
Screening of Entomopathogenic Fungal Culture Extracts with Honeybee Nosemosis Inhibitory Activity

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honeybees and N. ceranae
2.2. Entomopathogenic Fungal Culture Extract
2.3. Purification of Nosema Spores
2.4. In Vitro Germination Assay
2.5. Safety Test of Fungal Culture Extract
2.6. Inhibitory Activity of Fungal Culture Extracts on Nosema Infection in Honeybees
2.7. Statistical Analysis
3. Results
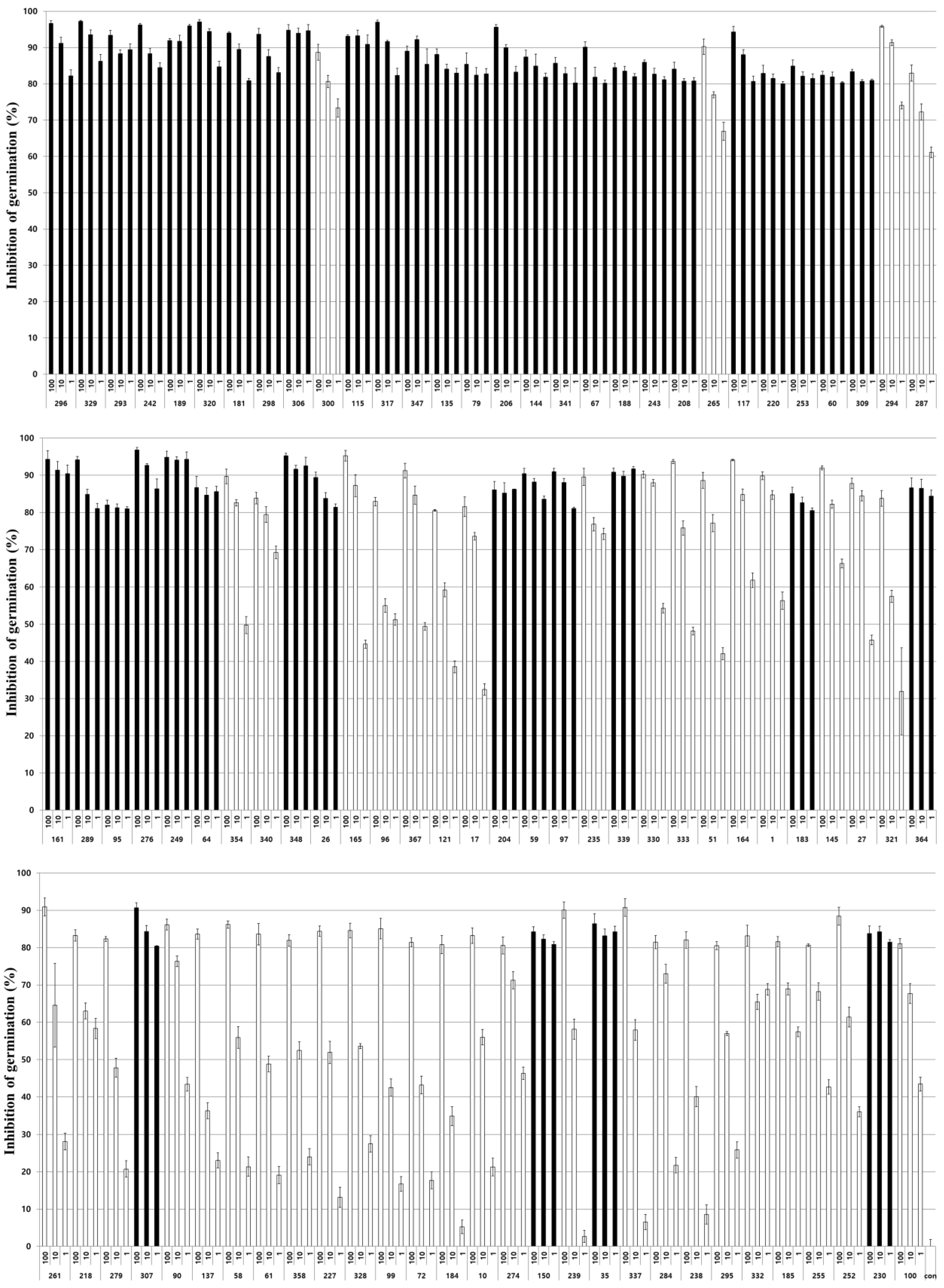
3.1. Inhibitory Activity of Entomopathogenic Fungal Culture Extracts on Nosema Germination
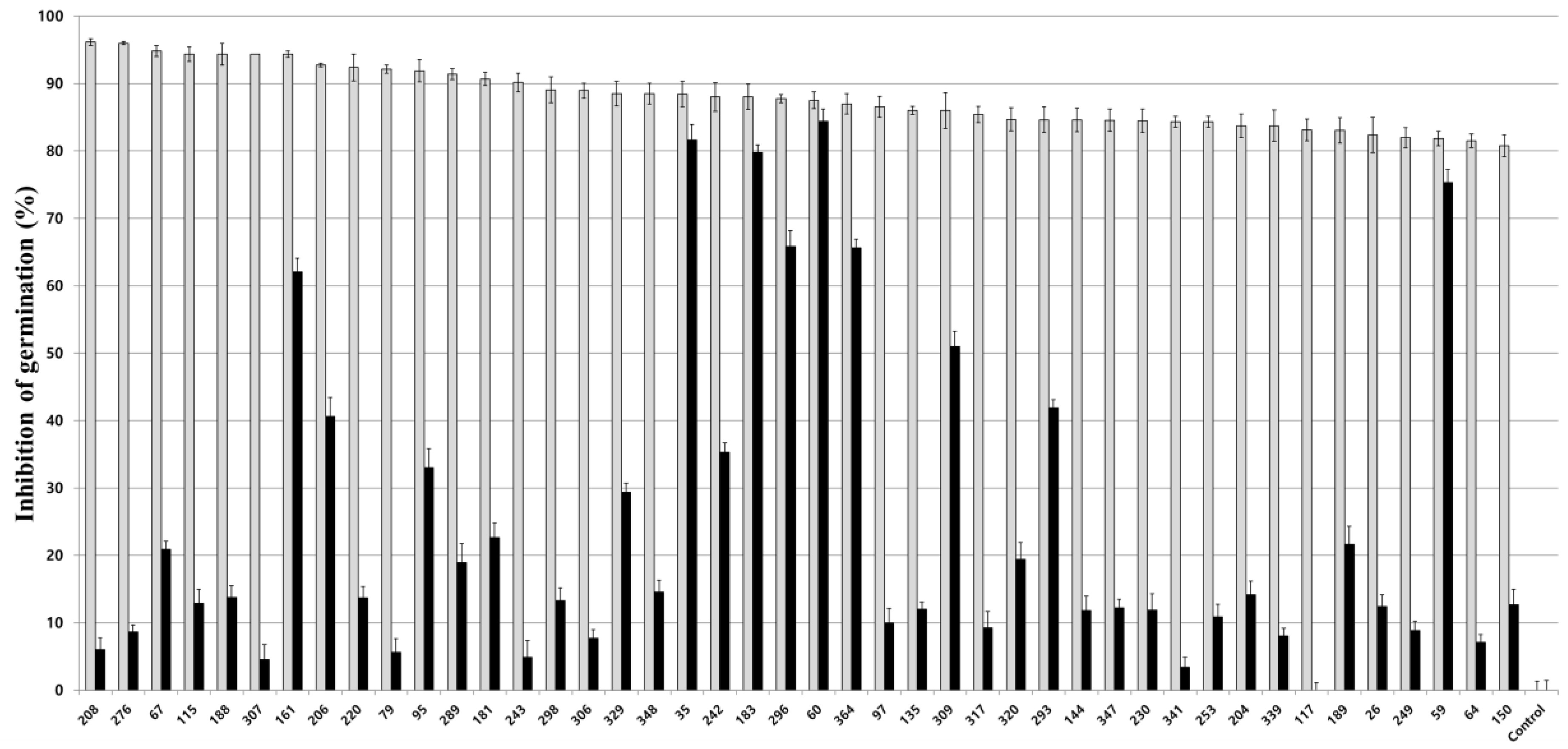
3.2. Inhibitory Activity of Fungal Culture Extracts on Nosema Spore Germination
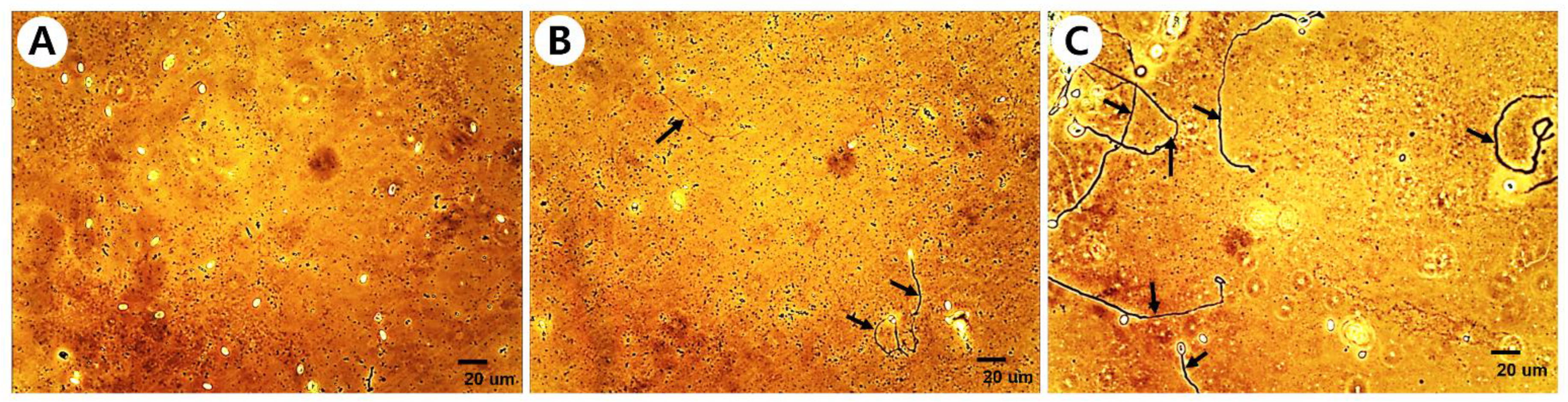
3.3. Mechanism of Inhibitory Activity of Fungal Culture Extracts on Nosema Spore Germination
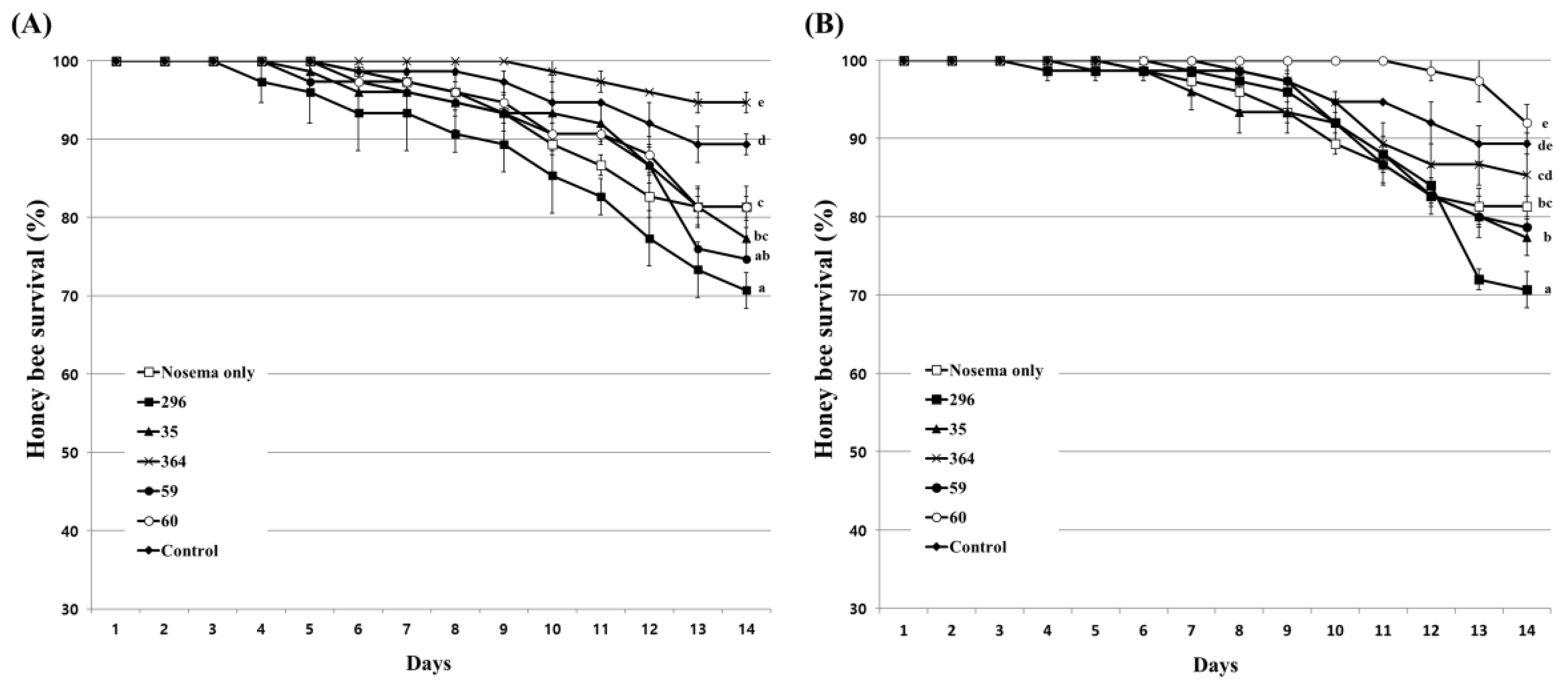
3.4. Influence of Fungal Culture Extracts on Honeybees
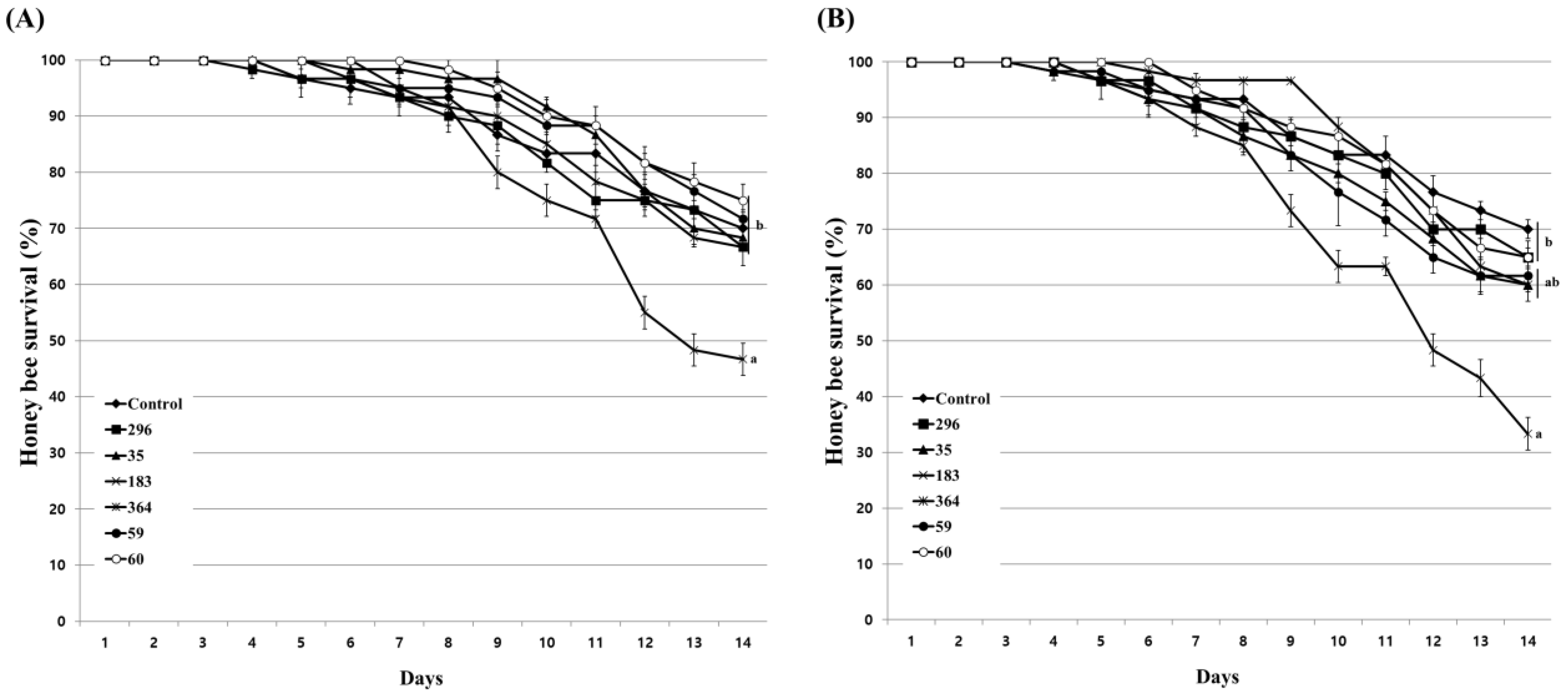
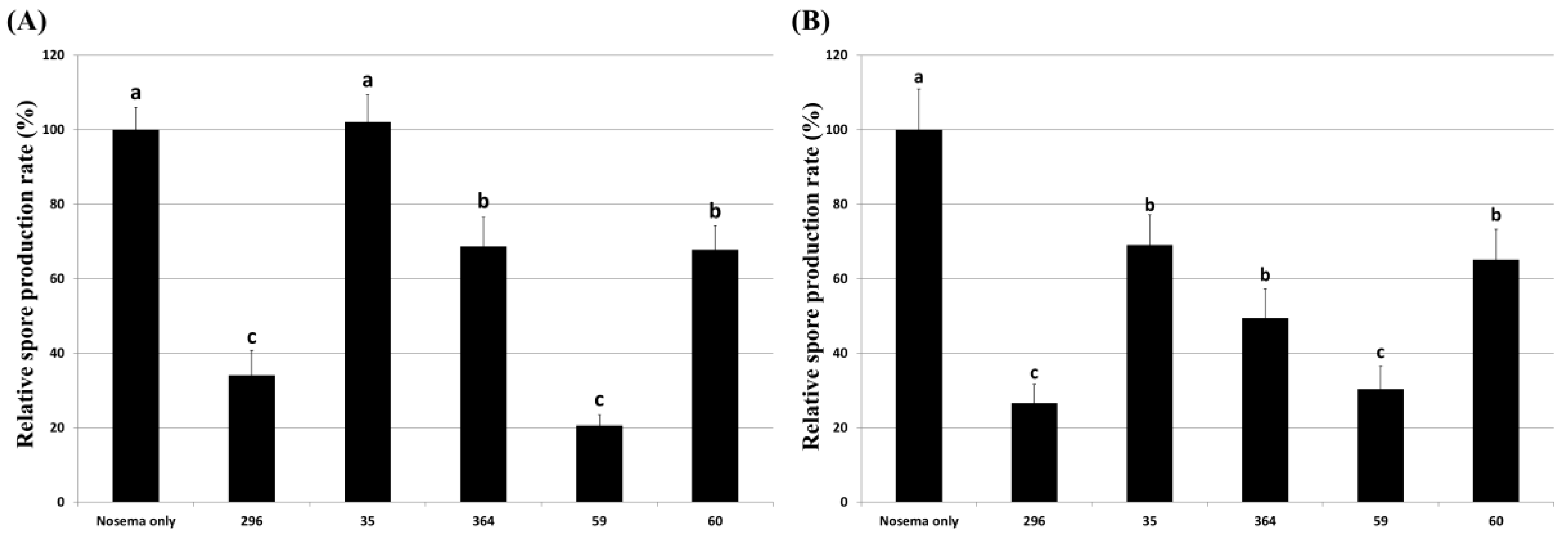
3.5. Inhibitory Effect of Culture Extract on Honeybee Nosemosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Breeze, T.D.; Bailey, A.P.; Balcombe, K.G.; Potts, S.G. Pollination services in the UK: How important are honeybees? Agric. Ecosyst. Environ. 2011, 142, 137–143. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Papa, G.; Maier, R.; Durazzo, A.; Lucarini, M.; Karabagias, I.K.; Plutino, M.; Bianchetto, E.; Aromolo, R.; Pignatti, G.; Ambrogio, A.; et al. The honey bee Apis mellifera: An insect at the interface between human and ecosystem health. Biology 2022, 11, 233. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. Honey and health: A review of recent clinical research. Pharmacogn. Res. 2017, 9, 121–127. [Google Scholar]
- Luo, X.; Dong, Y.; Gu, C.; Zhang, X.; Ma, H. Processing technologies for bee products: An overview of recent developments and perspectives. Front. Nutr. 2021, 8, 727181. [Google Scholar] [CrossRef] [PubMed]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.P.; Vanengelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors associated with honey bee colony losses: A mini-review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Stanimirovic, Z.; Glavinic, U.; Ristanic, M.; Aleksic, N.; Jovanovic, N.; Vejnovic, B.; Stevanovic, J. Looking for the causes of and solutions to the issue of honey bee colony losses. Acta Vet-Beograd 2019, 69, 1–31. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Del Nozal, M.J.; Mayo, R.; Bernal, J.L. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Vanengelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can Varroa explain part of the colony losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; van Engelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of Eukaryotes. J. Eukaryotic Microbiol. 2012, 59, 429–514. [Google Scholar] [CrossRef]
- Galajda, R.; Valenčáková, A.; Sučik, M.; Kandráčová, P. Nosema disease of European honey bees. J. Fungi 2021, 7, 714. [Google Scholar] [CrossRef]
- Fries, I. Nosema in European honey bees (Apis mellifera): An emerging threat to bee health. Insect Sci. 2010, 17, 418–424. [Google Scholar]
- Mayack, C.; Naug, D. Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 2010, 105, 20–27. [Google Scholar] [CrossRef]
- Goblirsch, M.J.; Spivak, M.; Kurtti, T.J. Nosema spp. infections in honey bees (Apis mellifera): Literature review and a survey of prevalence in North America. Insect Sci. 2013, 20, 1–22. [Google Scholar]
- Huang, W.F.; Solter, L.F.; Yau, P.M. Nosema ceranae causes asynchronous emergence of honey bee workers and reduces colony productivity. Apidologie 2018, 49, 11–22. [Google Scholar]
- Higes, M.; Martín-Hernández, R.; Meana, A. Nosema ceranae in European honey bees (Apis mellifera): An emerging infectious disease threatening apiaries and beekeeping. Insects 2020, 11, 886. [Google Scholar]
- Glavinic, U.; Stevanovic, J.; Ristanic, M.; Rajkovic, M.; Davitkov, D.; Lakic, N.; Stanimirovic, Z. Potential of fumagillin and Agaricus blazei mushroom extract to reduce Nosema ceranae in honey bees. Insects 2021, 12, 282. [Google Scholar] [CrossRef] [PubMed]
- Glavinic, U.; Rajkovic, M.; Vunduk, J.; Vejnovic, B.; Stevanovic, J.; Milenkovic, I.; Stanimirovic, Z. Effects of Agaricus bisporus mushroom extract on honey bees infected with Nosema ceranae. Insects 2021, 12, 915. [Google Scholar] [CrossRef] [PubMed]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chin, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Wide spread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Evans, J.D.; Simith, I.B.; Pettis, J.S. Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 2008, 97, 186–188. [Google Scholar] [CrossRef]
- Emsen, B.; Guzman-Novoa, E.; Hamiduzzaman, M.M.; Eccles, L.; Lacey, B.; Ruiz-Perez, R.A.; Nasr, M. Higher prevalence and levels of Nosema ceranae than Nosema apis infections in Canadian honey bee colonies. Parasitol. Res. 2016, 115, 175–181. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Vanengelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J.; Pettis, J.S. Entombed pollen: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef]
- Requier, F.; Garnery, L.; Kohl, P.L.; Njovu, H.K.; Pirk, C.W.W.; Crewe, R.M.; Steffan-Dewenter, I. The conservation of native honey bees is crucial. Trends Ecol. Evol. 2019, 34, 789–798. [Google Scholar] [CrossRef]
- Stanimirovic, Z.; Stevanovic, J.; Bajic, V.; Radovic, I. Evaluation of genotoxic effects of fumagillin (dicyclohexylamine) by citogenetic tests in vivo. Mutat. Res. 2006, 628, 1–10. [Google Scholar]
- Pajuelo, A.G.; Torres, C.; Bermejo, F.J.O. Colony losses: A double blind trial on the influence of supplementary protein nutrition and preventative treatment with fumagillin against Nosema ceranae. J. Apic. Res. 2008, 47, 84–86. [Google Scholar] [CrossRef]
- Williams, G.R.; Shutler, D.; Little, C.M.; Burgher-MacLellan, K.L.; Rogers, R.E.L. The microsporidian Nosema ceranae, the antibiotic Fumagilin-B®, and western honey bee (Apis mellifera) colony strength. Apidologie 2011, 42, 15–22. [Google Scholar] [CrossRef]
- Williams, G.R.; Sampson, M.A.; Shutler, D.; Rogers, R.E. Does fumagillin control the recently detected invasive parasite Nosema ceranae in western honey bees (Apis mellifera)? J. Apic. Res. 2018, 57, 714–721. [Google Scholar] [CrossRef]
- Sabaté, D.C.; Cruz, M.S.; Benítez-Ahrendts, M.R.; Audisio, M.C. Beneficial effects of Bacillus subtilis subsp. subtilis Mori2, a honey-associated strain, on honeybee colony performance. Probiot. Antimicrob. Proteins 2012, 4, 39–46. [Google Scholar] [CrossRef]
- Glavinic, U.; Blagojevic, J.; Ristanic, M.; Stevanovic, J.; Lakic, N.; Mirilovic, M.; Stanimirovic, Z. Use of thymol in Nosema ceranae control and health improvement of infected honey bees. Insects 2022, 13, 574. [Google Scholar] [CrossRef]
- Klassen, S.S.; VanBlyderveen, W.; Eccles, L.; Kelly, P.G.; Borges, D.; Goodwin, P.H.; Petukhova, T.; Wang, Q.; Guzman-Novoa, E. Nosema ceranae infections in honey bees (Apis mellifera) treated with pre/probiotics and impacts on colonies in the field. Vet. Sci. 2021, 8, 107. [Google Scholar] [CrossRef]
- Kunat-Budzyńska, M.; Budzyński, M.; Schulz, M.; Strachecka, A.; Gancarz, M.; Rusinek, R.; Ptaszyńska, A.A. Natural substances, probiotics, and synthetic agents in the treatment and prevention of honeybee nosemosis. Pathogens 2022, 11, 1269. [Google Scholar] [CrossRef]
- Lacey, L.A.; Frutos, R.; Kaya, H.K.; Vail, P. Insect pathogens as biological control agents: Do they have a future? Biol. Control 2001, 21, 230–248. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzon, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [PubMed]
- Isaka, M.; Kittakoop, P.; Kirtikara, K.; Hywel-Jones, N.L.; Thebtaranonth, Y. Bioactive substances from insect pathogenic fungi. Acc. Chem. Res. 2005, 38, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef]
- Shin, T.Y.; Bae, S.M.; Kim, D.J.; Yun, H.G.; Woo, S.D. Evaluation of virulence, tolerance to environmental factors and antimicrobial activities of entomopathogenic fungi against two-spotted spider mite, Tetranychus urticae. Mycoscience 2017, 58, 204–212. [Google Scholar] [CrossRef]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181–1206. [Google Scholar] [CrossRef]
- Shin, T.Y.; Lee, W.W.; Ko, S.H.; Choi, J.B.; Bae, S.M.; Choi, J.Y.; Lee, K.S.; Je, Y.H.; Jin, B.R.; Woo, S.D. Distribution and characterisation of entomopathogenic fungi from Korean soils. Biocontrol Sci. Technol. 2013, 23, 288–304. [Google Scholar] [CrossRef]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef]
- Kim, D.J.; Yun, H.G.; Kim, I.H.; Gwak, W.S.; Woo, S.D. Efficient method for the rapid purification of Nosema ceranae spores. Mycobiology 2017, 45, 204–208. [Google Scholar] [CrossRef]
- Chen, Y.P.; Evans, J.D.; Murphy, C.; Gutell, R.; Zuker, M.; Gundensen-Rindal, D.; Pettis, J.S. Morphological, molecular, and phylogenetic characterization of Nosema ceranae, a microsporidian parasite isolated from the European honey bee, Apis mellifera. J. Eukaryotic Microbiol. 2009, 56, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Shimanuki, H.; Knox, D.A. Diagnosis of Honey Bee Diseases; Agriculture Handbook No. AH-690; U.S. Department of Agriculture: Washington, DC, USA, 2000; p. 61.
- Olsen, P.E.; Rice, W.A.; Liu, T.P. In vitro germination of Nosema apis spores under conditions favorable for the generation and maintenance of sporoplasms. J. Invertebr. Pathol. 1986, 47, 65–73. [Google Scholar] [CrossRef]
- Graybill, J.R.; Burgess, D.S.; Hardin, T.C. Key issues concerning fungistatic versus fungicidal drugs. Eur. J. Clin. Microbiol. Infect. Dis. 1997, 16, 42–50. [Google Scholar] [CrossRef]
- Klarić, M.S.; Pepeljnjak, S. Bovericin: Kemizam, bioloski aspekti i rasirenost [Beauvericin: Chemical and biological aspects and occurrence]. Arh. Hig. Rada Toksikol. 2005, 56, 343–350. [Google Scholar] [PubMed]
- Liu, B.L.; Tzeng, Y.M. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front. Microbiol. 2021, 12, 705343. [Google Scholar] [CrossRef] [PubMed]
- Houard, J.; Aumelas, A.; Noël, T.; Pages, S.; Givaudan, A.; Fitton-Ouhabi, V.; Villain-Guillot, P.; Gualtieri, M. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J. Antibiot. 2013, 66, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.A.; Elkhalifa, A.E.O.; Siddiqui, A.J.; Patel, M.; Awadelkareem, A.M.; Snoussi, M.; Ashraf, M.S.; Adnan, M.; Hadi, S. Cordycepin for health and wellbeing: A potent bioactive metabolite of an entomopathogenic Cordyceps medicinal fungus and Its nutraceutical and therapeutic potential. Molecules 2020, 25, 2735. [Google Scholar] [CrossRef]
- Lacatena, F.; Marra, R.; Mazzei, P.; Piccolo, A.; Digilio, M.C.; Giorgini, M.; Woo, S.L.; Cavallo, P.; Lorito, M.; Vinale, F. Chlamyphilone, a novel Pochonia chlamydosporia metabolite with insecticidal activity. Molecules 2019, 24, 750. [Google Scholar] [CrossRef]
- Qin, F.; Li, Y.; Lin, R.; Zhang, X.; Mao, Z.; Ling, J.; Yang, Y.; Zhuang, X.; Du, S.; Cheng, X.; et al. Antibacterial radicicol analogues from Pochonia chlamydosporia and their biosynthetic gene cluster. J. Agric. Food Chem. 2019, 67, 7266–7273. [Google Scholar] [CrossRef]
- Dai, Z.B.; Wang, X.; Li, G.H. Secondary metabolites and their bioactivities produced by Paecilomyces. Molecules 2020, 25, 5077. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus | No. of Tested Isolates | No. of Isolates Showing the Inhibition of Spore Germination |
|---|---|---|
| All fungal isolates | 342 | 89 (26%) * |
| Acremonium strictum | 1 | 0 (0%) |
| Aspergillus lentulus. | 5 | 0 (0%) |
| Aspergillus versicolor | 3 | 1 (33.3%) |
| Beauveria bassiana | 110 | 48 (43.6%) |
| Beauveria brongniartii | 8 | 1 (12.5%) |
| Beauveria pseudobassiana | 8 | 1 (12.5%) |
| Bionectria ochroleuca | 7 | 0 (0%) |
| Clonostachys rosea | 1 | 0 (0%) |
| Cordyceps farinosa | 12 | 5 (41.7%) |
| Cordyceps fumosorosea | 6 | 1 (16.7%) |
| Cordyceps javanica | 22 | 5 (22.7%) |
| Fusarium oxysporum | 2 | 0 (0%) |
| Lecanicillium spp. | 8 | 1 (12.5%) |
| Metarhizium anisopliae | 64 | 15 (23.4%) |
| Metarhizium lepidiotae | 1 | 0 (0%) |
| Metarhizium pemphigus | 16 | 3 (18.8%) |
| Mucoromycotina spp. | 1 | 0 (0%) |
| Myrothecium spp. | 5 | 0 (0%) |
| Paecilomyces lilacinus | 7 | 1 (14.3%) |
| Paecilomyces marquandii | 5 | 1 (20%) |
| Paraconiothyrium sporulosum | 2 | 1 (50%) |
| Phialocephala spp. | 1 | 0 (0%) |
| Pochonia bulbillosa | 16 | 3 (18.8%) |
| Pochonia rubescens | 1 | 0 (0%) |
| Simplicillium aogashimaense | 1 | 1 (100%) |
| Simplicillium sp. | 2 | 0 (0%) |
| Tolypocladium album | 23 | 1 (4.3%) |
| Tolypocladium cylindrosporum | 3 | 0 (0%) |
| Verticillium insectorum | 1 | 0 (0%) |
| Fungus | No. of Isolate (%) |
|---|---|
| Aspergillus versicolor | 1 (2.3) * |
| Beauveria bassiana | 24 (54.5) |
| Beauveria brongniartii | 1 (2.3) |
| Cordyceps farinosa | 4 (9.1) |
| Cordyceps fumosorosea | 1 (2.3) |
| Cordyceps javanica | 1 (2.3) |
| Lecanicillium spp. | 1 (2.3) |
| Metarhizium anisopliae | 6 (13.6) |
| Paecilomyces lilacinus | 1 (2.3) |
| Paecilomyces marquandii | 1 (2.3) |
| Pochonia bulbillosa | 2 (4.5) |
| Tolypocladium album | 1 (2.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-J.; Woo, R.-M.; Kim, K.-S.; Woo, S.-D. Screening of Entomopathogenic Fungal Culture Extracts with Honeybee Nosemosis Inhibitory Activity. Insects 2023, 14, 538. https://doi.org/10.3390/insects14060538
Kim D-J, Woo R-M, Kim K-S, Woo S-D. Screening of Entomopathogenic Fungal Culture Extracts with Honeybee Nosemosis Inhibitory Activity. Insects. 2023; 14(6):538. https://doi.org/10.3390/insects14060538
Chicago/Turabian StyleKim, Dong-Jun, Ra-Mi Woo, Kyu-Seek Kim, and Soo-Dong Woo. 2023. "Screening of Entomopathogenic Fungal Culture Extracts with Honeybee Nosemosis Inhibitory Activity" Insects 14, no. 6: 538. https://doi.org/10.3390/insects14060538
APA StyleKim, D.-J., Woo, R.-M., Kim, K.-S., & Woo, S.-D. (2023). Screening of Entomopathogenic Fungal Culture Extracts with Honeybee Nosemosis Inhibitory Activity. Insects, 14(6), 538. https://doi.org/10.3390/insects14060538