov, M.; Kerzhner, I.M. Heteroptera Aus Korea. II. Aradidae, Berytidae, Lygaeidae, Pyrrhocoridae, Rhopalidae, Alydidae, Coreidae, Urostylidae, Acanthosomatidae, Sautelleridae, Pentatomidae, Cydnidae, Plataspidae. Fragm. Faun. 1978, 23, 137–196. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stål) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Arnold, K. Halyomorpha halys (Stål, 1855), eine für die europäische fauna neu nachgewiesene wanzenart (Insecta: Heteroptera: Pentatomidae: Cappaeini). Mitteilungen Thüringer Entomol. 2009, 16, 19. [Google Scholar]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stal (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitteilungen-Schweiz. Entomol. Ges. 2008, 81, 1. [Google Scholar]
- Heckmann, R. Erster nachweis von Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) für deutschland. Heteropteron 2012, 36, 17–18. [Google Scholar]
- Callot, H.; Brua, C. Halyomorpha halys (Stål, 1855), la Punaise diabolique, nouvelle espèce pour la faune de France (Heteroptera Pentatomidae). L’Entomologiste 2013, 69, 69–71. [Google Scholar]
- Maistrello, L.; Dioli, P.; Vaccari, G.; Nannini, R.; Bortolotti, P.; Caruso, S.; Costi, E.; Montermini, A.; Casoli, L.; Bariselli, M. Primi rinvenimenti in Italia della cimice esotica Halyomorpha halys, una nuova minaccia per la frutticoltura. ATTI Giornate Fitopatol. 2014, 1, 283–288. [Google Scholar]
- Vetek, G.; Papp, V.; Haltrich, A.; Rédei, D. First record of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae), in Hungary, with description of the genitalia of both sexes. Zootaxa 2014, 3780, 194–200. [Google Scholar] [CrossRef]
- Rabitsch, W.; Friebe, G.J. From the west and from the east? First records of Halyomorpha halys (Stål, 1855) (Hemiptera: Heteroptera: Pentatomidae) in Vorarlberg and Vienna, Austria. Beiträge Entomofaunist. 2015, 16, 115–139. [Google Scholar]
- Macavei, L.I.; Băețan, R.; Oltean, I.; Florian, T.; Varga, M.; Costi, E.; Maistrello, L. First detection of Halyomorpha halys Stål, a new invasive species with a high potential of damage on agricultural crops in Romania. Lucr. Ştiinţifice 2015, 58, 105–108. [Google Scholar]
- Milonas, P.G.; Partsinevelos, G.K. First report of brown marmorated stink bug Halyomorpha halys Stål (Hemiptera: Pentatomidae) in Greece. EPPO Bull. 2014, 44, 183–186. [Google Scholar] [CrossRef]
- Dioli, P.; Leo, P.; Maistrello, L. Prime segnalazioni in Spagna e in Sardegna della specie aliena Halyomorpha halys (Stål, 1855) e note sulla sua distribuzione in Europa (Hemiptera, Pentatomidae). Rev. Gaditana Entomol. 2016, 7, 539–548. [Google Scholar]
- Simov, N. The invasive brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) already in Bulgaria. Ecol. Montenegrina 2016, 9, 51–53. [Google Scholar] [CrossRef]
- Gapon, D.A. First records of the brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera, Pentatomidae) in Russia, Abkhazia, and Georgia. Entomol. Rev. 2016, 96, 1086–1088. [Google Scholar] [CrossRef]
- Mityushev, I.M. First record of Halymorpha halys detection in Russia. Zashchita Karantin Rasteniĭ 2016, 3, 48. (In Russian) [Google Scholar]
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive brown marmorated stink bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of invasion, range expansion, early stages of establishment, and first records of damage to local crops. Arthropod-Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Hemala, V.; Kment, P. First record of Halyomorpha halys and mass occurrence of Nezara viridula in Slovakia (Hemiptera: Heteroptera: Pentatomidae). Plant Prot. Sci. 2017, 53, 247–253. [Google Scholar] [CrossRef]
- Rot, M.; Devetak, M.; Carlevaris, B.; Žežlina, J.; Žežlina, I. First record of brown marmorated stink bug (Halyomorpha halys Stål, 1855)(Hemiptera: Pentatomidae) in Slovenia. Acta Entomol. Slov. 2018, 26, 5–12. [Google Scholar]
- Šapina, I.; Jelaska, L.Š. First report of invasive brown marmorated stink bug Halyomorpha halys (Stål, 1855) in Croatia. EPPO Bull. 2018, 48, 138–143. [Google Scholar] [CrossRef]
- Zovko, M.; Ostojić, I.; Jurković, D.; Karić, N. First report of the brown marmorated stink bug, Halyomorpha halys (Stål, 1855) in Bosnia and Herzegovina. Rad. Poljopr. Prehrambenog Fak. Univ. u Sarajev.\Work. Fac. Agric. Food Sci. Univ. Sarajevo 2019, 64, 68–78. [Google Scholar]
- EPPO. First reports of Halyomorpha halys in Belgium, Bulgaria and in Malta and Update for other European Countries. EPPO Reporting Service 06: Article 117. 2019. Available online: https://gd.eppo.int/reporting/article-6547 (accessed on 1 April 2023).
- Gariepy, T.D.; Haye, T.; Fraser, H.; Zhang, J. Occurrence, genetic diversity, and potential pathways of entry of Halyomorpha halys in newly invaded areas of Canada and Switzerland. J. Pest Sci. 2014, 87, 17–28. [Google Scholar] [CrossRef]
- Wiman, N.G.; Walton, V.M.; Shearer, P.W.; Rondon, S.I.; Lee, J.C. Factors affecting flight capacity of brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). J. Pest Sci. 2015, 88, 37–47. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y. Studies on the pear bug, Halyomorpha picus. Acta Agric. Bor.-Sin. 1988, 3, 96–101. [Google Scholar]
- Lee, H.; Chung, B.; Kim, T.; Kwon, J.; Song, W.; Rho, C. Damage of sweet persimmon fruit by the inoculation date and number of stink bugs, Riptortus clavatus, Halyomorpha halys and Plautia stali. Korean J. Appl. Entomol. 2009, 48, 485–491. [Google Scholar] [CrossRef]
- Funayama, K. Control effect on the brown-marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), by combined spraying of pyrethroid and neonicotinoid insecticides in apple orchards in northern Japan. Appl. Entomol. Zool. 2012, 47, 75–78. [Google Scholar] [CrossRef]
- Leskey, T. Emergence of Brown Marmorated Stink Bug, Halyomorpha halys (Stål), as a Serious Pest of Agriculture 2012. Available online: https://archive.epa.gov/pesticides/biopesticides/web/pdf/leskey-epa-nafta-workshop.pdf (accessed on 10 February 2022).
- Sauer, C. The Marbled tree bug occurs again on the Deutschschweizer Gemusebau. Die Marmorierte Baumwanze tritt neu im Deutschschweizer Gemüsebau auf.) Extension Gemüsebau, Forschungsanstalt Agroscope Changins-Wädenswil. Gemüsebau Info 2012, 28, 4–5. [Google Scholar]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Šeat, J. Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) a new invasive species in Serbia. Acta Entomol. Serbica 2015, 20, 167–171. [Google Scholar]
- Knight, K.M.; Gurr, G.M. Review of Nezara viridula (L.) management strategies and potential for IPM in field crops with emphasis on Australia. Crop Prot. 2007, 26, 1–10. [Google Scholar] [CrossRef]
- Freeman, P. A contribution to the study of the genus Nezara AMYOT and SERVILLE (Hemiptera, Pentatomidae). Trans. R. Entomol. Soc. Lond. 1940, 80, 351–374. [Google Scholar] [CrossRef]
- Yukawa, J.; Kiritani, K. Polymorphism in the southern green stink bug. Pac. Insects 1965, 7, 639–642. [Google Scholar]
- Ohno, K.; Alam, M.Z. Hereditary basis of adult color polymorphism in the southern green stink bug, Nezara viridula Linné (Heteroptera: Pentatomidae). Appl. Entomol. Zool. 1992, 27, 133–139. [Google Scholar] [CrossRef][Green Version]
- Todd, J.W. Ecology and behavior of Nezara viridula. Annu. Rev. Entomol. 1989, 34, 273–292. [Google Scholar] [CrossRef]
- Protić, L.J. New Heteroptera for the fauna of Serbia. Bull. Nat. Hist. Mus. 2011, 4, 119–125. [Google Scholar]
- Kereši, T.; Sekulić, R.; Protić, L.; Milovac, Ž. Occurrence of Nezara viridula L.(Heteroptera: Pentatomidae) in Serbia. Biljn. Lek./Plant Dr. 2012, 40, 296–304, (In Serbian, Summary in English). [Google Scholar]
- GISD-Global Invasive Species Database (2023). Available online: http://www.iucngisd.org/gisd/search.php (accessed on 5 July 2023).
- Leisnham, P.T.; Juliano, S.A. Impacts of climate, land use, and biological invasion on the ecology of immature Aedes mosquitoes: Implications for La Crosse emergence. Ecohealth 2012, 9, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Roiz, D.; Neteler, M.; Castellani, C.; Arnoldi, D.; Rizzoli, A. Climatic factors driving invasion of the tiger mosquito (Aedes albopictus) into new areas of Trentino, northern Italy. PLoS ONE 2011, 6, e14800. [Google Scholar] [CrossRef]
- Scaccini, D.; Falagiarda, M.; Tortorici, F.; Martinez-Sañudo, I.; Tirello, P.; Reyes-Domínguez, Y.; Gallmetzer, A.; Tavella, L.; Zandigiacomo, P.; Duso, C.; et al. An insight into the role of Trissolcus mitsukurii as biological control agent of Halyomorpha halys in Northeastern Italy. Insects 2020, 11, 306. [Google Scholar] [CrossRef]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the distribution of exotic egg parasitoids of Halyomorpha halys in Europe with a large-scale monitoring program. Insects 2021, 12, 316. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year of Introduction | Year of Establishment/Mass Occurrence |
|---|---|---|
| Aedes albopictus | 2009 | 2017 |
| Ae. japonicus | 2018 | 2022 |
| Halyomorpha halys | 2015 | 2017 |
| Nezara viridula | 2008 | 2010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kavran, M.; Konjević, A.; Petrić, D.; Ćupina, A.I. The Introduction and Establishment of Four Invasive Insect Species in Serbia. Insects 2023, 14, 728. https://doi.org/10.3390/insects14090728
Kavran M, Konjević A, Petrić D, Ćupina AI. The Introduction and Establishment of Four Invasive Insect Species in Serbia. Insects. 2023; 14(9):728. https://doi.org/10.3390/insects14090728
Chicago/Turabian StyleKavran, Mihaela, Aleksandra Konjević, Dušan Petrić, and Aleksandra Ignjatović Ćupina. 2023. "The Introduction and Establishment of Four Invasive Insect Species in Serbia" Insects 14, no. 9: 728. https://doi.org/10.3390/insects14090728
APA StyleKavran, M., Konjević, A., Petrić, D., & Ćupina, A. I. (2023). The Introduction and Establishment of Four Invasive Insect Species in Serbia. Insects, 14(9), 728. https://doi.org/10.3390/insects14090728