Simple Summary
Planting a diverse array of flowers in crop field settings can support insect crop pollinators, as many pollinating insects feed on floral pollen and nectar as adults. Although adult pollinating flies that feed on floral resources will also be supported by flower plantings, fly larvae rarely feed on floral nectar and pollen. Here, we deployed pools filled with habitats (decaying plant materials, soil, water) that pollinating fly larvae are known to feed on in seed carrot fields in an attempt to attract flies to lay eggs. We found many fly eggs and larvae within the pools after 12 to 21 days. More eggs were laid on decaying plant stems and carrot roots, compared to other locations (e.g., on decaying carrot flowers, leaves, etc.) within the pools. The habitat pools we trialed within the seed carrot fields could be a quick and easy way to support the reproduction of beneficial fly pollinators. These results can be used to support future studies to examine the effect of habitat pools in crop fields on the number of flies that visit crop flowers.
Abstract
The addition of floral resources is a common intervention to support the adult life stages of key crop pollinators. Fly (Diptera) crop pollinators, however, typically do not require floral resources in their immature life stages and are likely not supported by this management intervention. Here, we deployed portable pools filled with habitat (decaying plant materials, soil, water) in seed carrot agroecosystems with the intention of providing reproduction sites for beneficial syrphid (tribe Eristalini) fly pollinators. Within 12 to 21 days after the pools were deployed, we found that the habitat pools supported the oviposition and larval development of two species of eristaline syrphid flies, Eristalis tenax (Linnaeus, 1758) and Eristalinus punctulatus (Macquart, 1847). Each habitat pool contained an average (±S.E.) of 547 ± 117 eristaline fly eggs and 50 ± 17 eristaline fly larvae. Additionally, we found significantly more eggs were laid on decaying plant stems and carrot roots compared to other locations within the pool habitat (e.g., on decaying carrot umbels, leaves, etc.). These results suggest that deploying habitat pools in agroecosystems can be a successful management intervention that rapidly facilitates fly pollinator reproduction. This method can be used to support future studies to determine if the addition of habitat resources on intensively cultivated farms increases flower visitation and crop pollination success by flies.
1. Introduction
The abundance and diversity of insects that provide pollination services within agroecosystems often depend on suitable habitat options for the insects to complete their life cycles [1,2]. These habitats (e.g., remnant vegetation, semi-natural landscape features) are typically not within the crop system itself, but nearby, and provide food, reproduction sites, overwintering resources, and shelter from agricultural management practices such as tilling, harvesting, and pesticide application [3,4,5,6,7]. When non-crop habitat is maintained or restored near intensely managed fields, beneficial insect abundance and diversity generally increase [4,8,9,10,11,12]. Even small patches (e.g., tens of square meters or less) of non-crop habitat can enhance beneficial insect biodiversity in cropping systems [3,4], and result in native species spillover into agroecosystems [4,13,14]. However, most pollinator-friendly habitat enhancements focus on floral interventions, such as floral strips and hedgerows [15,16,17,18], to attract adult, wild pollinators, primarily bees. Few studies have focused specifically on interventions to support the habitat needs for non-bee taxa (but see [2,19,20] for exceptions) and their non-floral resource needs (see [21] for an example of bee non-floral resource needs).
While bees are highly dependent on flowers to obtain nutrition for both adults and larvae, non-bee pollinator taxa, such as flies (Diptera), typically do not require floral resources in their larval stages [22]. For example, the larvae of eristaline syrphid flies, which are easily distinguishable due to the siphon-like ‘tail’ they use to breathe in poorly oxygenated habitats, live in wet substrates commonly found in agricultural environments, including decaying plant materials and manure [23,24,25]. Adult eristaline syrphid flies, like the cosmopolitan species Eristalis tenax (Linnaeus, 1758), have been shown to effectively pollinate crops as they morphologically resemble honeybees (Apis mellifera Linnaeus 1758) in size and body hairiness [26,27,28], despite lacking specialized pollen-collecting structures (e.g.,: scopa, corbicula). In fact, E. tenax is already a non-bee pollinator alternative in New Zealand, where the fly is an effective pollinator of multiple crops including seed carrot [29].
Seed carrot is an ideal model crop to study a potential non-bee pollinator since the crop often pollination limited despite attracting high numbers of other insect visitors [30,31]. Honeybees generally find seed carrot flowers unattractive, as the nectar composition is high in ferulic acid, an insect-feeding deterrent, and low in caffeic acid, a bee attractor [32]. As some species of eristaline flies have been shown to be as effective as honey bees at pollinating seed carrot [30,33], we hypothesized that building up populations of these beneficial non-bee pollinators could increase free ecosystem service delivery within seed carrot agroecosystems [29]. Therefore, this study was based in the Riverina region of New South Wales (NSW), Australia (AU), where seed carrot growers plan for the crop to bloom late austral spring and summer (November to December), when almost no other crops are concurrently blooming to best facilitate honeybee pollination services.
In this study, we trialed the deployment of small, portable pools filled with non-floral habitats to support the reproduction of eristaline syrphid flies in seed carrot agroecosystems. Although the life cycle of eristaline syrphid flies is generally well known [34,35,36], to our knowledge there are no studies that address whether eristaline flies have oviposition preferences within the habitat they utilize to lay eggs in natural field conditions. We, therefore, tested two habitats (soil with decaying carrot plants in water or decaying carrot plants in water only) to determine if existing adult eristaline syrphid flies would utilize the habitat pools as oviposition sites and evaluated where the flies oviposited within the habitat provided. We addressed the following research questions:
- Will eristaline syrphid flies use provisioned habitat pools as oviposition sites in a commercial field setting otherwise unsuitable for larval development?
- Which of the two habitats resulted in the greatest number of eggs and larvae?
- What were the specific features within the habitat pools that resulted in the greatest oviposition?
2. Materials and Methods
2.1. Study Sites
Seven study fields, at a minimum of 315 m apart, of seed carrot monocultures in the Riverina region of NSW, AU, were chosen as sites in four locations comprising three commercial farms and one private farm managed by South Pacific Seeds (Figure 1). The seed carrot plots at the commercial farms varied from 5 to 14 hectares, while the private farm grew commercial-grade seed carrot in small (<500 m) trial rows. Other plant resources flowering nearby included onion (Allium cepa L.) at sites six and seven and small patches of native flowering trees, shrubs, and household gardens near sites one, four, and five as these sites were situated near residential areas. No other crops were observed flowering nearby.
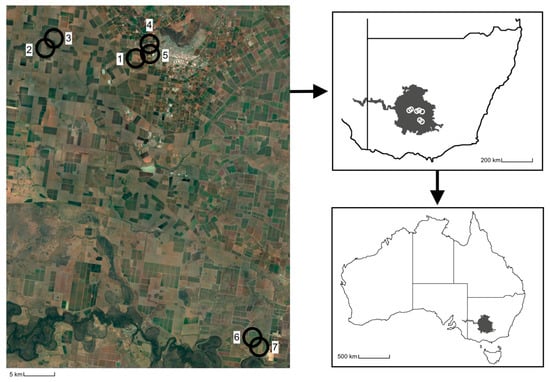
Figure 1.
Location of the seed carrot study sites (1–7) where the habitat pools were deployed within the Riverina region of New South Wales, Australia.
2.2. Deploying the Habitat Pools
In preliminary experiments at site one, we placed horse manure and wet, decaying carrot plants within a hybrid seed carrot plot to assess which substrates should be trialed as non-floral fly habitat. The substrates were observed frequently until we observed golden native droneflies, Eristalinus punctulatus (Macquart, 1847), oviposit within the wet, decaying carrot plants. As eristaline fly larvae are commonly reared in slurries of manure in laboratory conditions [24,36], this suggests that the larvae are more suited for semi-liquid environments. We did not trial manure at the field sites as golden native droneflies were not observed to oviposit within the manure and some landholders did not want manure placed on their properties; therefore, we chose to trial decaying carrot plants in water as reproduction sites for eristaline flies, with the presence or absence of farm soil. Thus, we hypothesized that more larvae would be found in the semi-liquid decaying carrot plant pool with soil, compared to the treatment with decaying plants and water only.
Pools were deployed during peak bloom (50% flowering) of seed carrot (15 November to 9 December 2021), when adult eristaline flies are most likely to visit seed carrot flowers. Two polypropylene pools (945 mm × 210 mm × 1100 mm each) were placed side by side in a paired experimental design at each site to trial two habitats as eristaline fly oviposition sites (n = 7 per treatment, 14 pools in total). The first habitat consisted of soil, discarded carrot plants and water, while the second habitat consisted of discarded carrot plants and water only. Soil from the farm site was placed within the first pool until the base of the pool was covered, while the second pool contained two bricks to anchor the pool from strong gusts of wind that frequently blow within the region. Three fully grown (150 cm foliage height) male carrot plants were then taken from the site and placed in each pool which was then filled with the same water used to water the seed carrot crop (Figure 2a). We did not include a treatment without water, since eristaline flies cannot survive in habitats devoid of water [35,36]. Likewise, sufficient solid food must be present within the water for eristaline larvae to complete development [30], so we did not include a treatment of water-only pools. Instead, we conducted preliminary surveys searching for the immature stages of eristaline flies within field sites before pools were deployed, to confirm that no eggs and larvae had been laid in dry soil, dry plant material, or within crop rows. As no eggs or larvae were found in the preliminary field surveys, we excluded them from analyses.

Figure 2.
Experimental design of the habitat pools deployed to attract eristaline flies: (a) a habitat pool yet to be filled with water within a seed carrot field; (b) an adult, female Eristalis tenax (Linnaeus, 1758) fly within a deployed habitat pool. Arrowheads are pointing to the habitat pool and adult eristaline fly for clarity.
After filling the pools with water, we left them undisturbed to allow the carrot plants within the pools to decay and for eristaline adults to locate the pools (Figure 2b). Due to unprecedented rain events at the time the pools were left undisturbed, site accessibility varied between farms; therefore, the pools were deployed for 12 to 21 days depending on field site accessibility.
2.3. Surveying the Immature Life Stages of Eristaline Flies
Once all farm sites were accessible on the same day, we conducted surveys in each pool to count eristaline syrphid fly egg clutches, a group of eggs laid together in a single oviposition attempt (Figure 3a), individual eggs, and larvae (Figure 3b). All egg clutches and individual eggs were counted on 9 December 2021 (starting at 06:00 and ending at 18:30) and were removed from the pools, so the eggs did not hatch before the larvae were counted. Additionally, we recorded the location of where the eggs were oviposited in the pools (Figure S1). All egg clutches and eggs that were displaced from their original positions in the pools (e.g., due to moving substrates) were counted but not included in statistical analyses.

Figure 3.
Immature stages of eristaline syrphid flies: (a) a clutch of eggs oviposited on a decaying carrot stem; (b) larvae found within a deployed habitat pool. Arrowheads are pointing to the immature stages of eristaline syrphid flies for clarity.
The day after the eggs were counted and collected, we returned to the pools and counted the eristaline fly larvae over a two-day period (with each pool counted only once for each immature life stage). To count the larvae, all plant material in the pools was thoroughly checked for individuals, and then removed from the pools. Next, we dislodged any larvae within the soil at the bottom of the soil treatment pools, by mixing the water with the soil sediment using a hand-held sieve. We then sifted the soil and water sediment through the sieve five times to determine the total number of larvae in the pools. When larvae were caught in the sieve, they were removed from the pools to avoid duplicate larval counts. For consistency, this procedure was also applied to the carrot and water treatment pools. We did not record where the larvae were found in the pools since we displaced all larvae when mixing the water with the sediment.
2.4. Rearing Eristaline Flies from Pools
Both eggs and larvae of various growth stages collected from each pool were reared to adulthood in controlled conditions on decaying carrot plants (from inside the pools) or a mixture of decaying carrot plants and sterilized horse manure to confirm species identities. Horse manure was mixed into the habitat as previous studies have successfully reared eristaline syrphid flies from manure [24,37]. Since eristaline syrphid flies have similar morphology in eggs and larvae and are therefore difficult to identify at these stages [23,37,38,39]; we waited until adults emerged to distinguish species identities using taxonomic keys [23,25].
2.5. Statistical Analyses
Statistical analyses were performed using R version 4.1.2. We created generalized linear mixed-effects models (GLMMs) using the MASS package to assess whether the number of eggs and larvae within pools differed based on treatment (two categories) or location (eight categories) [40,41]. To handle overdispersion in the collected count data, all GLMM models were fit to a negative binomial distribution [42]. Additionally, as the number of days the pools were left out in the sites to decay was not standardized as intended due to unprecedented weather conditions, we included the fixed effect ‘Day’ (continuous: 5 discrete values) in all models. We also included ‘Site’ as a random factor in all models, to account for the matched pair experimental design.
The DHARMa package was employed on all models to perform residual, dispersion, and zero-inflation checks of the data [43]. For all significant models, we performed Tukey pairwise post hoc multiple comparisons tests between fixed effects using the emmeans package [44]. All figures were created using the ggplot2 package [45].
3. Results
Two species of eristaline syrphid flies, the European dronefly, E. tenax, and the golden native dronefly, E. punctulatus, were reared out of both habitat pools at all seven sites. The fly E. tenax was successfully reared from all 14 pools, and E. punctulatus was reared from three of the 14 pools. Additionally, eggs and/or larvae of both species were found within all 14 pools (Table 1). The number of eggs within clutches ranged between 10 and 128 eggs (mean ± S.E. 54.7 ± 3.9 eggs/clutch) in the soil, decaying carrot plants, and water habitat and 15 to 125 eggs (mean ± S.E. 54.4 ± 3.6 eggs/clutch) in the decaying carrot plants and water only habitat.

Table 1.
Developmental stages of eristaline syrphid flies, Eristalis tenax (Linnaeus, 1758) and Eristalinus punctulatus (Macquart, 1847) found in two habitats (1 = soil, carrot plants, and water and 2 = carrot plants and water) at seven sites in the Riverina region of New South Wales, Australia. Both habitats were left to decay for a minimum of 12 days before surveying for fly egg clutches, eggs, and larvae.
Location within the habitat pools also influenced how many eggs were oviposited by female eristaline flies. We found significantly more eggs were oviposited within decaying carrot plant stems and decaying carrot vegetables compared to all other locations (Figure 4). There were no significant differences in the number of eggs laid within the pools based on habitat (p > 0.05 for both, Table S1). Additionally, the number of days the pools were left out to decay did not significantly impact the number of eggs laid within habitat pools (z1,4 = −0.012, p = 0.91).
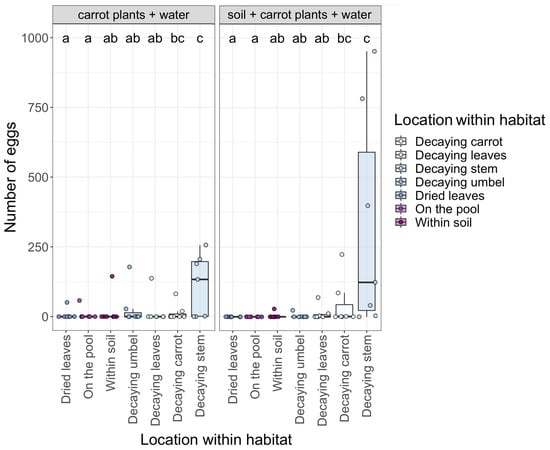
Figure 4.
The number of eggs oviposited by female eristaline syrphid flies within the deployed pools based on habitat (carrot plants + water only and the soil + carrot plants + water) and the location where the eggs were laid. Letters indicate significant differences between locations (p < 0.05). Individual data points representing each habitat pool (n = 14 in total) are jittered onto the figure for clarity. Lower to upper box boundaries indicate the inter-quartile range (IQR). Whiskers are extended to the furthest data point within 1.5x the IQR from each box end.
First, second, and third instar eristaline fly larvae were found within both habitat pools across all sites (Table 2). Significantly more living larvae were found compared to dead larvae (z1,1 = 6.13, p < 0.001); however, the longer the habitat pools were left out to decay, the fewer larvae of all three instars were found in the pools (Figure 5; see Table S2 for statistics). There were no significant differences in the number of larvae found within the pools based on habitat (z1,1 = −0.468, p = 0.64). Additionally, there were no larval instars more abundant than others within the habitat pools (p > 0.05 for all, Table S3).
Table 2.
Total number of larvae found within habitat pools (1 = soil, carrot plants, and water and 2 = carrot plants and water) deployed at seven seed carrot sites in the Riverina region of New South Wales, Australia.
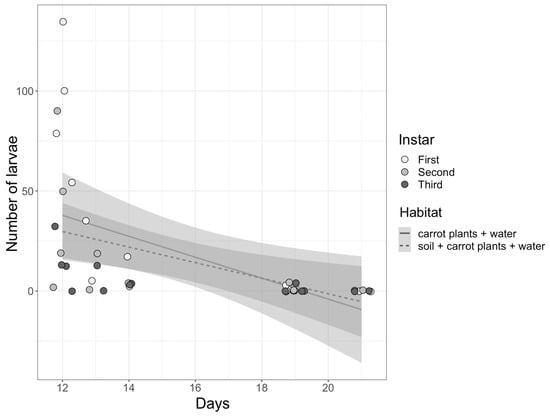
Figure 5.
The number of larvae (first, second and third instar) found within the pools based on habitat (soil, carrot plants, and water or carrot plants and water only) and the number of days the habitat pools were left to decay. Individual data points representing larval instars found per pool (n = 14 per instar; 42 in total) are jittered onto the figure for clarity. The shaded outline of the linear regression lines indicates standard error.
4. Discussion
In this study, we demonstrate that beneficial fly pollinators can utilize small, portable pools filled with locally available, cheap substrates (habitat) in seed carrot agroecosystems as oviposition sites. To our knowledge, no other study has deployed non-floral habitats to provide oviposition sites for pollinating eristaline flies. We found that all the habitat pools contained eggs or larvae of E. tenax, a cosmopolitan fly species that is an effective pollinator of carrot, onion, canola, and other cropping systems [28,46]. We also showed that the habitat pools encouraged oviposition by E. punctulatus, an eristaline fly endemic to the Australasian region [23]; however, other fly species in the genus Eristalinus Rondani, 1845 are known to be effective pollinators of other cropping systems including celery and fennel (Apiaceae), which are close relatives of seed carrot [47]. Species within the Eristalinus genus are found globally and have similar larval habitat and diet requirements [23,25,39]. Therefore, we predict that closely related flies from diverse biogeographical regions will be attracted to the habitat additions tested as well. Both fly species demonstrated oviposition preferences within the habitat, as more eggs were laid on decaying carrot stems, likely because this location within the pool offered protection for the eggs from the sun, preventing the eggs from desiccation, or from predators.
We conducted this study to determine whether habitat pools could host eristaline fly reproduction. Thus, while it has been demonstrated that these pools act as oviposition sites for resident populations of flies, it is unclear how many pools are required to impact pollination services within different-sized fields. While we did observe E. tenax and E. punctulatus flies visiting seed carrot flowers in low numbers before these habitat pools were deployed within fields, it was beyond the scope of the study to compare the effect of habitat pool presence and absence on crop yield.
It is well-known that many species of flies can develop in large numbers from small, transient resources [48,49,50]. The habitat pools tested were proven to be effective and were a quick and easy way to attract flies since both eggs and larvae were found within the habitat after a minimum of 12 days when the seed carrot crop was at peak bloom (50% flowering). For both fly species reared, the time needed to undergo different developmental stages is similar, as eggs hatch after 48 hours, and in optimal conditions, the larvae take an average of 12 days to develop before pupation [23,39]. Although the number of larval instars found did not differ significantly between pools, the majority had recently hatched and were in the first instar of development, and thus likely oviposited 48 to 96 hours previously. Hence, to best facilitate fly pollination during peak crop bloom, we suggest placing the pools nearby a different flowering crop or a small planting of flowers, 12 to 15 days before the flowering onset of the desired crop to ensure that adult eristaline flies locate the pools and two to three generations of syrphids emerge by the time the desired crop reaches peak bloom.
Environmental conditions in the region at the time the habitat pools were deployed significantly influenced pool management. While these results suggest that the deployed habitat pools were low maintenance, we suspect that, under more average (i.e., drier, hotter) environmental conditions at this time of year within the region, this may not be the case. As the Riverina region is typically hot and dry in austral summer, we anticipated refilling the pools with water at least once or moving the pools to a shaded location to ensure the deployed habitat remained a suitable oviposition site for eristaline flies; however, rain events were common when we performed this experiment, so farms became inaccessible to check on the progress of the pools. As the mean temperature between November to December 2021 in the region was 21 °C to 24 °C, none of the habitat pools dried out completely; however, the pools were shallow and not completely shaded, so the sun could have heated up the habitat pools, which could have negatively affected egg and larvae survival. Therefore, we recommend deploying pools in completely shaded environments and monitoring the water level within pools, to ensure that the pools do not become ecological traps for eristaline flies [51].
Similarly, fewer larvae were found within the pools the longer the pools were left out to decay, suggesting the larvae within the pools left undisturbed for longer had either crawled out of the pools to pupate, died competing for food resources, or had been predated upon. As large amounts of decaying carrot plant debris were found within all pools, it is unlikely that the larvae died competing for food resources. The only observed predator within the pools was the rove beetle Creophilus erythrocephalus (Fabricius, 1775) which was present in two pools at the same farm. These rove beetles are known predators of fly larvae [52,53], although they have not been recorded feeding on rat-tailed maggots, specifically. Further research is required to better understand how to scale up these habitats to meet pollination service needs, the length of time the portable habitat pools should be placed on farms, the water conditions that eristaline syrphid fly larvae require to survive, the potential predators of the fly larvae, and whether these pools attract non-target or potential pest species to crop fields.
5. Conclusions
In this study, we successfully trialed a non-floral resource habitat intervention which acted as oviposition sites for beneficial fly pollinators. These habitat pools are a unique, yet cheap, alternative for landholders who want to support pollinating fly reproduction but may not be able to set aside arable land for non-crop habitats. The flies oviposited within decaying carrot plant habitat, and larvae of all instars were found in pools within 12 days. Eristaline flies were found to preferentially oviposit underneath decaying plant stems, likely to protect eggs from predation or adverse environmental conditions. The substrates placed within the habitat pools (soil, discarded carrot plants, and water) are locally available, cheap, and the pools are small and portable, enabling placement and removal at key flowering times. This approach may increase the natural population of flies that provide critical pollination services to crops in intensely managed agricultural systems.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14050439/s1, Figure S1: Location of eggs within habitat pools [22]; Table S1: Egg results [22]; Table S2: Larval results [22]; Table S3: Larval instar pairwise comparisons [22].
Author Contributions
Conceptualization (A.E.D., C.S. and R.R.), methodology (A.E.D., L.A.S., S.H., C.S. and R.R.), validation (S.H., C.S. and R.R.), formal analysis (A.E.D.), investigation (A.E.D. and L.A.S.), resources (A.E.D., L.A.S., S.H. and R.R.), data curation (A.E.D.), writing—original draft preparation (A.E.D.), writing—review and editing (A.E.D., L.A.S., S.H., C.S. and R.R.), visualization (A.E.D., L.A.S. and R.R.), supervision (S.H., C.S. and R.R.), project administration (R.R.), and funding acquisition (R.R.). All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the Horticulture Innovation Australia Limited Project Managing Flies for Crop Pollination (PH16002) and an Australian Research Council Future Fellowship (FT210100851) awarded to R. Rader.
Data Availability Statement
The data presented in this study are available within the paper and its Supplementary Materials.
Acknowledgments
We thank the growers of the sites for allowing us to conduct this study on their properties; Mark Hancock, Mike Tarr and John Hall at South Pacific Seeds for discussion about the study; Karen Santos for help with initial set-up of the habitat pools; Blake Dawson for feedback on the final manuscript; Julius Roberts for help designing Figure 1; several anonymous reviewers for comments and suggestions that have improved the manuscript; and we acknowledge the Wiradjuri people as the traditional owners of the land this research was conducted on and pay respect to Wiradjuri Elders past, present, and future.
Conflicts of Interest
Author S.H. was employed by the company South Pacific Seeds. Author C.S. was employed by the company SeedPurity. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Fijen, T.P.M.; Read, S.F.J.; Walker, M.K.; Gee, M.; Nelson, W.R.; Howlett, B.G. Different landscape features within a simplified agroecosystem support diverse pollinators and their service to crop plants. Landsc. Ecol. 2022, 37, 1787–1799. [Google Scholar] [CrossRef]
- Finch, J.; Gilpin, A.-M.; Cook, J. Fishing for flies: Testing the efficacy of “stink stations” for promoting blow flies as pollinators in mango orchards. J. Pollinat. Ecol. 2023, 32, 79–100. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef]
- Knapp, M.; Rezac, M. Even the smallest non-crop habitat islands could be beneficial: Distribution of carabid beetles and spiders in agricultural landscape. PLoS ONE 2015, 10, e0123052. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: A review. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar] [CrossRef]
- Wilson, J.D.; Morris, A.J.; Arroyo, B.E.; Clark, S.C.; Bradbury, R.B. A review of the abundance and diversity of invertebrate and plant foods of granivorous birds in northern Europe in relation to agricultural change. Agric. Ecosyst. Environ. 1999, 75, 13–30. [Google Scholar] [CrossRef]
- Rischen, T.; Frenzel, T.; Fischer, K. Biodiversity in agricultural landscapes: Different non-crop habitats increase diversity of ground-dwelling beetles (Coleoptera) but support different communities. Biodivers. Conserv. 2021, 30, 3965–3981. [Google Scholar] [CrossRef]
- González, E.; Štrobl, M.; Janšta, P.; Hovorka, T.; Kadlec, T.; Knapp, M. Artificial temporary non-crop habitats support parasitoids on arable land. Biol. Conserv. 2022, 265, 109409. [Google Scholar] [CrossRef]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Schellhorn, N.A.; Bianchi, F.; Hsu, C.L. Movement of Entomophagous Arthropods in Agricultural Landscapes: Links to Pest Suppression. Annu. Rev. Entomol. 2014, 59, 559–581. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.E.; Peisley, R.K.; Rader, R.; Luck, G.W. Pollinators, pests, and predators: Recognizing ecological trade-offs in agroecosystems. Ambio 2016, 45, 4–14. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Scheper, J.; Bommarco, R.; Holzschuh, A.; Potts, S.G.; Riedinger, V.; Roberts, S.P.M.; Rundlof, M.; Smith, H.G.; Steffan-Dewenter, I.; Wickens, J.B.; et al. Local and landscape-level floral resources explain effects of wildflower strips on wild bees across four European countries. J. Appl. Ecol. 2015, 52, 1165–1175. [Google Scholar] [CrossRef]
- Jonsson, A.M.; Ekroos, J.; Danhardt, J.; Andersson, G.K.S.; Olsson, O.; Smith, H.G. Sown flower strips in southern Sweden increase abundances of wild bees and hoverflies in the wider landscape. Biol. Conserv. 2015, 184, 51–58. [Google Scholar] [CrossRef]
- Haenke, S.; Scheid, B.; Schaefer, M.; Tscharntke, T.; Thies, C. Increasing syrphid fly diversity and density in sown flower strips within simple vs. complex landscapes. J. Appl. Ecol. 2009, 46, 1106–1114. [Google Scholar] [CrossRef]
- Gillespie, M.; Wratten, S.; Sedcole, R.; Colfer, R. Manipulating floral resources dispersion for hoverflies (Diptera: Syrphidae) in a California lettuce agro-ecosystem. Biol. Control 2011, 59, 215–220. [Google Scholar] [CrossRef]
- Munoz, A.E.; Plantegenest, M.; Amouroux, P.; Zaviezo, T. Native flower strips increase visitation by non-bee insects to avocado flowers and promote yield. Basic Appl. Ecol. 2021, 56, 369–378. [Google Scholar] [CrossRef]
- Requier, F.; Leonhardt, S.D. Beyond flowers: Including non-floral resources in bee conservation schemes. J. Insect Conserv. 2020, 24, 5–16. [Google Scholar] [CrossRef]
- Davis, A.E.; Bickel, D.; Saunders, M.E.; Rader, R. Crop-pollinating Diptera have diverse diet and habitat needs in both larval and adult stages. Ecol. Appl. 2023, e2859. [Google Scholar] [CrossRef]
- Campoy, A.; Pérez-Bañón, C.; Aznar, D.; Rojo, S. Description of the preimaginal stages of the golden native dronefly from Australia, Eristalinus punctulatus (Macquart, 1847) (Diptera: Syrphidae). Austral Entomol. 2020, 59, 784–793. [Google Scholar] [CrossRef]
- Davis, A.E.; Deutsch, K.R.; Torres, A.M.; Loya, M.J.M.; Cody, L.V.; Harte, E.; Sossa, D.; Muniz, P.A.; Ng, W.H.; McArt, S.H. Eristalis flower flies can be mechanical vectors of the common trypanosome bee parasite, Crithidia bombi. Sci. Rep. 2021, 11, 15852. [Google Scholar] [CrossRef] [PubMed]
- Young, A.; Marshall, S.; Locke, M.; Moran, K.; Crins, W.; Skevington, J. Field Guide to the Flower Flies of Northeastern North America; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- Jauker, F.; Bondarenko, B.; Becker, H.C.; Steffan-Dewenter, I. Pollination efficiency of wild bees and hoverflies provided to oilseed rape. Agric. For. Entomol. 2012, 14, 81–87. [Google Scholar] [CrossRef]
- Jarlan, A.; De Oliveira, D.; Gingras, J. Effects of Eristalis tenax (Diptera: Syrphidae) pollination on characteristics of greenhouse sweet pepper fruits. J. Econ. Entomol. 1997, 90, 1650–1654. [Google Scholar] [CrossRef]
- Cook, D.; Voss, S.; Finch, J.; Rader, R.; Cook, J.; Spurr, C. The role of flies as pollinators of horticultural crops: An Australian case study with worldwide relevance. Insects 2020, 11, 341. [Google Scholar] [CrossRef]
- Howlett, B.G.; Gee, M. The potential management of the drone fly (Eristalis tenax) as a crop pollinator in New Zealand. N. Z. Plant Prot. 2019, 72, 221–230. [Google Scholar] [CrossRef]
- Gaffney, A.; Allen, G.R.; Brown, P.H. Insect visitation to flowering hybrid carrot seed crops. N. Z. J. Crop Hortic. Sci. 2011, 39, 79–93. [Google Scholar] [CrossRef]
- Gaffney, A.; Bohman, B.; Quarrell, S.R.; Brown, P.H.; Allen, G.R. Frequent insect visitors are not always pollen carriers in hybrid carrot pollination. Insects 2018, 9, 61. [Google Scholar] [CrossRef]
- Broussard, M.A.; Mas, F.; Howlett, B.; Pattemore, D.; Tylianakis, J.M. Possible mechanisms of pollination failure in hybrid carrot seed and implications for industry in a changing climate. PLoS ONE 2017, 12, e0180215. [Google Scholar] [CrossRef] [PubMed]
- Perez-Banon, C.; Petanidou, T.; Marcos-Garcia, M.A. Pollination in small islands by occasional visitors: The case of Daucus carota subsp commutatus (Apiaceae) in the Columbretes archipelago, Spain. Plant Ecol. 2007, 192, 133–151. [Google Scholar] [CrossRef]
- Reilly, J.; Artz, D.; Biddinger, D.; Bobiwash, K.; Boyle, N.; Brittain, C.; Brokaw, J.; Campbell, J.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators. Proc. R. Soc. B-Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef] [PubMed]
- Dolley, W.L., Jr.; Hassett, C.C.; Bowen, W.J. Laboratory culture of the drone fly, Eristalis tenax. Science 1933, 78, 313–314. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, S.; Thyselius, M.; Holden, M.; Nordstrom, K. Rearing and long-Term maintenance of Eristalis tenax hoverflies for research studies. J. Vis. Exp. 2018, 135, 57711. [Google Scholar] [CrossRef]
- Campoy, A.; Sáez, L.; Pérez-Bañón, C.; Rojo, S. Demography and population parameters of two species of eristaline flower flies (Diptera, Syrphidae, Eristalini). J. Appl. Entomol. 2020, 144, 133–143. [Google Scholar] [CrossRef]
- Campoy, A.; Aracil, A.; Pérez-Bañón, C.; Rojo, S. An in-depth study of the larval head skeleton and the external feeding structures related with the ingestion of food particles by the eristaline flower flies Eristalis tenax and Eristalinus aeneus. Entomol. Exp. Appl. 2020, 168, 783–798. [Google Scholar] [CrossRef]
- Campoy, A.; Pérez-Bañón, C.; Rojo, S. Intra-puparial development in the hoverflies Eristalinus aeneus and Eristalis tenax (Diptera: Syrphidae). J. Morphol. 2020, 281, 1436–1445. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2022. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 10 October 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rader, R.; Bartomeus, I.; Garibaldi, L.; Garratt, M.; Howlett, B.; Winfree, R.; Cunningham, S.; Mayfield, M.; Arthur, A.; Andersson, G.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; Belliure, B.; Montserrat, M.; Gil, J.; Velásquez, Y. Pollination by the hoverfly Eristalinus aeneus (Diptera: Syrphidae) in two hybrid seed crops: Celery and fennel (Apiaceae). J. Agric. Sci. 2022, 160, 1–13. [Google Scholar] [CrossRef]
- Levot, G.W.; Brown, K.R.; Shipp, E. Larval growth of some calliphorid and sarcophagid Diptera. Bull. Entomol. Res. 1979, 69, 469–475. [Google Scholar] [CrossRef]
- Carey, J.R.; Liedo, P.; Muller, H.G.; Wang, J.L.; Chiou, J.M. Relationship of age patterns of fecundity to mortality, longevity, and lifetime reproduction in a large cohort of Mediterranean fruit fly females. J. Gerontol. Ser. A-Biol. Sci. Med. Sci. 1998, 53, B245–B251. [Google Scholar] [CrossRef]
- Vasconcelos, S.D.; Cruz, T.M.; Salgado, R.L.; Thyssen, P.J. Dipterans associated with a decomposing animal carcass in a rainforest fragment in Brazil: Notes on the early arrival and colonization by necrophagous species. J. Insect Sci. 2013, 13, 145. [Google Scholar] [CrossRef]
- Cox, M.E.; Johnstone, R.; Robinson, J. Relationships between perceived coastal waterway condition and social aspects of quality of life. Ecol. Soc. 2006, 11, 35. Available online: https://www.jstor.org/stable/26267792 (accessed on 15 October 2022). [CrossRef]
- Greene, G.L. Rearing techniques for Creophilus maxillosus (Coleoptera: Staphylinidae), a predator of fly larvae in cattle feedlots. J. Econ. Entomol. 1996, 89, 848–851. [Google Scholar] [CrossRef]
- Bornemissza, G.F. An analysis of arthropod succession in carrion and the effect of its decomposition on the soil fauna. Aust. J. Zool. 1957, 5, 1–12. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).