
Evolution of piggyBac Transposons in Apoidea

 ,
,  , and
, and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. piggyBac Mining in Apoidea
2.2. Sequence Analysis and Phylogenetic Analysis
2.3. Evolutionary Dynamics Analysis
3. Results
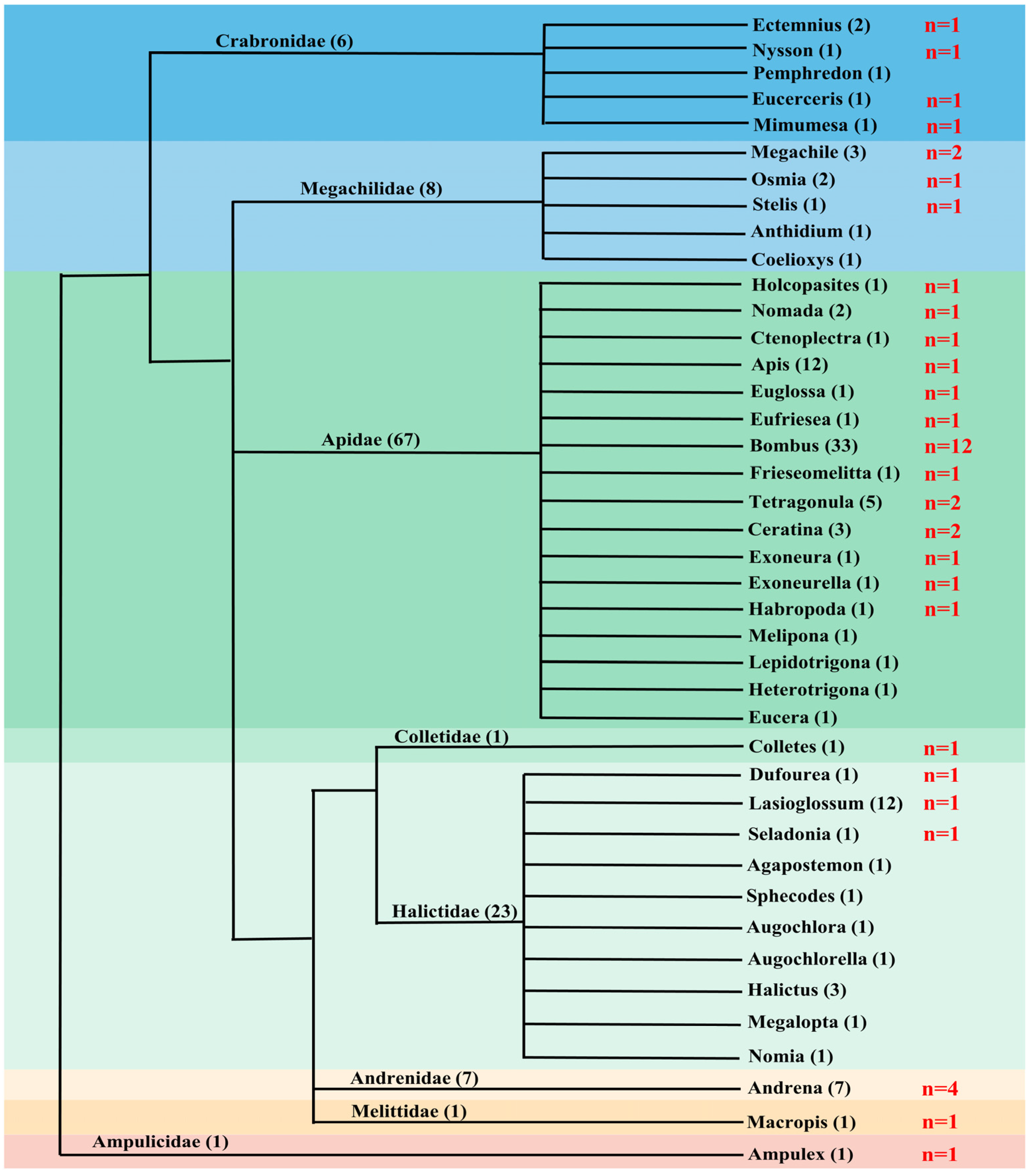
3.1. Distribution of piggyBac Transposons in Apoidea
3.2. Low Abundance of piggyBac Transposons in Apoidea
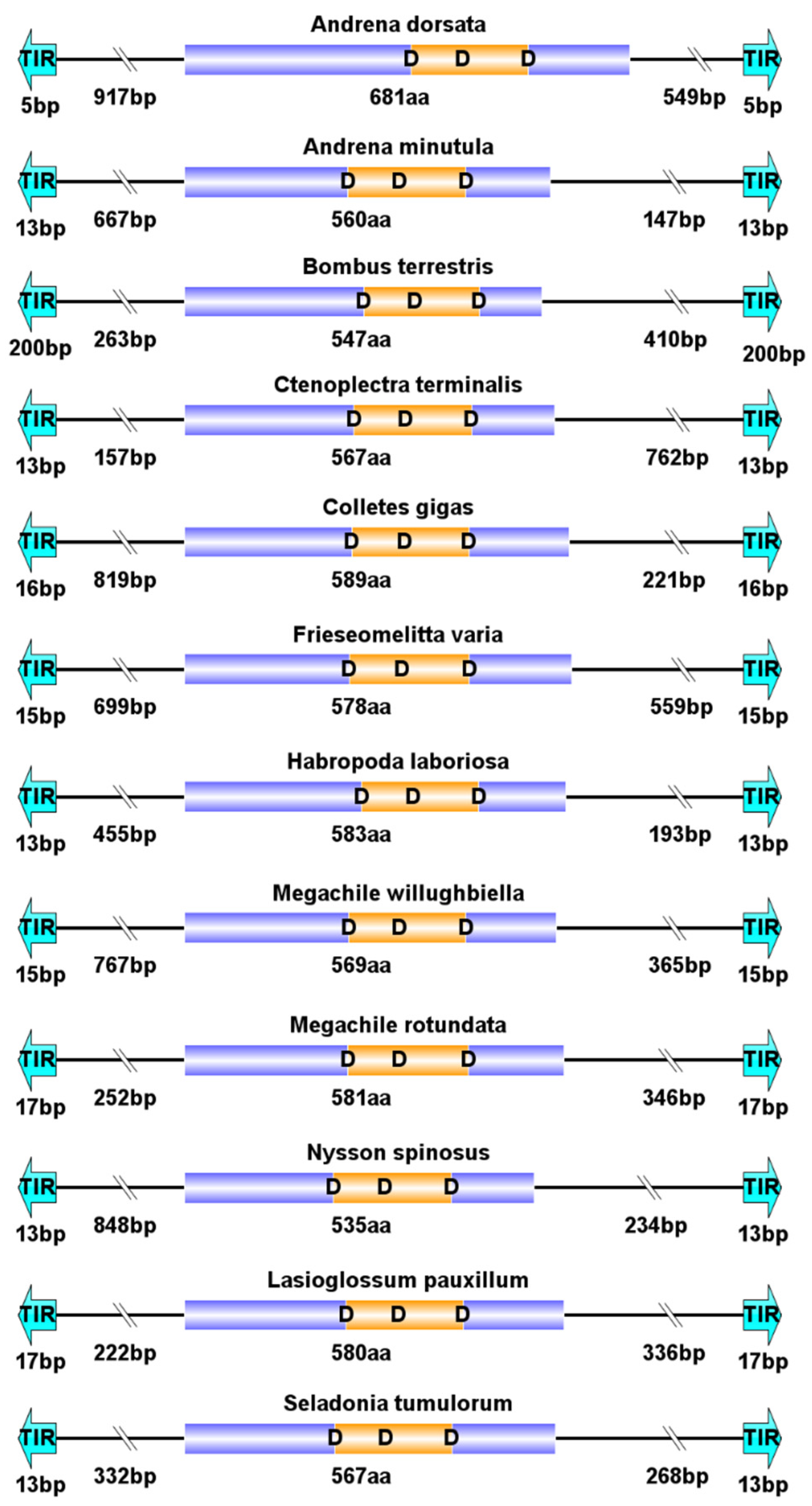
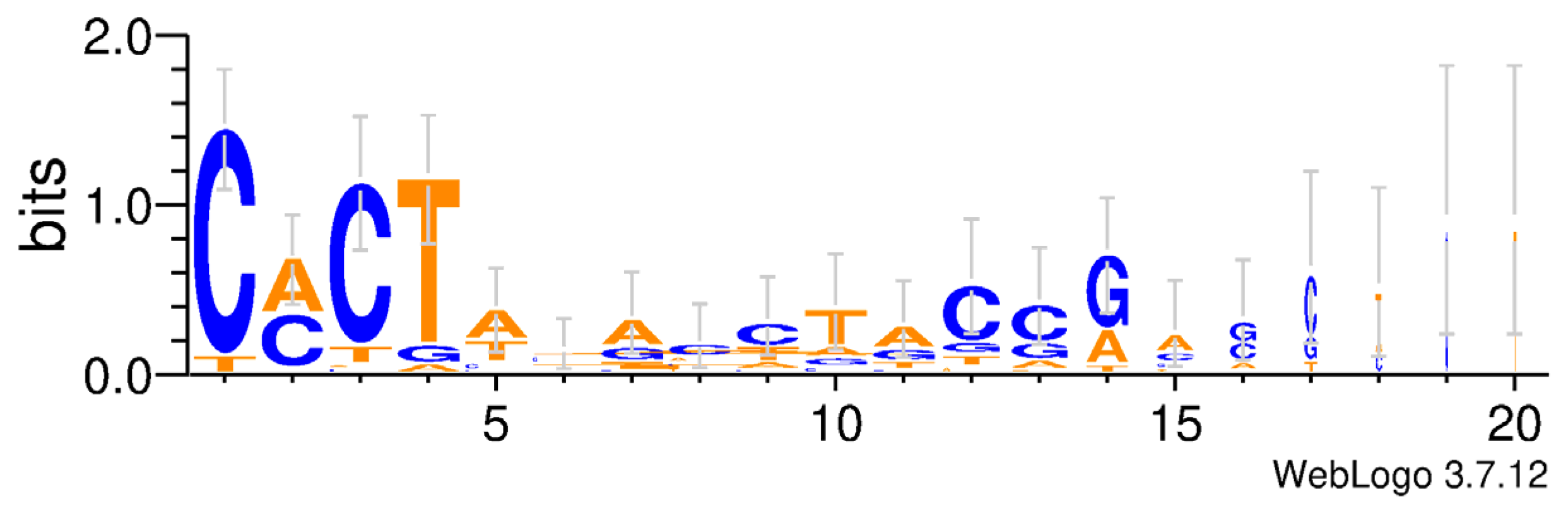
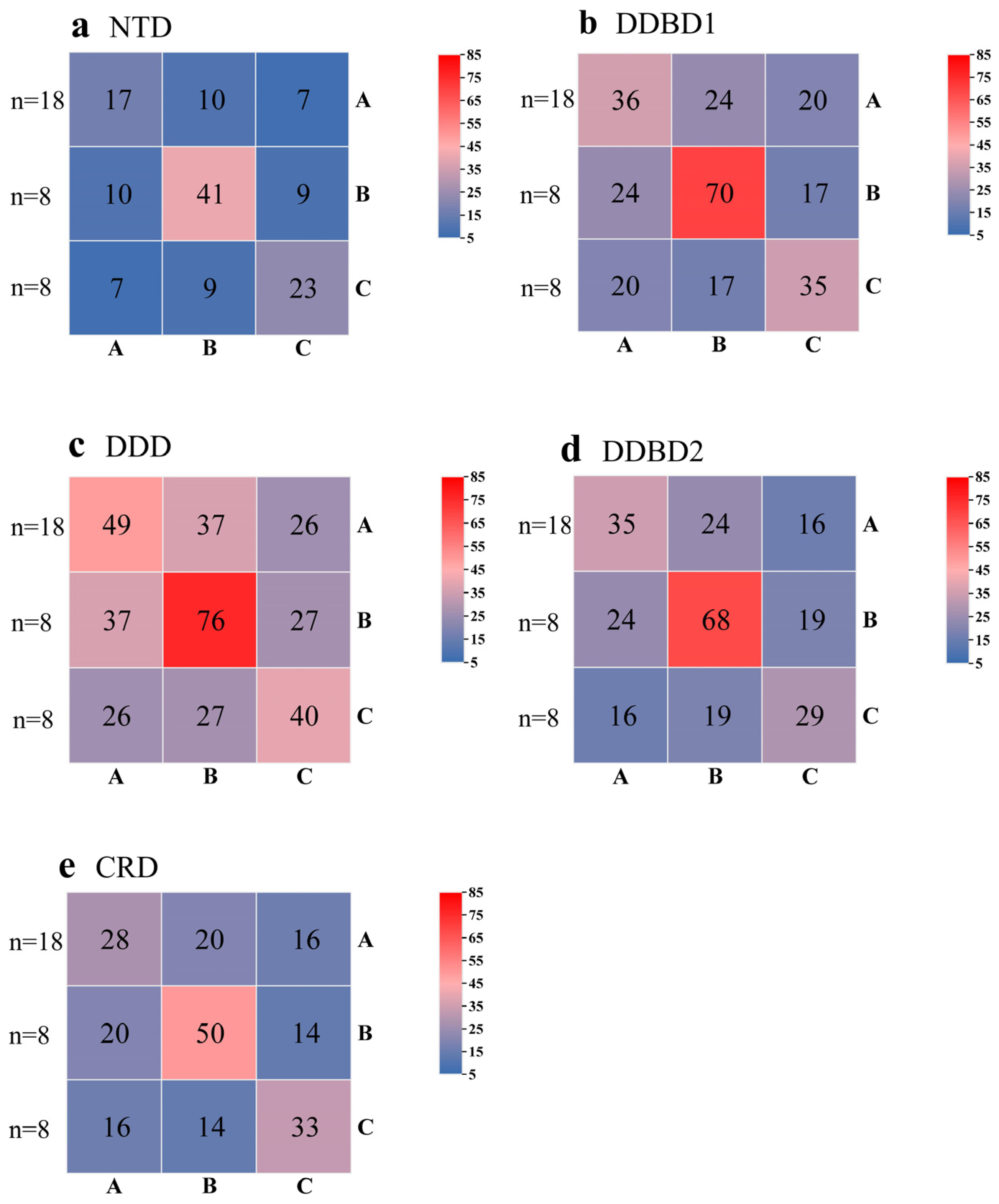
3.3. Structural Organization of piggyBac Transposons in Apoidea
3.4. Transposase Domain Organization
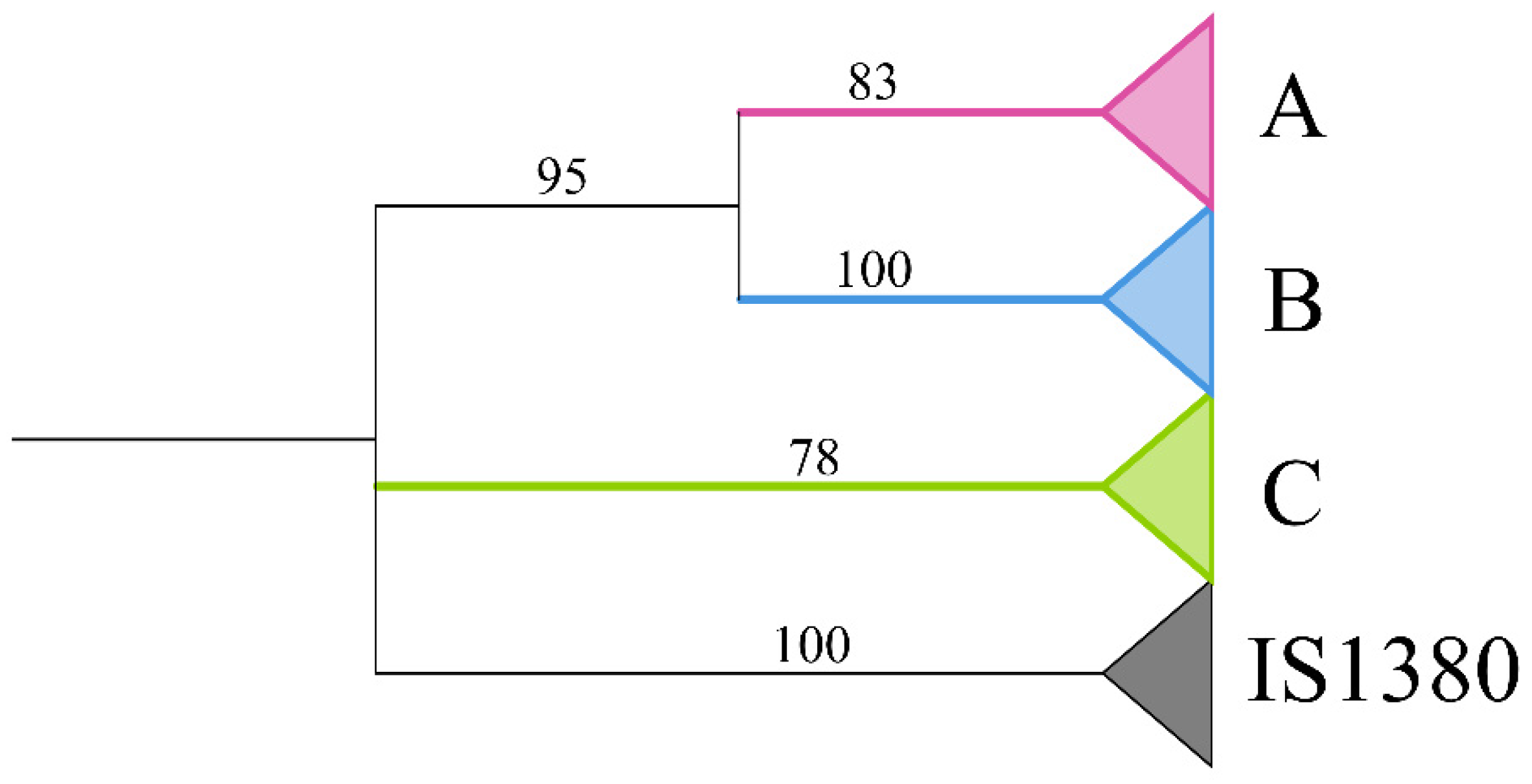
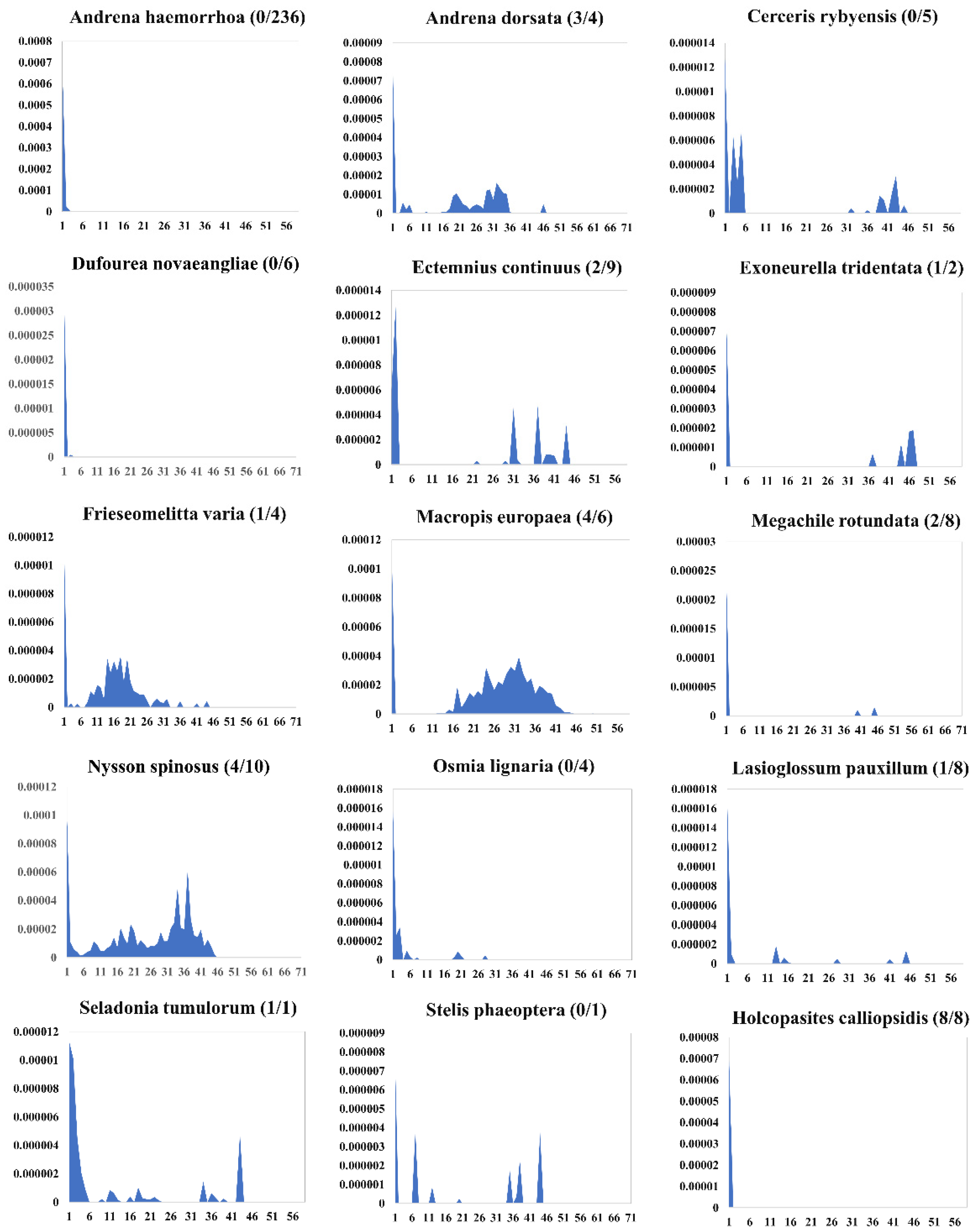
3.5. Evolution Dynamics of piggyBac in the Apoidea Genomes
4. Discussion
4.1. Distribution, Diversity and Copy Number of piggyBac in Apoidea
4.2. Structure Organization of piggyBac
4.3. Evolution Dynamics of piggyBac in Apoidea
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Danforth, B.N.; Sipes, S.; Fang, J.; Brady, S.G. The history of early bee diversification based on five genes plus morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 15118–15123. [Google Scholar] [CrossRef]
- Sann, M.; Niehuis, O.; Peters, R.S.; Mayer, C.; Kozlov, A.; Podsiadlowski, L.; Bank, S.; Meusemann, K.; Misof, B.; Bleidorn, C.; et al. Phylogenomic analysis of Apoidea sheds new light on the sister group of bees. BMC Evol. Biol. 2018, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D. The Professional Development of an Entomologist. Annu. Rev. Entomol. 2007, 52, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.-Y.; Cao, L.-J.; Tang, P.; van Achterberg, K.; Hoffmann, A.A.; Chen, H.-Y.; Chen, X.-X.; Wei, S.-J. Gene arrangement and sequence of mitochondrial genomes yield insights into the phylogeny and evolution of bees and sphecid wasps (Hymenoptera: Apoidea). Mol. Phylogenet. Evol. 2018, 124, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ohl, M.; Engel, M.S. Die Fossilgeschichte der Bienen und Ihrer Nächsten Verwandten (Hymenoptera: Apoidea). Denisia 2007, 20, 687–700. [Google Scholar]
- Debevec, A.H.; Cardinal, S.; Danforth, B.N. Identifying the sister group to the bees: A molecular phylogeny of Aculeata with an emphasis on the superfamily Apoidea. Zool. Scr. 2012, 41, 527–535. [Google Scholar] [CrossRef]
- Cardinal, S.; Danforth, B. The Antiquity and Evolutionary History of Social Behavior in Bees. PLoS ONE 2011, 6, e21086. [Google Scholar] [CrossRef] [PubMed]
- Cardinal, S.; Danforth, B. Bees diversified in the age of eudicots. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122686. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Patiny, S.; Danforth, B.N. The bee tree of life: A supermatrix approach to apoid phylogeny and biogeography. BMC Evol. Biol. 2013, 13, 138. [Google Scholar] [CrossRef]
- Danforth, B.N.; Fang, J.; Sipes, S. Analysis of family-level relationships in bees (Hymenoptera: Apiformes) using 28S and two previously unexplored nuclear genes: CAD and RNA polymerase II. Mol. Phylogenet. Evol. 2006, 39, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Danforth, B.N.; Cardinal, S.; Praz, C.; Almeida, E.A.; Michez, D. The Impact of Molecular Data on Our Understanding of Bee Phylogeny and Evolution. Annu. Rev. Entomol. 2013, 58, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zong, W.; Diaby, M.; Lin, Z.; Wang, S.; Gao, B.; Ji, T.; Song, C. Diversity and Evolution of pogo and Tc1/mariner Transposons in the Apoidea Genomes. Biology 2021, 10, 940. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Felipe, I.; Blanco, C.A.; Corona, M. Impact of Apoidea (Hymenoptera) on the World’s Food Production and Diets. Ann. Entomol. Soc. Am. 2020, 113, 407–424. [Google Scholar] [CrossRef]
- Hepburn, H. The Bees of the World. Afr. Zool. 2001, 36, 117. [Google Scholar] [CrossRef]
- Sang, Y.; Gao, B.; Diaby, M.; Zong, W.; Chen, C.; Shen, D.; Wang, S.; Wang, Y.; Ivics, Z.; Song, C. Incomer, a DD36E family of Tc1/mariner transposons newly discovered in animals. Mob. DNA 2019, 10, 45. [Google Scholar] [CrossRef]
- Han, M.-J.; Xu, H.-E.; Zhang, H.-H.; Feschotte, C.; Zhang, Z. Spy: A New Group of Eukaryotic DNA Transposons without Target Site Duplications. Genome Biol. Evol. 2014, 6, 1748–1757. [Google Scholar] [CrossRef]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.C.; Wides, R.; et al. The Genome Sequence of the Malaria Mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef]
- Kapitonov, V.V.; Jurka, J. Molecular paleontology of transposable elements in the Drosophila melanogaster genome. Proc. Natl. Acad. Sci. USA 2003, 100, 6569–6574. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome Sequence of Aedes aegypti, a Major Arbovirus Vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef]
- Xu, H.-E.; Zhang, H.-H.; Xia, T.; Han, M.-J.; Shen, Y.-H.; Zhang, Z. BmTEdb: A collective database of transposable elements in the silkworm genome. Database 2013, 2013, bat055. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; Fitzhugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [PubMed]
- Chinwalla, A.T.; Cook, L.L.; Delehaunty, K.D.; Fewell, G.A.; Fulton, L.A.; Fulton, R.S.; Graves, T.A.; Hillier, L.W.; Mardis, E.R.; McPherson, J.D.; et al. Initial Sequencing and Comparative Analysis of the Mouse Genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- De Koning, A.P.J.; Gu, W.; Castoe, T.A.; Batzer, M.A.; Pollock, D.D. Repetitive Elements May Comprise Over Two-Thirds of the Human Genome. PLoS Genet. 2011, 7, e1002384. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C.E.D.; Jiang, N.; Wessler, S.R. Plant transposable elements: Where genetics meets genomics. Nat. Rev. Genet. 2002, 3, 329–341. [Google Scholar] [CrossRef]
- Kidwell, M.G. Transposable elements and the evolution of genome size in eukaryotes. Genetica 2002, 115, 49–63. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.; Xia, J.; Jin, Y. Domestication of Transposable Elements into MicroRNA Genes in Plants. PLoS ONE 2011, 6, e19212. [Google Scholar] [CrossRef] [PubMed]
- Rebollo, R.; Romanish, M.T.; Mager, D.L. Transposable Elements: An Abundant and Natural Source of Regulatory Sequences for Host Genes. Annu. Rev. Genet. 2012, 46, 21–42. [Google Scholar] [CrossRef]
- Almeida, L.; Silva, I.T.; Silva, W.A.; Castro, J.P.; Riggs, P.K.; Carareto, C.M.; Amaral, M.E.J. The contribution of transposable elements to Bos taurus gene structure. Gene 2007, 390, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Mobile DNA and evolution in the 21st century. Mob. DNA 2010, 1, 4. [Google Scholar] [CrossRef]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Kojima, K.K. Structural and sequence diversity of eukaryotic transposable elements. Genes Genet. Syst. 2019, 94, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Cary, L.C.; Goebel, M.; Corsaro, B.G.; Wang, H.-G.; Rosen, E.; Fraser, M.J. Transposon mutagenesis of baculoviruses: Analysis of Trichoplusia ni transposon IFP2 insertions within the FP-locus of nuclear polyhedrosis viruses. Virology 1989, 172, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J.; Smith, G.E.; Summers, M.D. Acquisition of Host Cell DNA Sequences by Baculoviruses: Relationship between Host DNA Insertions and FP Mutants of Autographa californica and Galleria mellonella Nuclear Polyhedrosis Viruses. J. Virol. 1983, 47, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Sim, C.; Hong, Y.S.; Hogan, J.R.; Fraser, M.J.; Robertson, H.M.; Collins, F.H. Molecular evolutionary analysis of the widespread piggyBac transposon family and related “domesticated” sequences. Mol. Genet. Genom. 2003, 270, 173–180. [Google Scholar] [CrossRef]
- Bouallègue, M.; Rouault, J.-D.; Hua-Van, A.; Makni, M.; Capy, P. Molecular evolution of piggyBac superfamily: From selfishness to domestication. Genome Biol. Evol. 2017, 9, 323–339. [Google Scholar] [CrossRef]
- Ding, S.; Wu, X.; Li, G.; Han, M.; Zhuang, Y.; Xu, T. Efficient Transposition of the piggyBac (PB) Transposon in Mammalian Cells and Mice. Cell 2005, 122, 473–483. [Google Scholar] [CrossRef]
- Yusa, K.; Zhou, L.; Li, M.A.; Bradley, A.; Craig, N.L. A hyperactive piggyBac transposase for mammalian applications. Proc. Natl. Acad. Sci. USA 2011, 108, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Manuri, P.V.R.; Wilson, M.H.; Maiti, S.N.; Mi, T.; Singh, H.; Olivares, S.; Dawson, M.J.; Huls, H.; Lee, D.A.; Rao, P.H.; et al. piggyBac Transposon/Transposase System to Generate CD19-Specific T Cells for the Treatment of B-Lineage Malignancies. Hum. Gene Ther. 2010, 21, 427–437. [Google Scholar] [CrossRef]
- Kolacsek, O.; Wachtl, G.; Fóthi, Á.; Schamberger, A.; Sándor, S.; Pergel, E.; Varga, N.; Raskó, T.; Izsvák, Z.; Apáti, Á.; et al. Functional indications for transposase domestications–Characterization of the human piggyBac transposase derived (PGBD) activities. Gene 2022, 834, 146609. [Google Scholar] [CrossRef]
- Fraser, M.J.; Clszczon, T.; Elick, T.; Bauser, C. Precise excision of TTAA-specific lepidopteran transposons piggyBac (IFP2) and tagalong (TFP3) from the baculovirus genome in cell lines from two species of Lepidoptera. Insect Mol. Biol. 1996, 5, 141–151. [Google Scholar] [CrossRef]
- Lobo, N.; Li, X.; Fraser, M.J. Transposition of the piggyBac element in embryos of Drosophila melanogaster, Aedes aegypti and Trichoplusiani. Mol. Gen. Genet. 1999, 261, 803–810. [Google Scholar] [CrossRef]
- Yusa, K. piggyBac Transposon. Microbiol. Spectr. 2015, 3, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Fain-Thornton, J.; Craig, N.L. piggyBac can bypass DNA synthesis during cut and paste transposition. EMBO J. 2008, 27, 1097–1109. [Google Scholar] [CrossRef]
- Chen, Q.; Luo, W.; Veach, R.A.; Hickman, A.B.; Wilson, M.H.; Dyda, F. Structural basis of seamless excision and specific targeting by piggyBac transposase. Nat. Commun. 2020, 11, 3446. [Google Scholar] [CrossRef]
- Gibbs, R.A.; Weinstock, G.M.; Metzker, M.L.; Muzny, D.M.; Sodergren, E.J.; Scherer, S.; Scott, G.; Steffen, D.; Worley, K.C.; Burch, P.E.; et al. Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature 2004, 428, 493–521. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., III; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Pace, J.K.; Feschotte, C. The evolutionary history of human DNA transposons: Evidence for intense activity in the primate lineage. Genome Res. 2007, 17, 422–432. [Google Scholar] [CrossRef]
- Ray, D.A.; Feschotte, C.; Pagan, H.J.; Smith, J.D.; Pritham, E.J.; Arensburger, P.; Atkinson, P.W.; Craig, N.L. Multiple waves of recent DNA transposon activity in the bat, Myotis lucifugus. Genome Res. 2008, 18, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Pagan, H.J.T.; Smith, J.D.; Hubley, R.M.; Ray, D.A. PiggyBac-ing on a Primate Genome: Novel Elements, Recent Activity and Horizontal Transfer. Genome Biol. Evol. 2010, 2, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Du, Y.; Wang, S.; Brown, S.J.; Park, Y. Large diversity of the piggyBac-like elements in the genome of Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Ebert, P.R.; Hileman, J.P.; Nguyen, H.T. Primary sequence, copy number, and distribution of mariner transposons in the honey bee. Insect Mol. Biol. 1995, 4, 69–78. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Lee, D.; Schulman, A. FastPCR Software for PCR Primer and Probe Design and Repeat Search. Genes Genomes Genom. 2009, 3, 1–14. [Google Scholar]
- Yang, P.; Craig, P.A.; Goodsell, D.; Bourne, P.E. BioEditor—Simplifying macromolecular structure annotation. Bioinformatics 2003, 19, 897–898. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Yamada, K.D.; Tomii, K.; Katoh, K. Application of the MAFFT sequence alignment program to large data—Reexamination of the usefulness of chained guide trees. Bioinformatics 2016, 32, 3246–3251. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xie, Y.; Ma, J.; Luo, X.; Nie, P.; Zuo, Z.; Lahrmann, U.; Zhao, Q.; Zheng, Y.; Zhao, Y.; et al. IBS: An illustrator for the presentation and visualization of biological sequences. Bioinformatics 2015, 31, 3359–3361. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Hickman, A.B.; Chandler, M.; Dyda, F. Integrating prokaryotes and eukaryotes: DNA transposases in light of structure. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 50–69. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Tarailo-Graovac, M.; Chen, N. Using RepeatMasker to Identify Repetitive Elements in Genomic Sequences. Curr. Protoc. Bioinform. 2009, 25, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Dunemann, S.M.; Wasmuth, J.D. Horizontal transfer of a retrotransposon between parasitic nematodes and the common shrew. Mob. DNA 2019, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.-W.; Wessler, S.R. The catalytic domain of all eukaryotic cut-and-paste transposase superfamilies. Proc. Natl. Acad. Sci. USA 2011, 108, 7884–7889. [Google Scholar] [CrossRef]
- Ghanim, G.E.; Kellogg, E.H.; Nogales, E.; Rio, D.C. Structure of a P element transposase–DNA complex reveals unusual DNA structures and GTP-DNA contacts. Nat. Struct. Mol. Biol. 2019, 26, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Keith, J.H.; Schaeper, C.A.; Fraser, T.S.; Fraser, M.J. Mutational analysis of highly conserved aspartate residues essential to the catalytic core of the piggyBac transposase. BMC Mol. Biol. 2008, 9, 73. [Google Scholar] [CrossRef]
- Horn, C.; Offen, N.; Nystedt, S.; Häcker, U.; Wimmer, E.A. piggyBac-Based Insertional Mutagenesis and Enhancer Detection as a Tool for Functional Insect Genomics. Genetics 2003, 163, 647–661. [Google Scholar] [CrossRef]
- Aparicio, S.; Chapman, J.; Stupka, E.; Putnam, N.; Chia, J.-M.; Dehal, P.; Christoffels, A.; Rash, S.; Hoon, S.; Smit, A.; et al. Whole-Genome Shotgun Assembly and Analysis of the Genome of Fugu rubripes. Science 2002, 297, 1301–1310. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zong, W.; Miskey, C.; Ullah, N.; Diaby, M.; Chen, C.; Wang, X.; Ivics, Z.; Song, C. Intruder (DD38E), a recently evolved sibling family of DD34E/Tc1 transposons in animals. Mob. DNA 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.; Gao, B.; Miskey, C.; Chen, C.; Sang, Y.; Zong, W.; Wang, S.; Wang, Y.; Wang, X.; Ivics, Z.; et al. Multiple Invasions of Visitor, a DD41D Family of Tc1/mariner Transposons, throughout the Evolution of Vertebrates. Genome Biol. Evol. 2020, 12, 1060–1073. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Shi, S.; Diaby, M.; Danley, P.; Ullah, N.; Puzakov, M.; Gao, B.; Song, C. Horizontal transfer of Buster transposons across multiple phyla and classes of animals. Mol. Phylogenet. Evol. 2022, 173, 107506. [Google Scholar] [CrossRef]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef]
- Shi, S.; Puzakov, M.; Guan, Z.; Xiang, K.; Diaby, M.; Wang, Y.; Wang, S.; Song, C.; Gao, B. Prokaryotic and Eukaryotic Horizontal Transfer of Sailor (DD82E), a New Superfamily of IS630-Tc1-Mariner DNA Transposons. Biology 2021, 10, 1005. [Google Scholar] [CrossRef]
- Jia, W.; Asare, E.; Liu, T.; Zhang, P.; Wang, Y.; Wang, S.; Shen, D.; Miskey, C.; Gao, B.; Ivics, Z.; et al. Horizontal Transfer and Evolutionary Profiles of Two Tc1/DD34E Transposons (ZB and SB) in Vertebrates. Genes 2022, 13, 2239. [Google Scholar] [CrossRef] [PubMed]
- Elick, T.A.; Lobo, N.; Fraser, M.J. Analysis of the cis-acting DNA elements required for piggyBac transposable element excision. Mol. Genet. Genom. 1997, 255, 605–610. [Google Scholar] [CrossRef]
- Li, X.; Lobo, N.; Bauser, C.A.; Fraser, M.J. The minimum internal and external sequence requirements for transposition of the eukaryotic transformation vector piggyBac. Mol. Genet. Genom. 2001, 266, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Haniford, D.; Kleckner, N. Tn 10 transposition in vivo: Temporal separation of cleavages at the two transposon ends and roles of terminal basepairs subsequent to interaction of ends. EMBO J. 1994, 13, 3401–3411. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distribution a | Ampulicidae | Andrenidae | Apidae | Colletidae | Crabronidae | Halictidae | Megachilidae | Melittidae |
|---|---|---|---|---|---|---|---|---|
| Full Copy Number | 3 | 2–236 | 1–335 | 4 | 4–10 | 1–8 | 1–8 | 6 |
| Average b | 3.00 | 61.25 | 30.00 | 4.00 | 7.00 | 5.00 | 4.75 | 6.00 |
| Number of Species with Full Transposons | 1 | 4 | 26 | 1 | 4 | 3 | 4 | 1 |
| Intact Copy Number | 1 | 0–3 | 0–8 | 1 | 0–4 | 0–1 | 0–3 | 4 |
| Average C | 1.00 | 1.25 | 0.62 | 1.00 | 2.00 | 0.67 | 1.25 | 4.00 |
| Number of Species with Intact Transposons | 1 | 2 | 8 | 1 | 3 | 2 | 2 | 1 |
| Distribution a | Species Number | Length of Full Transposons (bp) b | Number of Species with Full Transposons | Length of Intact Transposon (bp) C | Number of Species with Intact Transposons | Transposase Length (aa) | TIR Length (bp) | TSD |
|---|---|---|---|---|---|---|---|---|
| Ampulicidae | 1 | 2380 | 1 | 2380 | 1 | 552 | 14 | TTAA |
| Andrenidae | 4 | 2519–3518 | 4 | 2519–3518 | 2 | 83–681 | 5–23 | TTAA |
| Apidae | 26 | 368–3518 | 26 | 1670–3413 | 8 | 24–612 | 7–493 | TTAA |
| Colletidae | 1 | 2838 | 1 | 2838 | 1 | 589 | 16 | TTAA |
| Crabronidae | 4 | 2334–2712 | 4 | 2334–2712 | 3 | 449–601 | 13–18 | TTAA |
| Halictidae | 3 | 2326–2596 | 3 | 2326–2596 | 2 | 477–580 | 13–17 | TTAA |
| Megachilidae | 4 | 900–2868 | 4 | 2292–2868 | 2 | 206–581 | 15–20 | TTAA |
| Melittidae | 1 | 2646 | 1 | 2646 | 1 | 567 | 13 | TTAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Guan, Z.; Wang, F.; Wang, Y.; Asare, E.; Shi, S.; Lin, Z.; Ji, T.; Gao, B.; Song, C. Evolution of piggyBac Transposons in Apoidea. Insects 2023, 14, 402. https://doi.org/10.3390/insects14040402
Li X, Guan Z, Wang F, Wang Y, Asare E, Shi S, Lin Z, Ji T, Gao B, Song C. Evolution of piggyBac Transposons in Apoidea. Insects. 2023; 14(4):402. https://doi.org/10.3390/insects14040402
Chicago/Turabian StyleLi, Xueyuan, Zhongxia Guan, Feng Wang, Yali Wang, Emmanuel Asare, Shasha Shi, Zheguang Lin, Ting Ji, Bo Gao, and Chengyi Song. 2023. "Evolution of piggyBac Transposons in Apoidea" Insects 14, no. 4: 402. https://doi.org/10.3390/insects14040402
APA StyleLi, X., Guan, Z., Wang, F., Wang, Y., Asare, E., Shi, S., Lin, Z., Ji, T., Gao, B., & Song, C. (2023). Evolution of piggyBac Transposons in Apoidea. Insects, 14(4), 402. https://doi.org/10.3390/insects14040402