


Insecticide Resistance of Cimex lectularius L. Populations and the Performance of Selected Neonicotinoid-Pyrethroid Mixture Sprays and an Inorganic Dust


Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Bed Bug Populations
2.2. Resistance to Two Neonicotinoids and a Pyrethroid Insecticide Using Topical Assays
2.3. Performance of Formulated Insecticides
2.3.1. Neonicotinoid-Pyrethroid Mixture Sprays
2.3.2. Inorganic Dust
2.4. Data Analysis
3. Results
3.1. Resistance to Two Neonicotinoids and a Pyrethroid Insecticide
3.2. Performance of Formulated Insecticides
3.2.1. Neonicotinoid-Pyrethroid Mixture Sprays
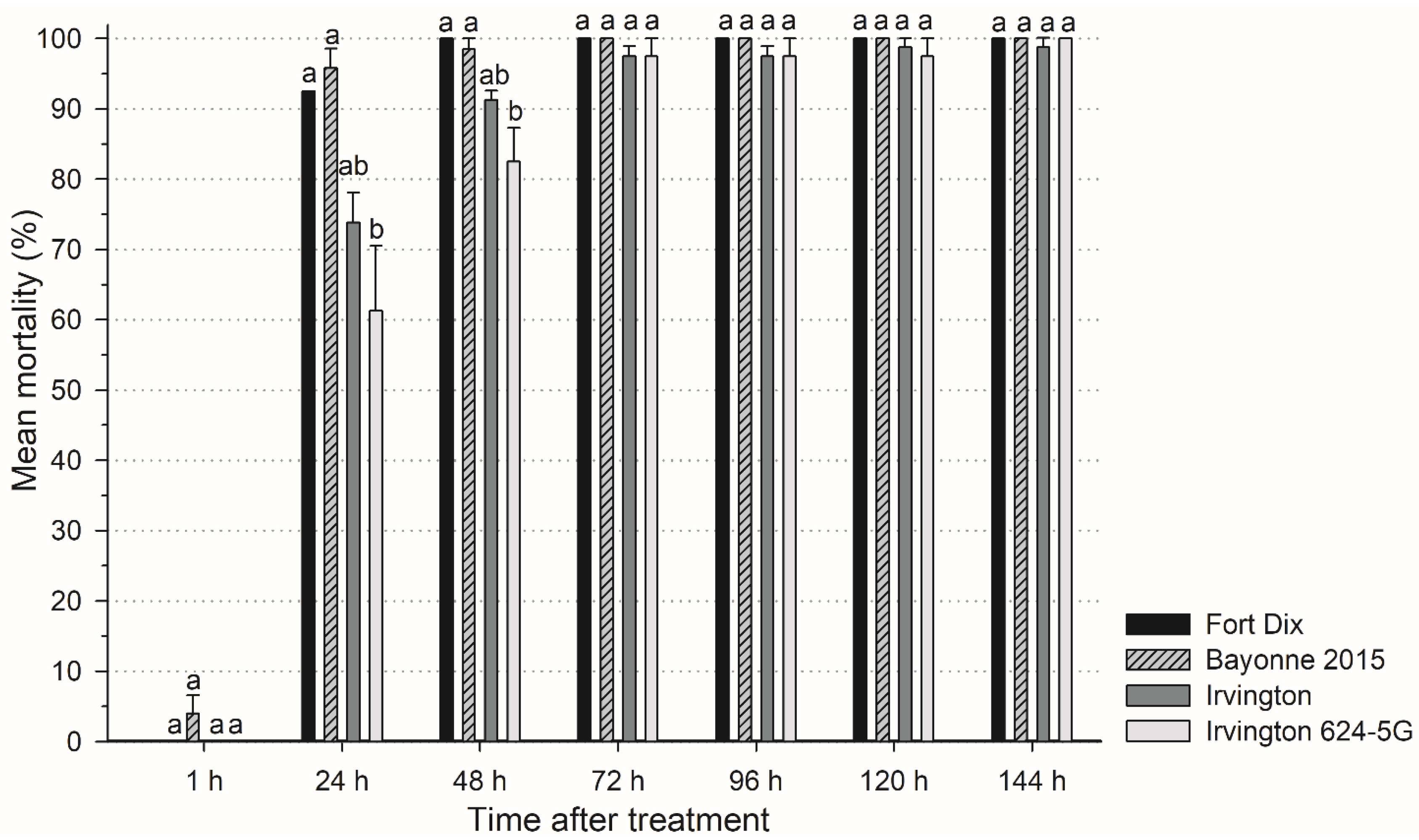
3.2.2. Inorganic Dust
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Usinger, R. Monograph of Cimicidae (Hemiptera, Heteroptera); Entomological Society of America: Annapolis, MD, USA, 1966. [Google Scholar]
- Doggett, S.L.; Geary, M.J.; Russell, R.C. The resurgence of bed bugs in Australia: With notes on their ecology and control. Environ. Health 2004, 4, 30–38. [Google Scholar]
- Potter, M.F. The perfect storm: An extension view on bed bugs. Am. Entomol. 2006, 52, 102–104. [Google Scholar] [CrossRef]
- Boase, C. Bedbugs-back from the brink. Pestic. Outlook 2001, 12, 159–162. [Google Scholar] [CrossRef]
- Doggett, S.L.; Miller, D.M.; Lee, C.-Y. (Eds.) Advances in the Biology and Management of Modern Bed Bugs; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Hwang, S.W.; Svoboda, T.J.; De Jong, I.J.; Kabasele, K.J.; Gogosis, E. Bed bug infestations in an urban environment. Emerg. Infect. Dis. 2005, 11, 533–538. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Y.; Zeng, L. Resurgence of bed bugs (Hemiptera: Cimicidae) in mainland China. Fla. Entomol. 2013, 96, 131–136. [Google Scholar] [CrossRef]
- Romero, A.; Potter, M.F.; Potter, D.A.; Haynes, K.F. Insecticide resistance in the bed bug: A factor in the pest’s sudden resurgence? J. Med. Entomol. 2007, 44, 175–178. [Google Scholar]
- Yoon, K.S.; Kwon, D.H.; Strycharz, J.P.; Hollingsworth, C.S.; Lee, S.H.; Clark, J.M. Biochemical and molecular analysis of deltamethrin resistance in the common bed bug (Hemiptera: Cimicidae). J. Med. Entomol. 2008, 45, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Doggett, S.L.; Lee, C.-Y. Historical and Contemporary Control Options Against Bed Bugs, Cimex spp. Annu. Rev. Entomol. 2022, 68, 169–190. [Google Scholar] [CrossRef]
- Kilpinen, O.; Kristensen, M.; Jensen, K.M.V. Resistance differences between chlorpyrifos and synthetic pyrethroids in Cimex lectularius population from Denmark. Parasitol. Res. 2011, 109, 1461–1464. [Google Scholar] [CrossRef]
- Tawatsin, A.; Thavara, U.; Chompoosri, J.; Phusup, Y.; Jonjang, N.; Khumsawads, C.; Bhakdeenuan, P.; Sawanpanyalert, P.; Asavadachanukorn, P.; Mulla, M.S.; et al. Insecticide resistance in bedbugs in Thailand and laboratory evaluation of insecticides for the control of Cimex hemipterus and Cimex lectularius (Hemiptera: Cimicidae). J. Med. Entomol. 2011, 48, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Zalucki, M.P.; Orton, C.J.; Russell, R.C.; Webb, C.E.; Doggett, S.L. Confirmation of insecticide resistance in Cimex lectularius Linnaeus (Hemiptera: Cimicidae) in Australia. Aust. J. Entomol. 2015, 54, 96–99. [Google Scholar] [CrossRef]
- Wang, C.; Singh, N.; Zha, C.; Cooper, R. Bed bugs: Prevalence in low-income communities, resident’s reactions, and implementation of a low-cost inspection protocol. J. Med. Entomol. 2016, 53, 639–646. [Google Scholar] [CrossRef]
- Potter, M.F.; Haynes, K.F.; Fredericks, J. Bed bugs across America: The 2015 bugs without borders survey. PestWorld 2015, 7, 4–14. [Google Scholar]
- Scharf, M.; Suiter, D. Insecticide mode of action. Pest Control Technol. 2011, 39, 10, 78–114. [Google Scholar]
- Potter, M.F. The history of bed bug management. Am. Entomol. 2011, 57, 14–25. [Google Scholar] [CrossRef]
- Gordon, J.R.; Goodman, M.H.; Potter, M.F.; Haynes, K.F. Population variation in and selection for resistance to pyrethroid-neonicotinoid insecticides in the bed bug. Sci. Rep. 2014, 4, 3836. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Kwon, D.H.; Lee, S.H.; Kim, J.H. Monitoring of pyrethroid resistance allele frequency in the common bed bug (Cimex lectularius) in the Republic of Korea. Korean J. Parasitol. 2020, 58, 99–102. [Google Scholar] [CrossRef]
- Holleman, J.G.; Robison, G.A.; Bellovich, I.J.; Booth, W. Knockdown resistance-associated mutations dominate populations of the common bed bug (Hemiptera: Cimicidae) across the South Central United States. J. Med. Entomol. 2019, 56, 1678–1683. [Google Scholar] [CrossRef]
- Zhu, F.; Gujar, H.; Gordon, J.R.; Haynes, K.F.; Potter, M.F.; Palli, S.R. Bed bugs evolved unique adaptive strategy to resist pyrethroid insecticides. Sci. Rep. 2013, 3, 1–8. [Google Scholar] [CrossRef]
- Lilly, D.G.; Dang, K.; Webb, C.E.; Doggett, S.L. Evidence for Metabolic Pyrethroid Resistance in the Common Bed Bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2016, 109, 1364–1368. [Google Scholar] [CrossRef]
- Gonzalez-Morales, M.A.; Romero, A. Effect of synergists on deltamethrin resistance in the common bed bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2019, 112, 786–791. [Google Scholar] [CrossRef]
- Cáceres, M.; Santo-Orihuela, P.L.; Vassena, C.V. Evaluation of resistance to different insecticides and metabolic detoxification mechanism by use of synergist in the common bed bug (Heteroptera: Cimicidae). J. Med. Entomol. 2019, 56, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Toi, C.S.; Lilly, D.G.; Bu, W.; Doggett, S.L. Detection of knockdown resistance mutations in the common bed bug, Cimex lectularius (Hemiptera: Cimicidae), in Australia. Pest Manag. Sci. 2015, 71, 914–922. [Google Scholar] [CrossRef]
- Zhu, F.; Wigginton, J.; Romero, A.; Moore, A.; Ferguson, K.; Palli, R.; Potter, M.F.; Haynes, K.F.; Palli, S.R. Widespread distribution of knockdown resistance mutations in the bed bug, Cimex lectularius (Hemiptera: Cimicidae), populations in the United States. Arch. Insect Biochem. Physiol. 2010, 73, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Latham, S.L.; Webb, C.E.; Doggett, S.L. Cuticle thickening in a pyrethroid-resistant strain of the common bed bug, Cimex lectularius L. (Hemiptera: Cimicidae). PLoS ONE 2016, 11, e0153302. [Google Scholar] [CrossRef] [PubMed]
- Potter, M.; Haynes, K.; Gordon, J.; Hardebeck, E.; Wickemeyer, W. Dual-action bed bug killers. Pest Control Technol. 2012, 40, 62–68, 75–76. [Google Scholar]
- Wang, C.; Singh, N.; Cooper, R. Field study of the comparative efficacy of three pyrethroid/neonicotinoid mixture products for the control of the common bed bug, Cimex lectularius. Insects 2015, 6, 197–205. [Google Scholar] [CrossRef]
- Gordon, J.R.; Potter, M.F.; Haynes, K.F. Insecticide resistance in the bed bug comes with a cost. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef]
- Bloomquist, J.R. Insecticides: Chemistries and Characteristics. University of Minnesota’s Electronic Textbook of Integrated Pest Management. 2009. Available online: https://ipmworld.umn.edu/bloomquist-insecticides (accessed on 2 December 2022).
- Wang, C.; Singh, N.; Zha, C.; Cooper, R. Efficacy of selected insecticide sprays and aerosols against the common bed bug. Cimex lectularius (Hemiptera: Cimicidae). Insects 2016, 7, 5. [Google Scholar]
- Romero, A.; Anderson, T.D. High levels of resistance in the common bed bug, Cimex lectularius (Hemiptera: Cimicidae), to neonicotinoid insecticides. J. Med. Entomol. 2016, 53, 727–731. [Google Scholar] [CrossRef]
- Campbell, B.E.; Miller, D.M. Insecticide resistance in eggs and first instars of the bed bug, Cimex lectularius (Hemiptera: Cimicidae). Insects 2015, 6, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Singham, G.V.; Doggett, S.L.; Lilly, D.G.; Lee, C.-Y. Effects of different surfaces and insecticide carriers on residual insecticide bioassays against bed bugs, Cimex spp. (Hemiptera: Cimicidae). J. Econ. Entomol. 2017, 110, 558–566. [Google Scholar] [PubMed]
- Akhtar, Y.; Isman, M.B. Horizontal transfer of diatomaceous earth and botanical insecticides in the common bed bug, Cimex lectularius L.; Hemiptera: Cimicidae. PLoS ONE 2013, 8, e75626. [Google Scholar] [CrossRef]
- Agnew, J.L.; Romero, A. Behavioral responses of the common bed bug, Cimex lectularius, to insecticide dusts. Insects 2017, 8, 83. [Google Scholar] [CrossRef]
- Romero, A. Moving from the old to the new: Insecticide research on bed bugs since the resurgence. Insects 2011, 2, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.; Field, L.; Williamson, M. The re-emergence of the bed bug as a nuisance pest: Implications of resistance to the pyrethroid insecticides. Med. Vet. Entomol. 2012, 26, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Kuronic, Z. Diatomaceous earths, a group of natural insecticides. Review. J. Stored Prod. Res. 1998, 34, 87–97. [Google Scholar] [CrossRef]
- Quarles, W.; Winn, P.S. Diatomaceous earth and stored product pests. IPM Pract. 1996, 18, 1–10. [Google Scholar]
- Benoit, J.B.; Phillips, S.A.; Croxall, T.J.; Christensen, B.S.; Yoder, J.A.; Denlinger, D.L. Addition of alarm pheromone components improves the effectiveness of desiccant dusts against Cimex lectularius. J. Med. Entomol. 2014, 46, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Wang, C.; Wang, D.; Cooper, R.; Zha, C. Comparative efficacy of selected dust insecticides for controlling Cimex lectularius (Hemiptera: Cimicidae). J. Econ. Entomol. 2016, 109, 1819–1826. [Google Scholar] [CrossRef]
- Ranabhat, S.; Wang, C. Effect of moisture on efficacy of selected insecticide dusts against the common bed bug, Cimex lectularius (Hemiptera: Cimicidae). J. Econ. Entomol. 2020, 113, 1933–1939. [Google Scholar] [CrossRef] [PubMed]
- Bartley, J.D.; Harlan, H.J. Bed bug infestation: Its control and management. Mil. Med. 1974, 139, 884–886. [Google Scholar] [CrossRef]
- Dang, K.; Doggett, S.L.; Leong, X.-Y.; Veera Singham, G.; Lee, C.-Y. Multiple Mechanisms Conferring Broad-Spectrum Insecticide Resistance in the Tropical Bed Bug (Hemiptera: Cimicidae). J. Econ. Entomol. 2021, 114, 2473–2484. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Lee, L.-C.; Lee, C.-Y. Insecticide resistance profiles and possible underlying mechanisms in German cockroaches, Blattella germanica (Linnaeus) (Dictyoptera: Blattellidae) from Peninsular Malaysia. Med. Entomol. Zool. 2004, 55, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Potter, M.; Haynes, K. Insecticide resistant bed bugs: Implications for the industry. Pest Control Technol. 2007, 35, 7, 42–46. [Google Scholar]
- Moore, D.J.; Miller, D.M. Laboratory evaluations of insecticide product efficacy for control of Cimex lectularius. J. Econ. Entomol. 2006, 99, 2080–2086. [Google Scholar] [CrossRef]
- Adelman, Z.N.; Kilcullen, K.A.; Koganemaru, R.; Anderson, M.A.; Anderson, T.D.; Miller, D.M. Deep sequencing of pyrethroid-resistant bed bugs reveals multiple mechanisms of resistance within a single population. PLoS ONE 2011, 6, e26228. [Google Scholar] [CrossRef]
- Anderson, J.F.; Cowles, R.S. Susceptibility of Cimex lectularius (Hemiptera: Cimicidae) to pyrethroid insecticides and to insecticidal dusts with or without pyrethroid insecticides. J. Econ. Entomol. 2012, 105, 1789–1795. [Google Scholar] [CrossRef]
- Seong, K.M.; Lee, D.-Y.; Yoon, K.S.; Kwon, D.H.; Kim, H.C.; Klein, T.A.; Clark, J.M.; Lee, S.H. Establishment of quantitative sequencing and filter contact vial bioassay for monitoring pyrethroid resistance in the common bed bug, Cimex lectularius. J. Med. Entomol. 2010, 47, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Polanco, A.M.; Brewster, C.C.; Miller, D.M. Population growth potential of the bed bug, Cimex lectularius L.: A life table analysis. Insects 2011, 2, 173–185. [Google Scholar] [CrossRef]
- Lewis, C.D.; Levine, B.A.; Schal, C.; Vargo, E.L.; Booth, W. Decade long upsurge in mutations associated with pyrethroid resistance in bed bug populations in the USA. J. Pest. Sci. 2023, 96, 415–423. [Google Scholar] [CrossRef]
- Leong, X.-Y.; Kim, D.-Y.; Dang, K.; Singham, G.V.; Doggett, S.L.; Lee, C.-Y. Performance of commercial insecticide formulations against different developmental stages of insecticide-resistant tropical bed bugs (Hemiptera: Cimicidae). J. Econ. Entomol. 2020, 113, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Mamidala, P.; Jones, S.C.; Mittapalli, O. Metabolic resistance in bed bugs. Insects 2011, 2, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Doggett, S.L.; Singham, G.V.; Lee, C.-Y. Insecticide resistance and resistance mechanisms in bed bugs, Cimex spp. (Hemiptera: Cimicidae). Parasites Vectors 2017, 10, 318. [Google Scholar] [CrossRef]
- Lilly, D.; Doggett, S.; Zalucki, M.; Orton, C.; Russell, R. Bed bugs that bite back, confirmation of insecticide resistance in the common bed bug, Cimex lectularius. Prof. Pest Manag. 2009, 13, 22–24. [Google Scholar]
- Dang, K.; Doggett, S.L.; Lee, C.-Y. Performance of pyrethroid-neonicotinoid mixture formulations against field-collected strains of the tropical bed bug (Hemiptera: Cimicidae) on different substrates. J. Econ. Entomol. 2022. [Google Scholar] [CrossRef]
- Zhu, F.; Sams, S.; Moural, T.; Haynes, K.F.; Potter, M.F.; Palli, S.R. RNA interference of NADPH-cytochrome P450 reductase results in reduced insecticide resistance in the bed bug, Cimex lectularius. PLoS ONE 2012, 7, e31037. [Google Scholar] [CrossRef]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Nauen, R.; Denholm, I. Resistance of insect pests to neonicotinoid insecticides: Current status and future prospects. Arch. Ins. Biochem. Physiol. 2005, 58, 200–215. [Google Scholar] [CrossRef]
- Tarshis, I.B. Laboratory and field studies with sorptive dusts for the control of arthropods affecting man and animal. Exp. Parasitol. 1961, 11, 10–33. [Google Scholar] [CrossRef] [PubMed]
- Ebeling, W. Sorptive dusts for pest control. Annu. Rev. Entomol. 1971, 16, 123–158. [Google Scholar] [CrossRef]
- Potter, M.; Haynes, K.; Gordon, J.; Washburn, L.; Washburn, M.; Hardin, T. Silica gel: A better bed bug desiccant. Pest Control Technol. 2014, 42, 76–81. [Google Scholar]
- Abbar, S.; Wang, C.; Cooper, R. Evaluation of a non-chemical compared to a non-chemical plus silica gel approach to bed bug management. Insects 2020, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Lilly, D.G.; Webb, C.E.; Doggett, S.L. Evidence of tolerance to silica-based desiccant dusts in a pyrethroid-resistant strain of Cimex lectularius (Hemiptera: Cimicidae). Insects 2016, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Soh, L.S.; Veera Singham, G. Cuticle thickening associated with fenitrothion and imidacloprid resistance and influence of voltage-gated sodium channel mutations on pyrethroid resistance in the tropical bed bug, Cimex hemipterus. Pest Manag. Sci. 2021, 77, 5202–5212. [Google Scholar] [CrossRef] [PubMed]
- Yahouédo, G.A.; Chandre, F.; Rossignol, M.; Ginibre, C.; Balabanidou, V.; Mendez, N.G.A.; Pigeon, O.; Vontas, J.; Cornelie, S. Contributions of cuticle permeability and enzyme detoxification to pyrethroid resistance in the major malaria vector Anopheles gambiae. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Yu, J.J.; Bong, L.J.; Panthawong, A.; Chareonviriyaphap, T.; Liu, W.T.; Neoh, K.B. Effects of piperonyl butoxide synergism and cuticular thickening on the contact irritancy response of field Aedes aegypti (Diptera: Culicidae) to deltamethrin. Pest Manag. Sci. 2021, 77, 5557–5565. [Google Scholar] [CrossRef]


{kind=link}
| Insecticide | * N | Model Parameters | Lethal Dose (ng/μL) | Model Fit | ||||
|---|---|---|---|---|---|---|---|---|
| ** Intercept ± SE | Slope ± SE | LD50 (95% CI) | LD90 (95% CI) | χ2 | df | p | ||
| Acetamiprid | 150 | −0.636 ± 0.162 | 1.509 ± 0.223 | 2.64 (1.78–3.80) | 18.64 (11.10–43.56) | 4.207 | 3 | 0.240 |
| Imidacloprid | 150 | −1.096 ± 0.220 | 2.873 ± 0.390 | 2.41 (1.92–2.94) | 6.73 (5.21–9.87) | 2.151 | 3 | 0.542 |
| Deltamethrin | 150 | 0.053 ± 0.188 | 1.156 ± 0.264 | 0.90 (0.55–2.78) | 11.49 (3.42–250.84) | 0.327 | 3 | 0.955 |
| Population | n | KT50 (min, 95% CI) | KT90 (min, 95% CI) | χ2 (df) | Slope ± SE | % Mean Mortality (±SE) at 72 h | * RR50 | RR90 |
|---|---|---|---|---|---|---|---|---|
| Fort Dix | 30 | 15 (13–18) | 46 (39–56) | 9.2 (11) | 2.692 ± 0.266 | 100 ± 0 | - | - |
| Aberdeen | 30 | 61 (51–76) | >4320 | 6.9 (12) | 1.694 ± 0.210 | 57 ± 3 | 4.1 | >93.9 |
| Bayonne | 30 | 37 (31–42) | 141 (110–200) | 4.8 (11) | 2.191 ± 0.220 | 100 ± 0 | 2.5 | 3.1 |
| Bayonne 2015 | 30 | 16 (12–19) | 80 (57–140) | 4.4 (7) | 1.814 ± 0.257 | 100 ± 0 | 1.0 | 1.7 |
| Canfield | 30 | 21 (17–24) | 75 (61–97) | 7.4 (12) | 2.290 ± 0.222 | 100 ± 0 | 1.4 | 1.6 |
| Cotton | 30 | 32 (28–39) | 126 (94–200) | 4.4 (10) | 2.159 ± 0.258 | 100 ± 0 | 2.1 | 2.7 |
| Dehart | 30 | 11 (7–14) | 65 (46–118) | 3.6 (7) | 1.636 ± 0.262 | 100 ± 0 | 1.0 | 1.4 |
| Hackensack | 30 | 28 (23–33) | 71 (54–119) | 15.5 (9) | 3.125 ± 0.360 | 100 ± 0 | 1.9 | 1.5 |
| Indy | 30 | 39 (35–44) | 115 (92–159) | 10.9 (12) | 2.716 ± 0.288 | 100 ± 0 | 2.6 | 2.5 |
| Irvington | 30 | 16 (8–24) | 82 (47–375) | 48.4 (11) | 1.804 ± 0.207 | 100 ± 0 | 1.1 | 1.8 |
| Irvington 624-5G | 30 | 26 (22–30) | 126 (96–188) | 14.1 (13) | 1.868 ± 0.204 | 100 ± 0 | 1.7 | 2.7 |
| Linden 2019 | 30 | >4320 | >4320 | - | - | 13 ± 3 | >288 | >93.9 |
| Masiello | 30 | 43 (36–50) | 239 (174–382) | 6.2 (16) | 1.711 ± 0.181 | 100 ± 0 | 2.9 | 5.2 |
| New Brunswick | 30 | 54 (46–65) | 249 (175–430) | 5.1 (10) | 1.934 ± 0.232 | 93 ± 3 | 3.6 | 5.4 |
| Population | n | KT50 (min, 95% CI) | KT90 (min, 95% CI) | χ2 (df) | Slope ± SE | % Mean Mortality (±SE) at 72 h | * RR50 | RR90 |
|---|---|---|---|---|---|---|---|---|
| Fort Dix | 30 | 16 (11–20) | 53 (42–77) | 12.1 (7) | 2.445 ± 0.317 | 100 ± 0 | - | - |
| Aberdeen | 30 | 50 (41–62) | 136 (98–255) | 32.0 (13) | 2.928 ± 0.291 | 100 ± 0 | 3.1 | 2.6 |
| Bayonne | 30 | 38 (31–48) | 122 (87–218) | 14.8 (9) | 2.532 ± 0.264 | 100 ± 0 | 2.4 | 2.3 |
| Bayonne 2015 | 30 | 20 (16–24) | 80 (60–127) | 5.7 (6) | 2.108 ± 0.272 | 100 ± 0 | 1.3 | 1.5 |
| Canfield | 30 | 19 (16–22) | 69 (52–112) | 10.3 (7) | 2.255 ± 0.309 | 100 ± 0 | 1.2 | 1.3 |
| Cotton | 30 | 26 (21–33) | 93 (65–179) | 15.5 (9) | 2.344 ± 0.263 | 100 ± 0 | 1.6 | 1.8 |
| Dehart | 30 | 34 (30–40) | 95 (73–144) | 7.8 (7) | 2.876 ± 0.365 | 100 ± 0 | 2.1 | 1.8 |
| Hackensack | 30 | 23 (21–26) | 53 (43–71) | 6.9 (6) | 3.627 ± 0.458 | 100 ± 0 | 1.4 | 1.0 |
| Indy | 30 | 38 (31–50) | 126 (85–259) | 1.5 (5) | 2.465 ± 0.381 | 100 ± 0 | 2.4 | 2.4 |
| Irvington | 30 | 19 (16–22) | 68 (54–94) | 11.2 (9) | 2.305 ± 0.263 | 100 ± 0 | 1.2 | 1.3 |
| Irvington 624-5G | 30 | 39 (33–46) | 174 (123–302) | 15.7 (11) | 1.960 ± 0.240 | 100 ± 0 | 2.4 | 3.3 |
| Linden 2019 | 30 | 1231 (760–2361) | >4320 | 10.2 (15) | 0.856 ± 0.087 | 60 ± 6 | 76.9 | >81.5 |
| Masiello | 20 | 35 (30–40) | 102 (80–148) | 6.1 (11) | 2.728 ± 0.332 | 100 ± 0 | 2.2 | 1.9 |
| New Brunswick | 20 | 75 (58–108) | 394 (223–1194) | 4.1 (8) | 1.775 ± 0.307 | 100 ± 0 | 4.7 | 7.4 |
| Population | n | KT50 (min, 95% CI) | KT90 (min, 95% CI) | χ2 (df) | Slope ± SE | % Mean Mortality at 72 h | * RR50 | RR90 |
|---|---|---|---|---|---|---|---|---|
| Fort Dix | 30 | 27 (24–29) | 47 (43–53) | 4.8 (10) | 5.206 ± 0.467 | 100 | - | - |
| Aberdeen | 30 | >4320 | >4320 | - | - | 17 ± 7 | >160 | >91.9 |
| Bayonne | 30 | 44 (39–48) | 86 (75–105) | 2.0 (9) | 4.294 ± 0.438 | 97 ± 3 | 1.6 | 1.8 |
| Bayonne 2015 | 30 | >4320 | >4320 | - | - | 13 ± 9 | >160 | >91.9 |
| Canfield | 30 | >4320 | >4320 | - | - | 20 ± 6 | >160 | >91.9 |
| Cotton | 30 | 44 (377–52) | >4320 | 4.7 (17) | 1.508 ± 0.166 | 73 ± 9 | 1.6 | >91.9 |
| Dehart | 30 | 24 (21–27) | 55 (47–69) | 2.7 (7) | 3.593 ± 0.394 | 93 ± 3 | 1.0 | 1.2 |
| Hackensack | 30 | 27 (21–34) | 56 (41–120) | 19.3 (7) | 4.015 ± 0.493 | 100 | 1.0 | 1.2 |
| Indy | 30 | 48 (40–64) | >4320 | 3.6 (10) | 1.699 ± 0.253 | 67 ± 7 | 1.8 | >91.9 |
| Irvington | 30 | >4320 | >4320 | - | - | 27 ± 7 | >160 | >91.9 |
| Irvington 624-5G | 30 | >4320 | >4320 | - | - | 0 | >160 | >91.9 |
| Linden 2019 | 30 | >4320 | >4320 | - | - | 3 ± 3 | >160 | >91.9 |
| Masiello | 30 | 54 (46–59) | 162 (137–202) | 9.9 (19) | 2.687 ± 0.217 | 90 ± 10 | 2.0 | 3.4 |
| New Brunswick | 30 | >4320 | >4320 | - | - | 7 ± 7 | >160 | >91.9 |
| Spray | Population | Slope (±SE) | Lethal Concentration (%) | Performance Ratio | Model Fit | |||
|---|---|---|---|---|---|---|---|---|
| * LC50 (95% CI) | LC90 (95% CI) | ** PR50 | PR90 | χ2 | df | |||
| Transport GHP | Fort Dix | 2.9 ± 0.4 | 0.0002 (0.0002–0.0003) a | 0.0006 (0.0005–0.0009) a | - | - | 2.6 | 4 |
| Bayonne 2015 | 2.1 ± 0.3 | 0.51 (0.31–0.67) b | 1.13 (0.93–1.56) b | 2550 | 1883 | 9.3 | 4 | |
| Irvington | 1.1 ± 0.3 | 0.04 (0.01–0.08) c | 0.54 (0.25–4.29) bc | 200 | 900 | 9.6 | 4 | |
| Irvington 624-5G | 1.7 ± 0.2 | 0.45 (0.35–0.54) b | 1.21 (1.08–1.40) b | 2250 | 2017 | 19.1 | 4 | |
| Temprid SC | Fort Dix | 1.9 ± 0.4 | 0.0012 (0.0005–0.0023) a | 0.0055 (0.0028–0.0365) a | - | - | 8.2 | 4 |
| Bayonne 2015 | 2.0 ± 0.3 | 0.16 (0.12–0.22) b | 0.71 (0.47–1.39) b | 133 | 129 | 13.7 | 4 | |
| Irvington | 1.1 ± 0.2 | 0.03 (0.01–0.07) c | 0.41 (0.13–15.30) bc | 25 | 75 | 9.3 | 4 | |
| Irvington 624-5G | 2.6 ± 0.3 | 0.10 (0.08–0.12) d | 0.30 (0.23–0.41) c | 83 | 55 | 4.2 | 4 | |
| Tandem | Fort Dix | 1.2 ± 0.1 | 0.0004 (0.0003–0.0006) a | 0.0052 (0.0032–0.0096) a | - | - | 5.3 | 4 |
| Bayonne 2015 | 2.5 ± 0.4 | 0.58 (0.45–0.75) b | 1.01 (0.88–1.57) b | 1450 | 194 | 8.5 | 4 | |
| Irvington | 1.6 ± 0.2 | 0.08 (0.52–0.11) c | 0.52 (0.33–1.01) b | 200 | 100 | 4.9 | 4 | |
| Irvington 624-5G | 2.0 ± 0.0 | 0.24 (0.19–0.30) d | 1.02 (0.75–1.59) b | 600 | 196 | 3.2 | 4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.-J.; Ranabhat, S.; Wang, C. Insecticide Resistance of Cimex lectularius L. Populations and the Performance of Selected Neonicotinoid-Pyrethroid Mixture Sprays and an Inorganic Dust. Insects 2023, 14, 133. https://doi.org/10.3390/insects14020133
Yu J-J, Ranabhat S, Wang C. Insecticide Resistance of Cimex lectularius L. Populations and the Performance of Selected Neonicotinoid-Pyrethroid Mixture Sprays and an Inorganic Dust. Insects. 2023; 14(2):133. https://doi.org/10.3390/insects14020133
Chicago/Turabian StyleYu, Jin-Jia, Sabita Ranabhat, and Changlu Wang. 2023. "Insecticide Resistance of Cimex lectularius L. Populations and the Performance of Selected Neonicotinoid-Pyrethroid Mixture Sprays and an Inorganic Dust" Insects 14, no. 2: 133. https://doi.org/10.3390/insects14020133
APA StyleYu, J.-J., Ranabhat, S., & Wang, C. (2023). Insecticide Resistance of Cimex lectularius L. Populations and the Performance of Selected Neonicotinoid-Pyrethroid Mixture Sprays and an Inorganic Dust. Insects, 14(2), 133. https://doi.org/10.3390/insects14020133